Cell Death of P. vivax Blood Stages Occurs in Absence of Classical Apoptotic Events and Induces Eryptosis of Parasitized Host Cells
Carolina Moreira Blanco, Hugo Amorim dos Santos de Souza, Priscilla da Costa Martins, Juliana Almeida-Silva, Ana Marcia Suarez-Fontes, Yury Oliveira Chaves, Marcos André Vannier-Santos, Lilian Rose Pratt-Riccio, Cláudio Tadeu Daniel-Ribeiro, Stefanie Costa Pinto Lopes

TL;DR
This study shows that P. vivax parasites die without typical apoptosis signs and cause host cell death during infection.
Contribution
The first study to investigate apoptosis-like cell death in P. vivax and its effect on host eryptosis.
Findings
P. vivax cell death occurs without DNA fragmentation or chromatin condensation.
Staurosporine reduces P. vivax viability but does not trigger classical apoptotic markers.
Dying P. vivax parasites induce eryptosis in host erythrocytes marked by phosphatidylserine externalization.
Abstract
Elucidation of pathways regulating parasite cell death is believed to contribute to identification of novel therapeutic targets for protozoan diseases, and in this context, apoptosis-like cell death has been reported in different groups of protozoa, in which metacaspases seem to play a role. In the genus Plasmodium, apoptotic markers have been detected in P. falciparum and P. berghei, and no study focusing on P. vivax cell death has been reported so far. In the present study, we investigated the susceptibility of P. vivax to undergo apoptotic cell death after incubating mature trophozoites with the classical apoptosis inducer staurosporine. As assessed by flow cytometry assays, staurosporine inhibited parasite intraerythrocytic development, which was accompanied by a decrease in cell viability, evidenced by reduced plasmodial mitochondrial activity. However, typical signs of apoptosis,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
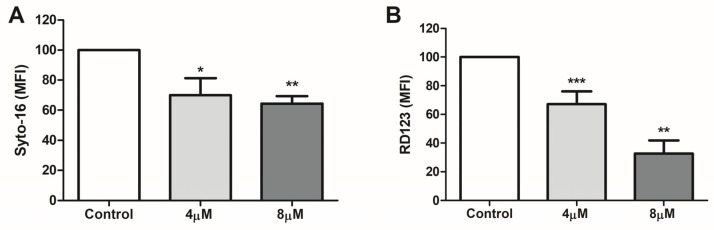 Figure 1
Figure 1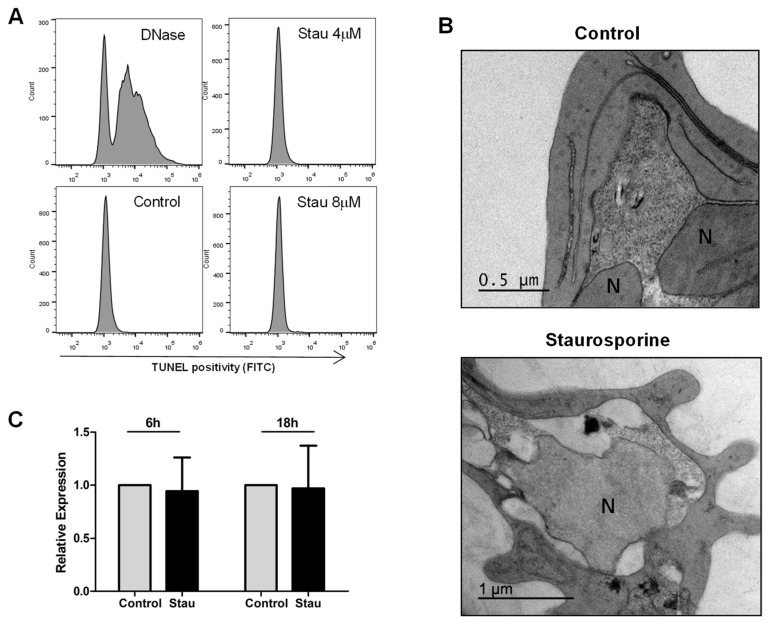 Figure 2
Figure 2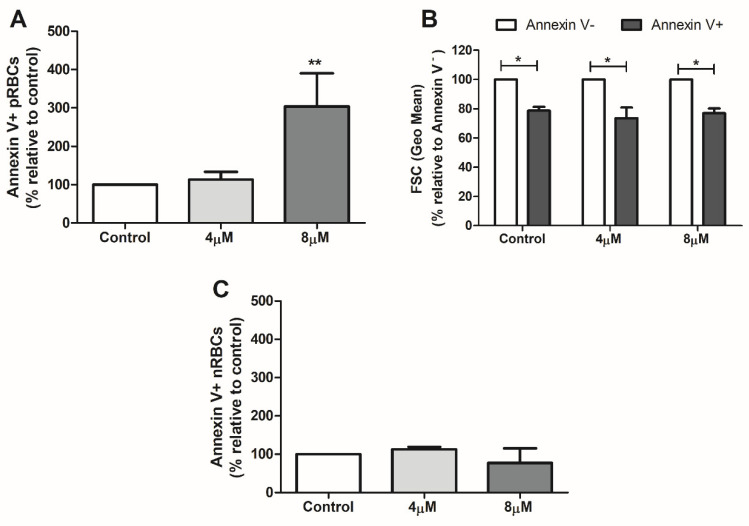 Figure 3
Figure 3- —Instituto Oswaldo Cruz (IOC-Fiocruz)
- —Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ)
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCell death mechanisms and regulation · Toxin Mechanisms and Immunotoxins
1. Introduction
Malaria is a parasitic disease caused by protozoa of the genus Plasmodium, and is transmitted to humans by female Anopheles mosquitoes. This tropical disease remains one of the major public health problems in the world, accounting for 249 million cases and 608 thousand deaths worldwide in 2022 according to WHO estimates [1]. In this scenario, P. vivax is the most geographically widespread Plasmodium species and contributes significantly to the malaria burden outside Africa [1]. Moreover, P. vivax parasites resistant to chloroquine, which comprises the first-line treatment for vivax malaria in combination with primaquine, have been reported in many endemic areas, including Brazil, constituting a concern for malaria control programs [2,3].
