Effects of Heat-Induced Oxidative Stress and Astaxanthin on the NF-kB, NFE2L2 and PPARα Transcription Factors and Cytoprotective Capacity in the Thymus of Broilers
Donna Lee Kuehu, Yuanyuan Fu, Masaki Nasu, Hua Yang, Vedbar S. Khadka, Youping Deng

TL;DR
This study examines how astaxanthin protects the chicken thymus from heat-induced oxidative stress and inflammation.
Contribution
The study reveals astaxanthin's potential to mitigate heat stress effects by modulating key gene pathways in the thymus.
Findings
Heat stress activates NF-kB, NFE2L2, and PPARα pathways, increasing inflammation and oxidative stress.
Astaxanthin treatment reduced the negative effects of heat stress on growth performance and gene expression.
Astaxanthin may help restore thymus function by mitigating heat-induced inflammation and oxidative damage.
Abstract
The thymus, a central lymphoid organ in animals, serves as the site for T cell development, differentiation and maturation, vital to adaptive immunity. The thymus is critical for maintaining tissue homeostasis to protect against tumors and tissue damage. An overactive or prolonged immune response can lead to oxidative stress from increased production of reactive oxygen species. Heat stress induces oxidative stress and overwhelms the natural antioxidant defense mechanisms. This study’s objectives were to investigate the protective properties of astaxanthin against heat-induced oxidative stress and apoptosis in the chicken thymus, by comparing the growth performance and gene signaling pathways among three groups: thermal neutral, heat stress, and heat stress with astaxanthin. The thermal neutral temperature was 21–22 °C, and the heat stress temperature was 32–35 °C. Both heat stress…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
