Multi-Omics Revealed Resveratrol and β-Hydroxy-β-methyl Butyric Acid Alone or in Combination Improved the Jejunal Function in Tibetan Sheep
Qiurong Ji, Fengshuo Zhang, Yu Zhang, Quyangangmao Su, Tingli He, Shengzhen Hou, Linsheng Gui

TL;DR
This study shows that resveratrol and HMB, alone or together, improve gut health in Tibetan sheep by boosting antioxidants, immunity, and gut bacteria.
Contribution
The study reveals how resveratrol and HMB improve jejunal function in ruminants through multi-omics analysis of microbiota and metabolites.
Findings
RES and HMB supplementation increased antioxidant enzymes and immune markers in Tibetan sheep.
Jejunal morphology improved with RES and HMB, including villus height and muscle layer thickness.
RES-HMB increased butyrate concentration and beneficial microbes like Methanobrevibacter and Bacillus.
Abstract
Previous research studies confirmed that both resveratrol (RES) and β-hydroxy-β-methyl butyric acid (HMB) improved growth performance by altering intestinal microbiota. However, the mechanism underlying of RES and HMB on intestinal function remains unclear in ruminant. In this study, supplements of RES and HMB alone or in combination were evaluated as promoters of antioxidant capacity, immune response and barrier function, and modulators of the microbiota and metabolite profiles in the jejunum of Tibetan sheep. A total of 120 two-month-old Tibetan rams were randomly divided into four treatments (n = 30 per treatment), which were supplemented with a basal diet with 1.5 g RES/d (RES group), 1.25 g HMB/d (HMB group), 1.5 g RES/d plus 1.25 g HMB/d (RES-HMB group), and without additions (Control group). The results showed that RES and HMB improved the antioxidant capacity (CAT, GSH-Px, SOD,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
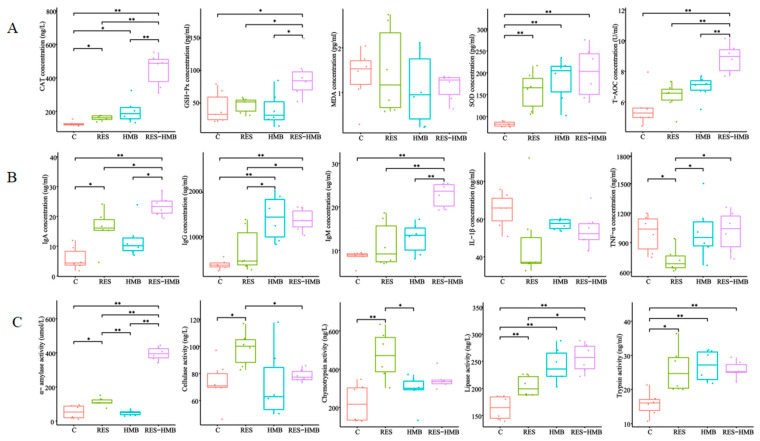 Figure 1
Figure 1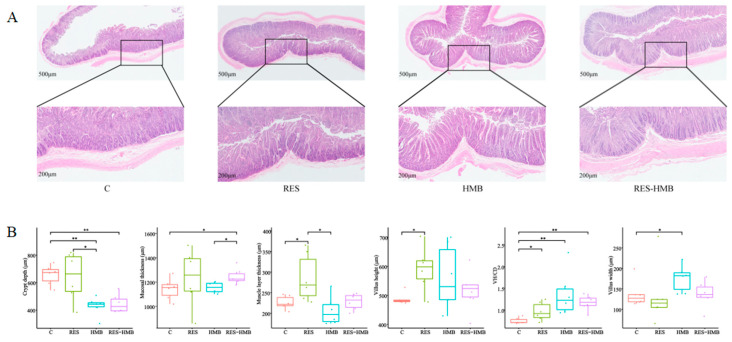 Figure 2
Figure 2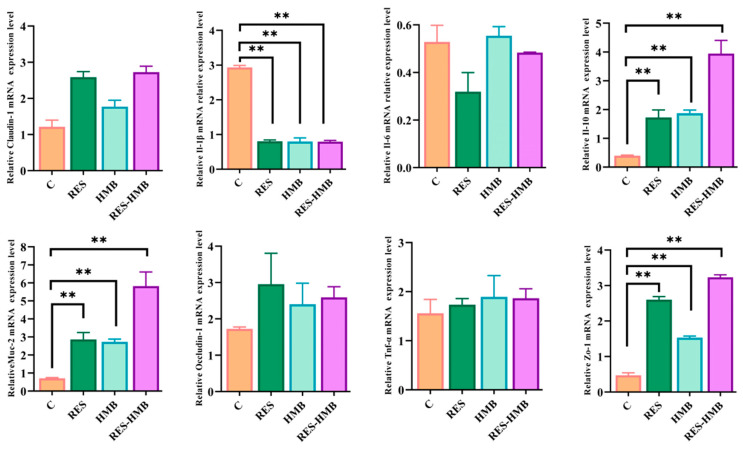 Figure 3
Figure 3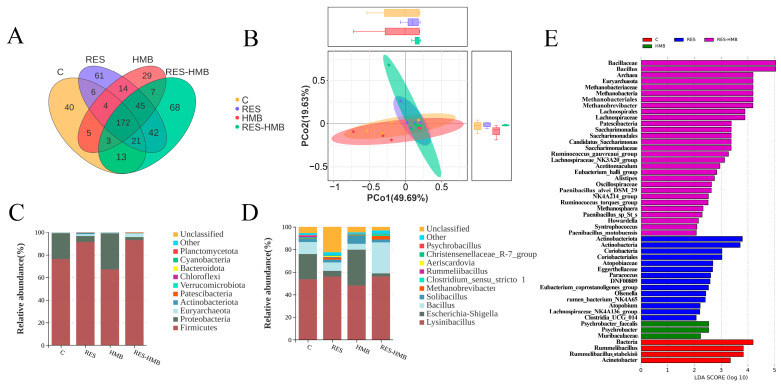 Figure 4
Figure 4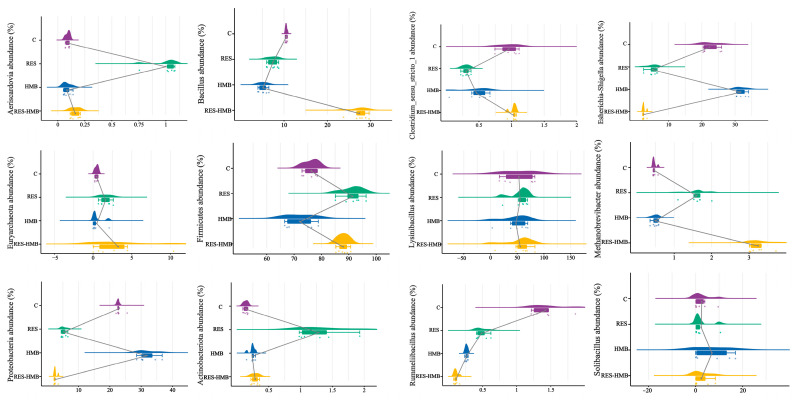 Figure 5
Figure 5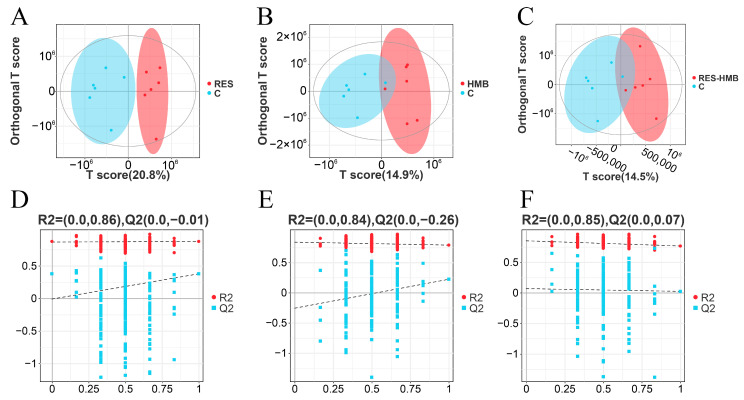 Figure 6
Figure 6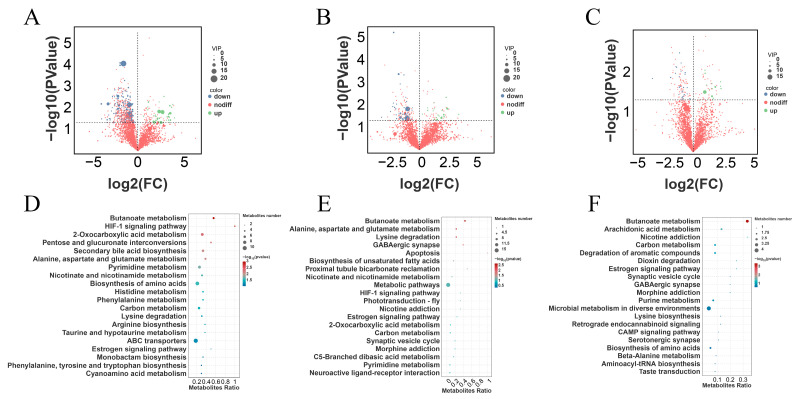 Figure 7