In view of the risk of parasite chemoresistance and the consequent need for development of new antimalarial drugs, knowledge about the machinery of cell death in Plasmodium is of ultimate importance. Although the signaling pathways driving cell death in unicellular eukaryotes are unknown, hallmarks of apoptosis have been observed in different protozoa, such as Toxoplasma gondii, Blastocystis hominis, and Trichomonas vaginalis and species of the family Trypanosomatidae, under different stimuli, suggesting susceptibility to regulated cell death (RCD) processes [4,5,6,7]. Indeed, the use of caspase inhibitors argues for the existence of a cysteine protease-RCD in protozoa [8,9]. However, genes encoding caspases are absent in non-metazoan organisms, and the involvement of a cysteine protease family structurally related to caspases, named metacaspase, has been considered [9,10].
Metacaspases are expressed in the genus Plasmodium [11,12], and the capacity of P. falciparum metacaspase 1 (PfMCA1) to trigger apoptosis has been demonstrated in a yeast model [13]. In parallel, apoptotic events, including phosphatidylserine externalization, chromatin condensation, DNA fragmentation, and caspase-like activity, have been reported in P. falciparum blood stages and P. berghei ookinetes [14,15,16,17,18,19]. Nevertheless, in our earlier studies, blood stages of P. falciparum underwent a caspase-like independent cell death process marked by morphological evidence of autophagy [20], while a necrotic phenotype was detected by Porter and colleagues [21]. In P. vivax, although the genetic diversity of metacaspase 1 (PvMCA1) has been studied in different endemic countries [22,23,24], no focus has been given to the profile of cell death suffered by this Plasmodium species.
Therefore, in the present study, we investigated for the first time the susceptibility of P. vivax blood forms to undergo apoptotic events using a classical inducer of apoptosis, staurosporine.
2. Materials and Methods
2.1. P. vivax Parasites and Cell Death Induction
Clinical isolates of P. vivax were obtained from patients attending the Fundação de Medicina Tropical Doutor Heitor Vieira Dourado (FMT-HVD) in Manaus, Amazonas State, Brazil, according to the procedures approved by the Research Ethics Committee of FMT-HVD (CAAE 54234216.1.0000.0005 and 75894223.9.0000.0005). Patients presenting parasitemia higher than 500 parasites/µL blood with a predominance of asexual forms at the trophozoite stage (>60%), as evaluated by thick blood smears, were included and venous blood samples of each patient were collected by venipuncture with heparinized tubes.