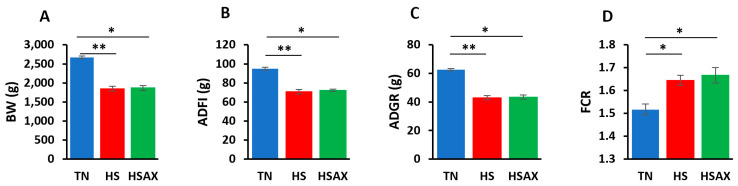 Figure 1
Figure 1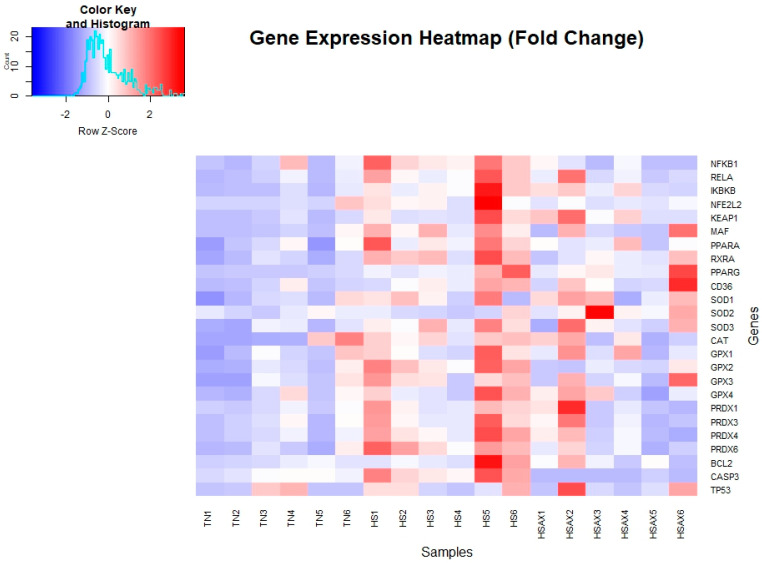 Figure 2
Figure 2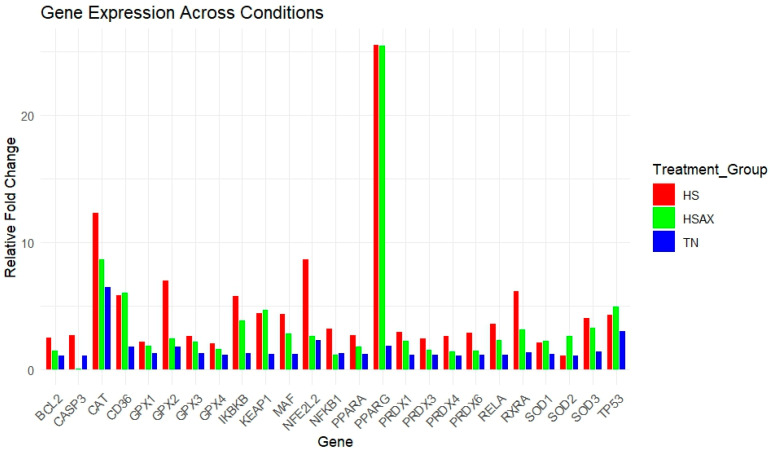 Figure 3
Figure 3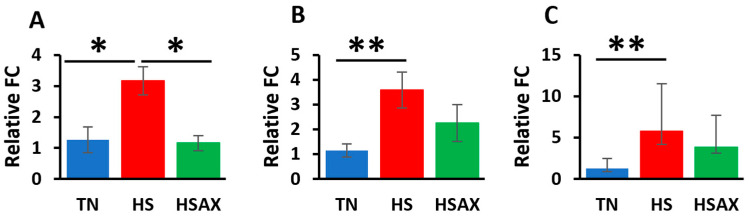 Figure 4
Figure 4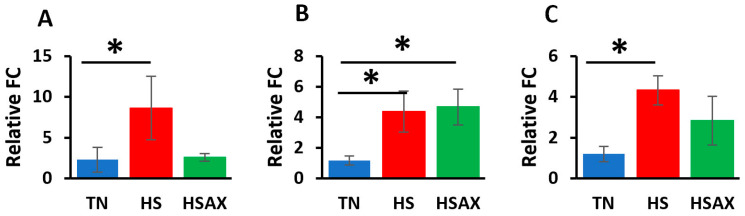 Figure 5
Figure 5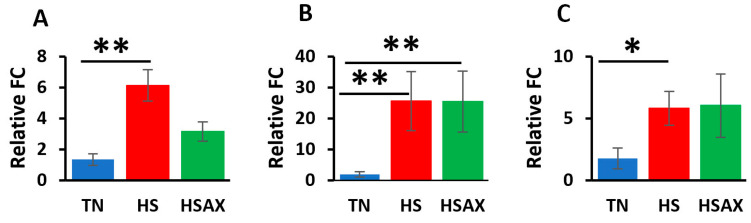 Figure 6
Figure 6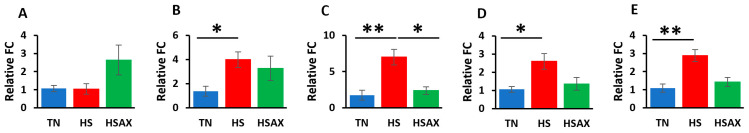 Figure 7
Figure 7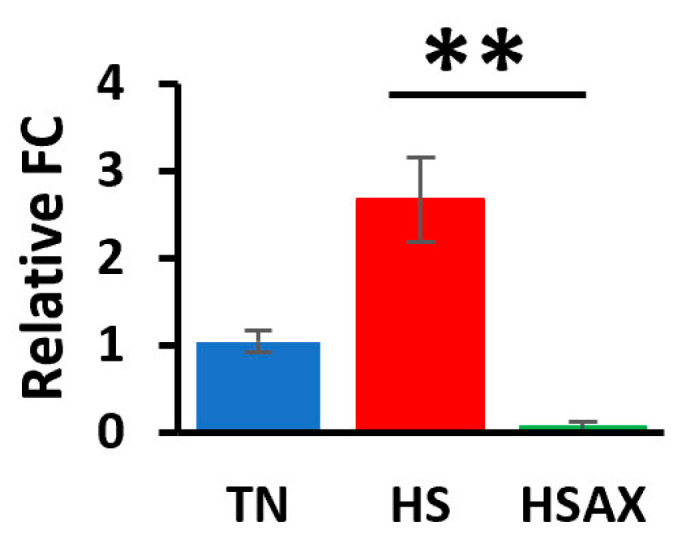 Figure 8
Figure 8- —NIH
- —American Indian Science and Engineering Society Advancing Agricultural Science Opportunities for Native Americans (AASONA)
- —First Nations Development Institute
- —University of Hawaii
- —NSF
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFood Quality and Safety Studies · NF-κB Signaling Pathways · Pediatric health and respiratory diseases
1. Introduction
The thymus is a crucial central lymphoid organ in animals, playing an integral role in the immune response by serving as the site for T cell development, differentiation, and maturation [1,2,3]. These mature T cells subsequently colonize secondary lymphoid organs to combat invading pathogens [4]. Beyond its role in adaptive immunity, the thymus is also a critical mediator of innate immune responses, providing protection against tumors, pathogens, and tissue damage [5]. The integrity of the thymus is essential for maintaining tissue homeostasis and a fully functional immune system. Thymic injury can lead to immune impairment, resulting in significant consequences due to the development of an immature immune system, which can leave the organism immunocompromised [6,7,8].