Figure 7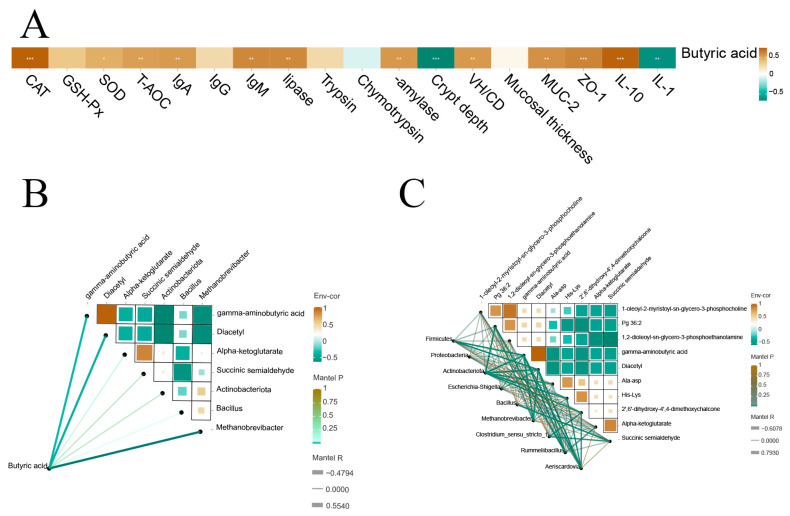 Figure 8
Figure 8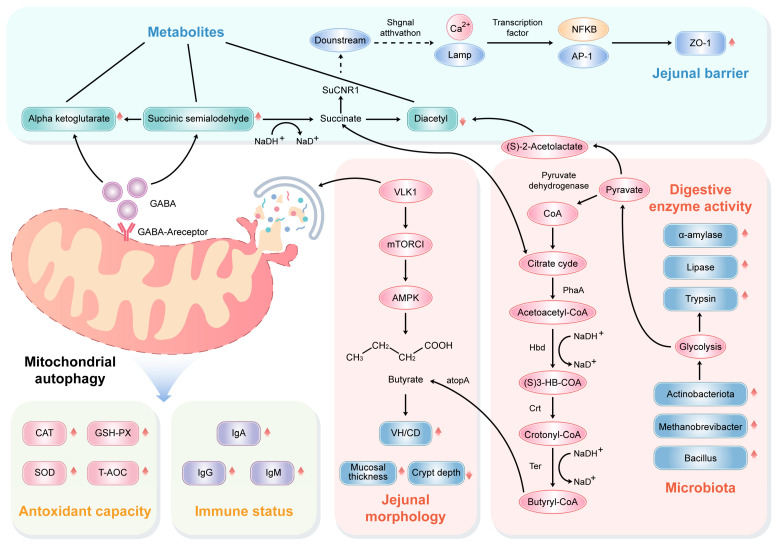 Figure 9
Figure 9- —Construction of Standardized Production System for Improving quality and efficiency of Tibetan sheep industry
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFrench Historical and Cultural Studies
1. Introduction
As a selectively permeable barrier, the intestine serves the absorption of nutrients including organic acids, electrolytes, and water, as well as protecting against food-borne pathogens [1]. Within the intestinal tract of mammals exists abundant and diverse communities of symbiotic microbes [2]. The gut microbiome is a widely explored diverse population and participates in maintaining metabolism, stabilizing immune function, and resisting pathogens in the health of the host [3]. Research has demonstrated that regulating animal feed can affect the composition, abundance, and metabolites of gut microbiota, thereby manipulating animal health [4]. Therefore, research into substitutes such as lysozyme, essential oils, bacteriophages, and prebiotics is crucial [5].