Enrichment of parasites was performed as previously described [25]. Briefly, blood samples were centrifuged for plasma separation, leukocytes were depleted by filtration using columns of cellulose (Sigma-Aldrich, St. Louis, MO, USA), and blood pellets were diluted in RPMI 1640 medium (Sigma-Aldrich) to 10% hematocrit. Trophozoites were enriched in a 45% Percoll density gradient (GE-Healthcare, Uppsala, Sweden), washed with RPMI 1640 medium (Sigma), and then resuspended in IMDM (Gibco Industries, Big Cabin, OK, USA) supplemented with 20% of heat-inactivated human AB serum.
To evaluate the effect of staurosporine (Sigma) on the growth and cell death of P. vivax, enriched parasitized red blood cells (pRBCs; 1 × 10^6^ pRBCs/well) were incubated for 6–24 h in 96-well culture plates at 37 °C under an atmosphere of 5% CO_2_ (White Martins, Manaus, Brazil) in the presence or absence of staurosporine at concentrations (4 µM and 8 µM) described to induce cell death in different group of protozoa, including P. falciparum [4,5,7,9,20,21,26,27,28,29].
2.2. Evaluation of Parasite Growth Inhibition
Inhibition of parasite maturation was assessed by flow cytometry after 24 h using the Syto-16 nucleic acid stain (Thermo Fisher Scientific, Eugene, OR, USA) [30]. Briefly, pRBCs were washed by centrifugation (350× g, 5 min) in phosphate-buffered saline (PBS—0.15 mM NaCl, 2.5 mM KCl, 8 mM Na_2_HPO_4_ 7H_2_O, 1.5 mM KH_2_PO_4_), resuspended in PBS containing 100 µM Syto-16, and incubated at 37 °C for 40 min. After incubation, pRBCs were washed and resuspended in PBS, and then a minimum of 50,000 events were acquired on a FACSCanto II flow cytometer equipped with blue (488 nm) and red (633 nm) lasers (Becton Dickinson, Franklin Lakes, NJ, USA) using the FL1 channel (530/30 nm). Data were analyzed using FlowJo 10.0 software (Becton Dickinson) and are expressed as mean fluorescence intensity (MFI).
2.3. Viability and Apoptosis Assays
Parasite viability was assessed by measuring the mitochondrial transmembrane potential (ΔΨm) using rhodamine 123 (Thermo Fisher Scientific) as previously described [20], with modifications. After 24 h incubation, pRBCs were washed by centrifugation in PBS (350× g, 5 min) and incubated at 37 °C for 5 min in 1 µg/mL rhodamine. After removing rhodamine solution, pRBCs were washed and incubated for 30 min in PBS, resuspended in the same buffer, and analyzed in a flow cytometer. Cells were acquired using the FL1 channel, and data are expressed as mean fluorescence intensity (MFI).
The TUNEL (terminal deoxynucleotidyl transferase dUTP nick-end labeling) assay was used to verify parasite DNA integrity. Initially, pRBCs (5 × 10^6^) were treated with 0.1% saponin in PBS to obtain free parasites, as previously described [9], and then parasites were stained using an APO-DIRECT KIT (BD Pharmingen, San Diego, CA, USA) according to the manufacturer’s instructions. Briefly, parasites were fixed in 2% paraformaldehyde (1 h, 4 °C), washed twice by centrifugation in PBS (400× g, 5 min) and permeabilized with 70% ice-cold ethanol at −20 °C. Following removal of ethanol, parasites were incubated at 37 °C for 1 h in staining solution containing terminal deoxytransferase (TdT) and FITC-tagged deoxyuridine triphosphate nucleotides (FITC-dUTP). After incubation, parasites were rinsed twice with rinse buffer and finally analyzed in PBS by a flow cytometer using the FL1 channel. As a positive control, fixed/permeabilized free parasites were previously treated for 40 min at room temperature with 10 U/mL DNase I in DNase reaction buffer provided by the manufacturer (Thermo Fisher Scientific Life Sciences Solutions, CA, USA). Data are expressed as percentage of positive cells.
Externalization of phosphatidylserine was examined on the surface of pRBCs by annexin V assay (BD Pharmingen) in combination with Syto-16 staining. Syto-16 staining was performed as indicated above: pRBCs were resuspended in PBS containing 100 µM Syto-16, incubated at 37 °C for 40 min, and washed in PBS alone. Then, pRBCs were resuspended in 100 µL annexin-binding buffer containing 5 µL annexin V–APC and after 15 min incubation at room temperature, were fivefold diluted with annexin-binding buffer and lastly analyzed by flow cytometry. FL1 (Syto-16; 530/30 nm) and FL5 (annexin V-APC; 660/20 nm) channels were used, and data are expressed as a percentage of positive cells. Forward scatter (FSC) measurement (geometric mean) was additionally performed in both positive and negative pRBCs to estimate the difference in cell size.