Stress represents a physiological and biochemical defense mechanism through which the body responds to adverse environmental effects. This response helps the organism adapt to its environment and maintain internal equilibrium [9]. While moderate stress can enhance immunity, excessive stress can negatively impact growth, development, and production performance in animals. Importantly, it can also lead to immune suppression, increasing susceptibility to diseases and potentially resulting in death [10]. Persistent oxidative stress (OS), in particular, impairs immune function through mechanisms such as cellular DNA damage and biomolecule fragmentation [11,12,13]. Normally, the body’s oxidation-antioxidant system maintains a dynamic balance, but OS occurs when the antioxidant (AOX) defenses are overwhelmed [14,15]. This condition can lead to slower growth rates, decreased feed conversion efficiency, reduced production performance, and in severe cases, significant economic losses in industries like poultry farming [16,17]. OS initiates various signaling pathways and inflammatory responses, further compounding its impact on health [18].
The nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB) signaling pathway is a key mediator of OS-induced inflammation [19,20,21,22]. Activation of NF-kB leads to the production of inflammatory cytokines, which are part of the body’s response to harmful stimuli. Inflammation, closely linked to the immune system, is a pathological response to such stimuli [23]. Additionally, the transcription factor nuclear factor erythroid 2-like-2 (NFE2L2/NRF2) plays a significant role in cytoprotection by stimulating the expression of AOX and detoxifying enzymes, including NAD(P)H: quinone oxidoreductase-1 (NQO-1), glutathione S-transferase (GST), and heme-oxygenase-1 (HO-1) [24]. The AOX defense system comprises both enzymatic AOXs, such as superoxide dismutase (SOD), peroxidase (PRDX), catalase (CAT), and glutathione peroxidase (GPX1) and non-enzymatic AOXs, including vitamin E, carotenoids, and vitamin C. An imbalance between oxidative and AOX systems can lead to excessive reactive oxygen species (ROS) and reactive nitrogen species (RNS) accumulation, resulting in the destruction of lipids, proteins, and nucleic acids, ultimately causing cellular damage and cell death. OS is thus implicated in a variety of pathological conditions, including inflammatory reactions and tumor development [25,26,27]. Peroxisome proliferator-activated receptor alpha (PPARα) is a nuclear receptor that regulates the genes involved in lipid metabolism, fatty acid oxidation, and inflammation and plays a crucial role in maintaining metabolic homeostasis. PPARα has been shown to produce significant anti-inflammatory effects [28,29] and inhibit the activation of NF-kB [30].
Heat-induced OS poses a particular challenge in animals, affecting their ability to metabolize carotenoids, which necessitates dietary supplementation [31]. Astaxanthin (AST), a potent lipid-soluble AOX from the carotenoid family, has demonstrated exceptional free radical scavenging and anti-inflammatory properties [32,33,34,35]. Its unique structure, featuring hydroxyl and keto groups on each ionone ring, enhances its AOX activity, thereby protecting cellular membranes from oxidation [36]. Despite its presence in aquatic organisms and birds, AST cannot be synthesized by animals and must be obtained from dietary sources such as green algae, red yeast, and crustacean byproducts, with Haematococcus pluvialis being a particularly rich source [37,38]. The carotenoid antioxidant value of powder AST is described as the Oxygen Radical Absorbance Capacity (ORAC) and expressed in micromoles of Trolox equivalents per 100 g (μmol TE/100 g), a vitamin E analog. AST has been shown to have 6000 times the AOX capacity of vitamin C, 800 times that of CoQ10, and 550 times that of vitamin E [39]. Supplementation with AST has shown numerous health benefits, including anti-inflammatory, immunomodulatory, cardiovascular, neuroprotective, and anticancer effects [40]. In poultry farming, particularly under heat-stress conditions, AST holds promise as a natural AOX feed supplement, potentially mitigating the adverse effects of heat stress and improving overall health and productivity.
The main objectives of the present study were to investigate the protective properties of AST against heat-induced OS and apoptosis in the chicken thymus. Our research question is as follows: “How does heat-induced OS affect the NF-kB, NFE2L2, PPARα, cytoprotective capacity genes and apoptotic pathways in the broiler thymus, and can AST help mitigate the stress?” We hypothesize that heat-induced OS in the broiler thymus correlates with poor growth performance, disrupting the physiological and biochemical defense mechanisms, and that H. pluvialis-derived AST dietary supplementation may play a role in mitigating the effects through modulation of transcription pathways.
2. Materials and Methods
2.1. Ethics Statement
The animal protocol (Protocol No. 17-2605) used in this study was approved by the University of Hawaii Institutional Animal Care and Use Committee (IACUC). Animals were raised under animal welfare guidelines and euthanized in accordance with humane protocols in preparation for necropsy.
2.2. Experimental Animal Design
Cobb 500 unsexed broiler chicks were obtained from Asagi Hatchery (Honolulu, HI, USA). Asagi Hatchery is a local commercial producer that sells newly hatched chicks for commercial or research purposes; we have permission to utilize the resource they provide without a need for written consent, but we acknowledge them as a source. Several mitigation strategies were simultaneously tested in parallel, and the findings were reported separately based on treatment. In this trial, the feed additive treatment is AST. One-day-old chicks were weighed, wing tagged, and then separated into groups of 4 birds in each pen, (n = 60). The pens were established on concrete flooring, prepared with pine shavings, and stocked with water bottles and open feeders. Animals were provided a normal starter feed from 0 to 21 days, and a normal finisher feed on 22–42 days with free access to feed and water. Nutritional compositions of the supplemented diets are listed in Appendix A (Table A1). The animals were reared under two temperature conditions: thermal neutral (TN) (n = 24) at 21–22 °C and 50% RH, and heat stress (HS) (n = 36) at 32–35 °C and 42–50% RH. In the first 21 days, all birds were raised under the TN temperature conditions and fed the same diet. In the last 21 days, the two groups raised under the HS temperature conditions were further divided into two dietary regimens, i.e., basal diet HS (n = 18, treatment 1), and basal diet with 1.33 mg/kg AX supplement (HSAX) (n = 18, treatment 2). The light cycle was set at a 1:23 dark:light cycle throughout the trial. Six broiler hens were randomly selected from each group and euthanized on day 42, and thymus tissue samples were collected at necropsy.
2.3. Astaxanthin-Rich Dietary Supplement
The diet was supplemented with P25HB provided by AstaReal^®^, Inc. (Burlington, NJ, USA). PH25B contains 2.5% (w/w) dried Haematococcus pluvialis algae as well as other components including modified starch, gum Arabic, mixed tocopherols, L-ascorbyl palmitate, silicon dioxide, xanthan gum, γ–cyclodextrin, polysorbate 80, rosemary extract and ferulic acid. A comparable nutritional composition of H. pluvialis algae is listed in Appendix A (Table A2). The natural forms of astaxanthin mainly comprise mono-esterified, followed by di-esterified and free forms 3,3′-dihydroxy-β, β-carotene-4 and 4′-dione (C_40_H_52_0_4_ free form). Fuji Health Sciences, Inc. (AstaReal Inc., Moses Lake, WA, USA), AstaReal ORAC value is reported at 2,822,200 μmol TE/100 g, supported by Non-US Gov’t: Brunswick Laboratories Test Report Batch No. B-10267b-2010.
2.4. Growth Indicators
Weekly feed intake was recorded, and the average daily feed intake (ADFI), average daily gain ratio (ADG), and feed conversion ratio (FCR) were calculated. The body weight (BW) of each bird was recorded using a Mettler Toledo scale before heat stress treatment and at the end of the heat treatment.
2.5. Tissue Sample Collection
Immediately after euthanizing, thymus tissue samples were collected from 6 randomly selected birds from each group, which were snapped-frozen in liquid nitrogen and stored at −80 °C.
2.6. Total RNA Extraction and cDNA Preparation
Total RNA was isolated from frozen tissues (50–100 mg) using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. The concentration of total RNA was determined using a NanoPhotometer^®^ P330 (IMPLEN, Los Angeles, CA, USA). Complementary DNA (cDNA) was synthesized from 1 µg of total RNA (20 µL reaction of RT mixture) using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA) and further diluted with nuclease-free water (1:25) for the qPCR reaction outlined below.
2.7. Bioinformatics: Genome Assembly and Gene Primer Design
The National Center for Biotechnology Information (NCBI) genome browser was used to search and compile genes for Gallus gallus domesticus related to heat stress, oxidative stress, cytoprotective, epithelial integrity, transcription factors and housekeeping genes. The NCBI-Basic Local Alignment Search Tool (BLAST) was used to design primers for polymerase chain reaction (PCR) from the accession numbers obtained from the list of genes presented in Appendix A (Table A3). The primer parameters were set for a PCR product size between a minimum of 100 and maximum of 250 for 5 primers to return. The primer melting temperatures were set for a minimum of 55 °C, optimum of 57 °C, and maximum of 60 °C with a maximum Tm difference of 3 °C. The exon junction span was set so that the primers must span an exon–exon junction. The organism specified was Gallus gallus (taxid 9031). The forward and reverse primer sequences (5′→ 3′) were then submitted to Integrated DNA Technologies (Coralville, IA, USA) for synthesis.