A FACSCanto II flow cytometer and FlowJo 10.0 software (Becton Dickinson) were used for all experiments, and a minimum of 50,000 events was acquired. Supplementary Figures S1 and S2 show the gating strategy for the analysis of Syto-16 and rhodamine 123 staining (Figure S1) as well as annexin V/Syto-16 assays (Figure S2).
2.4. Analysis of PvMCA1 Gene Expression
Transcriptional expression of PvMCA1 under cell death stimulation (staurosporine, 4 µM) was evaluated after 6 h and 18 h incubation. Parasites (1 × 10^6^) were stored in RNA stabilizing solution (Thermo Fisher Scientific Baltics, Vilnius, Lithuania) at −20 °C and total RNA was extracted using a PureLink RNA mini-Kit (Life Technologies Corporation, Austin, TX, USA), followed by treatment with DNase (Thermo Fisher Scientific Life Sciences Solutions) and reverse-transcription reaction using a high-capacity cDNA reverse transcription kit (Thermo Fisher Scientific Baltics), all according to the instructions of the manufacturers. cDNA obtained was quantified by a Qubit ssDNA Assay Kit (Molecular Probes, Eugene, OR, USA) and quantitative polymerase chain reaction (qPCR) was performed with a PowerUp SYBR Green Master Mix kit (Thermo Fisher Scientific Baltics) in a 7500 Real-Time PCR System (Applied Biosystems, Waltham, MA, USA). The qPCR conditions used were as follows: 2 min at 50 °C, followed by 2 min at 95 °C and 40 cycles of denaturation (95 °C/1 s) and annealing (60 °C/1 min). The specificity of amplification was verified through analysis of melting curves (95 °C/15 s, 60 °C/1 min, 95 °C/15 s). The reaction mixture consisted of 20 µL final volume containing 1XPowerUp SYBR Green Master Mix, primers (800 nM), cDNA (10 ng), and UltraPureDNase/RNase-Free Distilled Water (Thermo Fisher Scientific Life Sciences Solutions). All reactions were performed in duplicate and included DNase untreated samples and non-template negative controls. The P. vivax β-tubulin (Pvβ-T) housekeeping gene was assayed as internal control [31] and the following oligonucleotide primers were used: PvMCA1 (forward, 5′-ACCCCAGTGGACCACCAA-3′; reverse 5’-CACGAGGGTAAGTAACCCCA-3′); Pvβ-T (forward, 5′-CCAAGAATATGATGTGTGCAAGTG-3′; reverse, 5′-GGCGCAGGCGGTTAGG-3′). CT values were analyzed in the 7500 software (version 2.0.6), in which the fold change relative to the internal control was calculated through the 2^−∆∆ct^ formula [32].
2.5. Transmission Electron Microscopy
The morphology of the parasites induced to cell death was examined by transmission electron microscopy. Briefly, pRBCs were washed with PBS and fixed at 4 °C in 0.1 M sodium cacodylate buffer containing 2.5% glutaraldehyde. Then, samples were post-fixed in 1% osmium tetroxide, 5 mM calcium chloride, and 0.8% potassium ferricyanide in sodium cacodylate buffer, followed by dehydrating in acetone. Samples were embedded in Polybed epoxy resin (Polysciences Inc., Warrington, PA, USA), and after polymerization at 60 °C, were sectioned on an ultramicrotome. The sections were collected on copper grids, counterstained with 3% lead citrate and 5% uranyl acetate in water, and observed under a transmission electron microscope (JEOL-JEM-1011; JEOL USA Inc., Peabody, MA, USA).
2.6. Statistical Analysis
Statistical analyses were performed using GraphPad Prism 5.0 software (San Diego, CA, USA) and differences were tested by t test or one-way ANOVA with Dunnett’s post-test. A p-value of <0.05 was considered statistically significant.