2.8. Quantitative Real-Time RT-PCR (qPCR)
The qPCR was performed using PerfeCTa SYBR Green FastMix (Quantabio, Beverly, MA, USA) on a Q—qPCR instrument (Quantabio, Beverly, MA, USA). The qPCR reaction mixture consisted of 2 µL of cDNA, 10 µL PerfeCTa SYBR Green FastMix, 1 µL of each forward and reverse primer (5 µmol concentration), and 6 µL of sterile deionized water to make a final reaction mixture of 20 µL. Specific primer pairs for the detection of each gene were designed using the NCBI Primer-Blast tool. The qPCR reaction was carried out following the standard cycling mode. The amplification conditions were 50 °C for 2 min (hold) and then 95 °C for 2 min (hold) followed by 40 repeat cycles of 95 °C for 15 s (denaturation), 60 °C for 15 s (annealing), and 72 °C for 1 min (extension). A melting curve was also generated to confirm SYBR Green-based objective amplicon, and further qPCR products were confirmed using 2% agarose gel electrophoresis. Three house-keeping genes, glyceraldehyde 3-phosphate dehydrogenase (GAPDH), beta-actin (ACTB), and TATA-Box Binding Protein (TBP), were analyzed in triplicates in each bird to determine the most stable house-keeping gene. Based on the uniformity of the expression level across samples, ACTB was chosen as the housekeeping gene. The gene expression level was determined using cycle threshold (Ct) values following the standard curve method after normalization with housekeeping genes. The fold change for each gene was calculated using the 2^−ΔΔCt^ method and presented as mean ± standard error [41].
2.9. Gene Ontology
Significantly differentially expressed genes identified from the qPCR procedure were searched in the Ensembl genome database for chicken (GRCg6a) species to obtain gene ontology (GO) information (https://www.ensembl.org/index.html, accessed on 2 April 2024) [42]. The GO information included the cellular component, molecular function and biological process of these genes identified with ENSGAL Transcript IDs.
2.10. Statistical Analysis
Statistical analysis was performed using the Kruskal–Wallis rank sum test with statistical significance set at p < 0.05, followed by the Dunn post hoc test for comparison between three groups: TN, HS, and HSAX, and the p-value adjusted using the Bonferroni method. Growth performance measurements were calculated based on the data collected at the end of the 42-day trial period. Analysis was conducted using the R open source program, libraries ‘FSA’, ‘dunn.test’, and ‘gplots’, R Core Team (2023) (https://www.R-project.org/ accessed on 2 April 2024) [43].
3. Results
3.1. Growth Performance
At the end of the 42-day poultry trial, the TN group growth performance indicators were found to be significantly higher for BW, ADFI, and ADG compared to the HS and HSAX groups, as established through a Kruskal–Wallis test to evaluate differences among the three groups (p < 0.01). Although the results clearly showed negative impacts of heat on the HS and HSAX groups, when performing the post hoc pairwise comparisons using Dunn’s test and applying the Bonferroni correction to control increased risk of error due to multiple comparisons, the negative impacts were found to be more significant in the HS group compared to the TN (p < 0.01), whereas the negative impact was slightly less in the HSAX group (p < 0.05). In addition, the FCR showed that the TN group was significantly lower in feed conversion, requiring less feed to maintain body weight compared to the HS and p < 0.05 (Table 1 and Figure 1).
3.2. Quantitative Real-Time RT-PCR (qPCR) Gene Expression
For gene expression studies of the thymus, three genes were considered for housekeeping genes: GAPDH, ACTB, and TBP. ACTB was selected for its high and relatively stable expression under the experimental conditions performed, making it a reliable reference for normalizing the gene expression data collected. Kruskal–Wallis rank sum test with statistical significance set at p < 0.05 was conducted comparing the three groups: TN, HS, and HSAX (Table 2). A gene expression heat map of all samples profiling the visual aspect of the fold change data is displayed in Figure 2. A histogram of gene expression of relative fold change levels across all genes provides a summary comparison between the treatment groups (Figure 3).
3.2.1. NF-kB Transcription Signaling Pathway Genes
The studies showed the impact of HS and AOX treatment expressed through the NF-kB transcription factor signaling pathway and pairwise comparisons (Figure 4). NFKB1 (p = 0.028), NF-kB subunit (RELA) (p = 0.009), and inhibitor of nuclear factor kappa B kinase subunit beta (IKBKB) (p = 0.011) were found to be more upregulated in the HS group in comparison to the TN group. In addition, the expression of NFKB1 (p = 0.028) was found to be more upregulated in the HS group than in the HSAX group.
3.2.2. NFE2L2-Mediated Signaling Pathway Genes
The results of the impact of HS on the gene expression of the NFE2L2-mediated signaling pathway showed that, compared to the TN group, the HS treatment group demonstrated higher upregulation of NFE2L2 (p = 0.033), kelch-like ECH-associated protein 1 (KEAP1) (p = 0.028), and musculoaponeurotic fibrosarcoma (MAF) (p = 0.024) (Figure 5). The expression of KEAP1 (p = 0.028) was also more upregulated in the HSAX group compared to the TN group.
3.2.3. PPARα Signaling Pathway Genes
Consistent with the findings for the NF-kB and NFE2L2 pathways, the PPARα pathway showed similar results with the HS group showing higher upregulation in terms of the gene expressions of retinoid X receptor alpha (RXRA) (p = 0.003), peroxisome proliferator-activated receptor gamma (PPARγ) (p = 0.011), and cluster of differentiation 36 (CD36) (p = 0.045) compared to the TN group (Figure 6). The HSAX group also showed higher upregulation of PPARγ (p = 0.011) compared to the TN group.
3.2.4. Cytoprotective Capacity Genes
Results of the cytoprotective capacity genes provided similar results to those of the previously examined transcription factor pathways where the HS group showed higher upregulation than the TN group for the following expressions: superoxide dismutase 3 (SOD3) (p = 0.039), glutathione peroxidase 2 (GPX2) (p = 0.006), peroxiredoxin 4 (PRDX4) (p = 0.028), and peroxiredoxin 6 (PRDX6) (p = 0.006) (Figure 7). In addition, the HS group was found to be upregulated higher than the HSAX group in the expression of GPX2 (p = 0.0448). Interestingly, although the Kruskal–Wallis test for SOD2 was shown to be significant, the post hoc test did not indicate significance in the comparisons between the groups.
3.2.5. Apoptotic Gene
The results of the apoptotic pathway showed a clear indicator in the upregulation of the HS group over the HSAX group in the expression of caspase 3 (CASP3) (p = 0.001) (Figure 8).
4. Discussion
4.1. Growth Performance
Growth performance is anticipated to suffer under any stress condition and was confirmed in this study under HS [44]. Reduction in voluntary feed intake, decreased energy availability, altered nutrient digestibility and metabolism, and the breakdown of intestinal epithelial structure and function are some direct negative impacts of heat stress on poultry behavior and physiology [45]. Although the growth performance indexes did not show any significant benefit from an AST supplement based on BW measurements related to feed intake and weight gain, the health of the poultry was further elucidated through the molecular mechanisms of gene expression providing insights affecting the health and wellbeing of the broilers.
4.2. Gene Ontology Enrichment and Expression Analysis
4.2.1. NF-kB Transcription Signaling Pathway Genes
The NF-kB transcription signaling pathway is a critical regulator of immune and inflammatory responses. NF-kB can be found throughout the cell in the nucleus, mitochondrion, chromatin and cytoplasm. When not upregulated, NF-kB is generally sequestered in the cytoplasm by IkB inhibitor proteins such as IKBKB, located in the cytoplasm and cytosol. RELA found in the cytoplasm is a key subunit of the NF-kB transcription factor complex, needed to form heterodimers of NF-kB for translocation into the nucleus upon activation for DNA-binding transcription activity. When activated by stimuli such as stress, free radicals, pathogens and cytokines, IkBs are phosphorylated by a kinase enzyme for ubiquitination and degradation, freeing the NF-kB to translocate into the nucleus and promote the transcription of target genes involved in inflammation, immune response, cell proliferation, and survival. The NF-kB pathway plays a significant role in responding to cellular stress and homeostasis Appendix A (Table A4) [46].
The outcome of our studies indicates that the upregulation of the HS group over the TN group is consistent with the function of the NF-kB complex to respond to cellular stress, and the administration of AST had an attenuating effect. HS exposure has been found to increase NF-kB p65 mRNA expression in the spleen of the broiler chicken but inhibited the activation of NF-kB p65, reducing the inflammatory response with dietary-seaweed-derived polysaccharides [47]. While there is no significance between the TN and HSAX groups, it indicates that AST has some effect in reducing cellular stress, which coincides with the results in which there is a significant difference between the upregulation of the HS over the HSAX group.
4.2.2. NFE2L2-Mediated Signaling Pathway Genes
The NFE2L2 (NRF2) plays a crucial role in cellular defense against OS. NFE2L2 is found throughout the cell in the nucleus, Golgi apparatus, chromatin, plasma membrane and cytoplasm. Under normal conditions, NFE2L2 is bound by KEAP1 in the cytoplasm, and in response to OS or electrophilic stimuli, NFE2L2 is released to translocate to the nucleus, where it heterodimerizes with small MAF proteins. This NFE2L2-MAF complex binds to antioxidant response elements (AREs) in the promoters of target genes, leading to transcription of various cytoprotective genes (see Appendix A (Table A4)) [46,48].