3. Results and Discussion
To study the susceptibility of P. vivax blood stages to apoptosis-like cell death, we initially evaluated the potential of staurosporine to inhibit parasite development after 24 h incubation of mature trophozoites, which are the most metabolic stages of the Plasmodium intraerythrocytic cycle [33], constituting ideal targets for our pioneering investigation on P. vivax cell death. Staurosporine is a broad-spectrum protein kinase inhibitor that has been widely used as an inducer of apoptosis in metazoan cells [34,35], presenting also the capacity to both inhibit cell growth and trigger apoptotic events in different groups of protozoa [7,36,37]. Indeed, protein kinases are key components of cell signaling and regulation found ubiquitously across the domains of life [38], and their blockage in Plasmodium has been shown to arrest the progression of intraerythrocytic development in a dose-dependent manner, with the appearance of abnormal morphological forms [39,40]. Therefore, it was not surprising that staurosporine also significantly inhibited the maturation of P. vivax trophozoites, as herein evidenced by the diminished DNA content in flow cytometry assays compared to non-treated parasites (Figure 1A).
Due to the obstacles to evaluating parasite viability applying classical cell membrane permeability dyes, as Plasmodium parasites develop inside erythrocytes surrounded by two additional membranes (host cell membrane and parasitophorous vacuole membrane), the mitochondrial membrane potential (ΔΨm), which is disrupted in dying cells, was measured as an indicator of plasmodial viability [20,41,42,43,44]. Some studies have reported that the mechanism of staurosporine-induced cell death involves increased ROS generation [45,46,47] that in turn results in mitochondrial damage and loss of ΔΨm [48]. In accordance with this pathway, disruption of ΔΨm was observed after induction of both apoptotic and necrotic cell death in P. falciparum trophozoites by staurosporine [9,21], a mitochondrial alteration that was also recorded by us in P. vivax and that seemed to be dose-dependent (Figure 1B).
Although the occurrence of RCD processes in malaria parasites is still a matter of debate, studies on P. berghei have demonstrated that ookinetes suffering spontaneous or induced cell death display typical features of apoptosis, including loss of mitochondrial membrane potential, exposure of phosphatidylserine, chromatin condensation, DNA fragmentation, and caspase-like activity [49,50,51]. In P. falciparum, cell death has been mainly explored in blood stages under different in vitro conditions, and apoptosis-like cell death has been fundamentally reported through the detection of DNA fragmentation, as also shown by Arambage and colleagues (2009) [50] for P. falciparum ookinetes. In our study, however, no expressive levels of DNA fragmentation (˂1.5% parasites) were observed in the P. vivax blood stage after incubation, either in the presence or absence of the cell death inducer staurosporine (Figure 2A).
Similar levels of TUNEL-positive parasites and even absence of DNA fragmentation [21,52] have already been found in P. falciparum blood forms stimulated to cell death by different inducers, including staurosporine [20,53], and additional ultrastructural analysis of dying parasites showed no signs of classical apoptotic processes, i.e., early chromatin condensation or nuclear segregation [9,20,21,53,54,55], arguing against deflagration of apoptotic cell death in blood forms of P. falciparum. Indeed, electron microscopy examination has not been routinely applied to the evaluation of P. falciparum apoptosis [14,15,16,17,18,19], and in agreement with the abovementioned works, we observed that staurosporine-induced cell death in P. vivax parasites was not characterized by typical ultrastructural alterations of apoptosis, but by a degenerative cell process with loss of cytoplasmic content (Figure 2B), in which the involvement of parasite digestive vacuoles must still be investigated [55,56]. Thus, it is possible that the TUNEL positivity observed by us resulted from non-specific cell degeneration related to late steps of cell death, as occurs in necrosis, or alternatively that it reflects other cell processes in which the 3’-hydroxyl termini of DNA strands become exposed, as during DNA repair and gene transcription [57]. Above all, it must be considered that the ultrastructural morphology of staurosporine-treated P. vivax herein observed (Figure 2B) was very similar to the necrotic P. falciparum trophozoites induced by staurosporine in a study conducted by Porter and colleagues [21]. In such study, dying parasites with degradation of cytoplasm and absence of typical apoptotic signs were also found under incubation of P. falciparum with heat (40 °C) or chloroquine, supporting necrosis being a common type of cell death in Plasmodium.