The results of our study show that the HS group was more upregulated in comparison to the TN group in response to OS throughout the NFE2L2 signaling pathway. NFE2L2 is expressed in many tissues and plays a crucial role in combating oxidative stress. Under stressful conditions, NFE2L2 disassociates from KEAP1, leading to increased NFE2L2 expression levels and enhancing the cell’s antioxidant capacity, which was demonstrated in liver tissue and plasma of broilers subjected to heat stress and supplemented with dietary glutamine [49]. HSAX is upregulated over the TN group in the KEAP1 gene expression, a critical regulator of the NFE2L2 signaling pathway, acting as a sensor for OS and controlling the activity of NFE2L2, likely a homeostatic feedback mechanism to maintain a balanced redox environment.
4.2.3. PPARα Signaling Pathway Genes
The PPARα signaling pathway plays a vital role in lipid metabolism, energy homeostasis and inflammation. Although PPARα**,** which is predominantly found in the nucleus, has only marginal significance in the upregulation of the HS over the TN group, RXRA, PPARγ and CD36 are significantly expressed. PPARγ in the HSAX group is also more upregulated in comparison to the TN group. PPARα is generally in an inactive state in the nucleus and is activated by fatty acids or specific ligands to form a heterodimer with RXRA, and then further undergoes conformational changes for DNA-binding transcription factor activity in the promoter of target genes. Among the target genes is CD36**,** which facilitates fatty acid uptake and enzymes for lipid metabolism. PPARγ can interact with PPARα to help regulate lipid metabolism, reduce inflammation and maintain energy homeostasis (see Appendix A (Table A4)) [50].
The results from our study indicate that although there is a subtle response of PPARα to heat stress, PPARγ appears to be involved in the cellular response to heat stress, suggesting a role in AOX-mediated response to stress. The PPARα pathway is involved with lipid metabolism and fatty acid oxidation, where serum malondialdehyde (MDA) has been shown to significantly increase lipid peroxidation due to heat stress in broilers. Spirulina platensis supplementation modulated the negative impact of heat stress by reducing the MDA concentration [51]. RXRA forms heterodimers with PPARα and is essential for transcriptional activity. The upregulation of CD36 in the HS group compared to the TN group suggests increased lipid metabolism or fatty acid utilization, placing demand on energy due to heat-induced OS.
4.2.4. Cytoprotective Capacity Genes
Cytoprotective capacity genes safeguard cells from various stressors including oxidative damage by enhancing AOX defense mechanisms to promote cell survival. SOD2 is found in the mitochondrion and responds to OS through the oxidation-reduction process for the removal of superoxide radicals. SOD3 is found in the extracellular space and plays a crucial role in scavenging superoxide radicals outside the cell. GPX2 is found in the cytosol, intercellular bridge and mitotic spindle, involved in detoxifying peroxides, such as hydrogen peroxide. PRDX4 is found in the cytoplasm and endoplasmic reticulum, and PRDX6 is also found in the cytoplasm, as well as the nucleus. Both peroxiredoxins are involved in scavenging for peroxides, providing peroxide detoxification and protection of cells from OS (see Appendix A (Table A4)) [52,53].
The results showed significant upregulation of the HS compared to the TN groups in the expression of SOD3, GPX2, PRDX4 and PRDX6. There was also significant upregulation of the HS group over the HSAX group for GPX2, but only marginally for PRDX6. In addition, when analyzing the expression of SOD3, there was marginal upregulation of HS over the HSAX group. Related results were produced in a study of meat quality of heat-stressed broilers provided with various supplementation levels of AST for AOX efficacy. In this study, the liver tissue showed an increase in gene expression of the heat-stressed birds for HSP27 and HSP70**,** which scavenge for free radicals and assist in proper protein folding, respectively, to reduce oxidative damage. Conversely, AST treatment lowered the gene expression, which may reflect the anti-stress effects of AST and may be due to AOX effects that reduce the adverse impact of heat stress [54]. A separate study on layer hens exposed to heat stress and supplemented with AST resulted in liver tissue showing decreased GPX and enzyme activities of glutathione peroxidase and glutathione S-transferase, suggesting that AST neutralizes free radicals directly or indirectly by removing harmful substrates [55]. On the contrary, a related study conducted on the mRNA expression of ileum tissue of the same subject heat-stressed broilers treated with AST resulted in the HSAX group being significantly upregulated over the HS for HSF2, SOD2, and TXN genes and HSAX upregulated over TN for GPX3. The overall outcome was improved epithelial integrity with the upregulation of LOX, CLDN1 and MUC2 genes in the HSAX group [45]. Similarly, broilers subjected to heat stress resulted in the duodenum tissue mRNA expression of Nrf2, HO-1, GPx1 and GSTT1 being downregulated by HS, and when provided a dietary supplementation of algae-derived polysaccharides, the mRNA expression was upregulated [56]. This indicates a distinct difference between organs with different structures, functions and cell types. Overall, our findings highlight the dynamic regulation of AOX genes in response to heat-induced OS and the potential modulatory effects of AST-AOX supplementation on cellular AOX defense mechanisms.
4.2.5. Apoptotic Pathway Genes
The CASP3 gene can be found in the cytoplasm and nucleus, where it encodes for a cysteine protease involved in the execution phase of apoptosis. BCL2 encodes for an anti-apoptotic protein that inhibits cell death, and TP53 encodes for a tumor suppressor protein involved in cell cycle regulation and apoptosis (see Appendix A (Table A4)) [53,57].
Our findings show significant upregulation of CASP3 in the HS group over the HSAX group, which suggests the activation of apoptotic pathways, potentially indicating cellular damage or heat-induced OS. Studies using algae-derived polysaccharides to attenuate heat stress in the duodenum and spleen resulted in an increase in the apoptosis rate following heat stress and a decrease in the apoptosis rate after supplementation [47,56]. The lack of significant expression of BCL2 and TP53 may suggest that heat stress alone may not induce significant alterations in those apoptotic regulators. However, it appears that the significant reduction in CASP3 in the HSAX group seems to indicate a protective factor from apoptosis due to the AST supplementation, which may play a role in mitigating the response to heat stress.
5. Conclusions
Overall, our experimental findings highlight the dynamic regulation of gene expression in the thymus related to the NF-kB, NFE2L2, PPARα, cytoprotective capacity and apoptotic pathways. Our main objective of the present study was to investigate the protective properties of AST against heat-induced OS and apoptosis in the chicken thymus. Our research provided insights into the molecular regulatory mechanisms that respond to heat-induced OS, and the potential therapeutic implementation of AST-AOX supplementation to mitigate the effects through the modulation of transcription pathways. The complexities of such mechanisms from different tissue types and the varied responses of AOX, whether endogenous or applied, leave us with knowledge gaps that require further research to understand the therapeutic potentials of AST-AOX and investigate the effects of dosing.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Vos J.G. Moore J.A. Immune suppression as related to toxicology CRC Crit. Rev. Toxicol.197756710110.3109/1040844770910134217515 · doi ↗ · pubmed ↗
- 2Murray J.M. Kaufmann G.R. Hodgkin P.D. Lewin S.R. Kelleher A.D. Davenport M.P. Zaunders J.J. Naive T cells are maintained by thymic output in early ages but by proliferation without phenotypic change after age twenty Immunol. Cell Biol.20038148749510.1046/j.1440-1711.2003.01191.x 14636246 · doi ↗ · pubmed ↗
- 3Gameiro J. Nagib P. Verinaud L. The thymus microenvironment in regulating thymocyte differentiation Cell Adhes. Migr.2010438239010.4161/cam.4.3.1178920418658 PMC 2958614 · doi ↗ · pubmed ↗
- 4Savino W. The thymus is a common target organ in infectious diseases P Lo S Pathog.20062 e 6210.1371/journal.ppat.002006216846255 PMC 1483230 · doi ↗ · pubmed ↗
- 5Thapa P. Farber D.L. The role of the thymus in the immune response Thorac. Surg. Clin.20192912313110.1016/j.thorsurg.2018.12.00130927993 PMC 6446584 · doi ↗ · pubmed ↗
- 6Raviola E. Karnovsky M.J. Evidence for a blood-thymus barrier using electronopaque tracers J. Exp. Med.197213646649810.1084/jem.136.3.4664115129 PMC 2139259 · doi ↗ · pubmed ↗
- 7Lynch H.E. Goldberg G.L. Chidgey A. Van den Brink M.R.M. Boyd R. Sempowski G.D. Thymic involution and immune reconstitution Trends Immunol.20093036637310.1016/j.it.2009.04.00319540807 PMC 2750859 · doi ↗ · pubmed ↗
- 8Dai X. Zhang D. Wang C. Wu Z. Liang C. The pivotal role of thymus in atherosclerosis mediated by immune and inflammatory response Int. J. Med. Sci.2018151555156310.7150/ijms.2723830443178 PMC 6216065 · doi ↗ · pubmed ↗