It has already been described that cell death in malaria parasites is a Ca^2+^-dependent and transcriptionally regulated process involving the activity of cysteine proteases [9,10,56]. Since caspases are absent in unicellular eukaryotes, the members of the metacaspase family have been implicated in the execution of cell death in protists [58], and in this context, the ability of P. falciparum metacaspase 1 (PfMCA1) to trigger cell death machinery was elegantly demonstrated in a yeast cell model [13]. Therefore, to obtain some insight into the P. vivax death pathway, we evaluated the expression of the pvmca1 gene under cell death stimulation, and no significant change was detected comparing non-treated parasites and parasites treated with staurosporine for 6 h and 18 h (Figure 2C), suggesting that PvMCA1 is not related to cell death signaling in the P. vivax blood stage, at least at the transcriptional level. Of note, a role of metacaspase 2 in the progression of the parasite life cycle was recently shown in P. falciparum and P. berghei [59], and in the fungi models Saccharomyces cerevisiae and Ustilago maydis, metacaspases played a role in the clearance of intracellular insoluble protein aggregates [60,61], raising the possibility of a non-death function of PvMCA1.
Lastly, we examined the impact of dying parasites on the eryptosis of parasitized red blood cells. Eryptosis is an RCD process occurring in red blood cells that can be triggered by diverse stimuli and is characterized by apoptotic hallmarks of nucleated cells, including phosphatidylserine externalization and cell shrinkage [62]. In malaria, increased levels of eryptosis have been described in both parasitized and non-parasitized RBCs, and the contribution of such a cell phenomenon to the pathogenesis of the disease has been considered [30,63]. Interestingly, we observed that induction of cell death in P. vivax trophozoites by staurosporine also positively modulated eryptosis of pRBCs, which was evidenced using annexin V staining and cell size measurement (Figure 3A,B), while no significant alteration in the levels of annexin V-positive nRBCs was detected (Figure 3C).
This observation is in agreement with the well-known capacity of intracellular parasites to manipulate the host cell death pathways [64,65], and most importantly brings additional insight into the parasite–host relationship, in which dying parasites seem to retain the ability to trigger cell death of their host cells, indicating the existence of cross talk between the cell death signaling of these entities. Although the relevance and mechanisms whereby this interconnection takes place are unknown, one can hypothesize that induction of apoptosis/eryptosis in host cells by intracellular dying parasites can operate as an altruistic strategy of immune system evasion, considering that cells undergoing apoptotic processes are endowed with anti-inflammatory properties [66]. Further studies addressing this topic are necessary to better clarify the potential of dying parasites to modulate the host cell death machinery.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1WHO World Malaria Report 2023 WHO Geneva, Switzerland 2023
- 2Price R.N. von Seidlein L. Valecha N. Nosten F. Baird J.K. White N.J. Global Extent of Chloroquine-Resistant Plasmodium vivax: A Systematic Review and Meta-Analysis Lancet Infect. Dis.20141498299110.1016/S 1473-3099(14)70855-225213732 PMC 4178238 · doi ↗ · pubmed ↗
- 3Kaur D. Sinha S. Sehgal R. Global Scenario of Plasmodium vivax Occurrence and Resistance Pattern J. Basic Microbiol.2022621417142810.1002/jobm.20220031636125207 · doi ↗ · pubmed ↗
- 4Chose O. Noël C. Gerbod D. Brenner C. Viscogliosi E. Roseto A. A Form of Cell Death with Some Features Resembling Apoptosis in the Amitochondrial Unicellular Organism Trichomonas vaginalis Exp. Cell Res.2002276323910.1006/excr.2002.549611978006 · doi ↗ · pubmed ↗
- 5Yin J. Howe J. Tan K.S.W. Staurosporine-Induced Programmed Cell Death in Blastocystis Occurs Independently of Caspases and Cathepsins and Is Augmented by Calpain Inhibition Microbiol. Read. Engl.20101561284129310.1099/mic.0.034025-020056704 · doi ↗ · pubmed ↗
- 6Lüder C.G. Campos-Salinas J. Gonzalez-Rey E. van Zandbergen G. Impact of Protozoan Cell Death on Parasite-Host Interactions and Pathogenesis Parasites Vectors 2010311610.1186/1756-3305-3-11621126352 PMC 3003647 · doi ↗ · pubmed ↗
- 7Ni Nyoman A.D. Lüder C.G.K. Apoptosis-like Cell Death Pathways in the Unicellular Parasite Toxoplasma gondii Following Treatment with Apoptosis Inducers and Chemotherapeutic Agents: A Proof-of-Concept Study Apoptosis 20131866468010.1007/s 10495-013-0832-823468121 PMC 3634991 · doi ↗ · pubmed ↗
- 8Menna-Barreto R.F.S. Cell Death Pathways in Pathogenic Trypanosomatids: Lessons of (Over)Kill Cell Death Dis.2019109310.1038/s 41419-019-1370-230700697 PMC 6353990 · doi ↗ · pubmed ↗