Resveratrol (RES) is a polyphenolic plant antitoxin produced by plants in response to environmental stress, present in at least 72 plants, commonly found in grapes, peanuts, and berries [6]. This plant antitoxin exhibits a wide range of biological properties such as antioxidant [7], anti-saccharification [8], anti-aging [9], anti-inflammatory [10], anti-cancer [11], and neuroprotective properties [12]. Previous studies indicated that the gut microbiota maintained redox homeostasis by regulating the production of reactive oxygen species (ROS) and RES served as a scavenger for scavenging free radicals such as ROS. Given the bidirectional relationship between the two terms, it is hypothesized that the gut microbiota may serve as a target for resveratrol to maintain intestinal homeostasis [13]. In an early study of a dextran sodium sulfate (DSS) rat colitis model, adding RES to the diet repaired the colon barrier and reversed the ecological imbalance of microorganisms [14]. Upon arterial remodeling, RES regulated the microbiota-derived metabolites (e.g., short-chain fatty acids) [15].
β- Hydroxyl group- β- Methyl butyric acid (HMB) is an organic acid containing five carbons, which is produced by leucine [16], and promotes protein synthesis [17]. According to the report, 0.10% HMB supplementation altered the abundance, diversity, and composition of gut microbiota in broilers, thereby inhibiting liver fat deposition [18]. Studies have also shown that as a probiotic formulation, HMB can utilize its mechanism to reprogram gut microbiota and metabolism to reverse HFD-induced obesity [19]. However, the endogenous production of HMB is very low, and the amount that can be converted into HMB from the normal intake of leucine does not exceed 10%. Therefore, it is necessary to supplement HMB with a diet to fully exert its effects [20].
However, there have not been reports on the study of the dietary supplementation of RES and HMB in ruminant. It is hypothesized that RES and HMB could have a positive effect on the microbiota and metabolome, thereby supporting intestinal function. Therefore, the aims of present study to explore the mechanism of RES and HMB alone or in combination on the jejunal function using 16s RNA gene sequencing and non-targeted metabolomics in Tibetan sheep.
2. Materials and Methods
2.1. Ethical Statement
The protocol and methodology of the present study were approved by the Institution of Animal Care and Use Committee at Qinghai University, China (Xining, China; Permit No. QUA-2020-0709).
2.2. Experimental Design
The Tibetan rams were selected from Qinghai Xiangkameiduo animal husbandry Co., Ltd. (Gonghe, Qinghai province, China). All individuals were immunized by a standardized procedure (e.g., ovine braxy, struck, lamb dysentery, enterotoxemia, and ovinia) and weaned at 60 d of age. In this experiment, one hundred and twenty Tibetan rams aged 2 months old (initial weight: 15.5 ± 0.14 kg) were clinically evaluated and were apparently healthy, and they were randomly divided into 4 treatments optionally. The four treatments were supplemented with a basal diet with 1.5 g RES/d (RES group), 1.25 g HMB/d (HMB group), 1.5 g RES/d plus 1.25 g HMB/d (RES-HMB group), and without additions (Control group), respectively. Each treatment consisted of 5 replicate pens with 6 rams. The feeding trial continued for 100 days, including a 10-day acclimation period and a 90-day experimental period. All of the sheep had access to fresh water ad libitum. The total mixed ratio contained 70% concentration and 30% forage (dry matter basis). The diet composition and nutrient levels are listed in Table 1.
2.3. Sample Collection
At the end of the experiment, twenty-four Tibetan lambs (n = 6 per treatment) were slaughtered at a commercial slaughterhouse. The jejunal contents were collected and stored at −80 °C for omics analysis. Synchronously, the jejunal tissues were collected and fixed with 4% paraformaldehyde for histological analysis.
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
The jejunal contents were centrifuged (2500× g) for 15 min at 4 °C. The indicators of supernatant including antioxidant capacity, immune response, and digestive enzyme activity were determined using ELISA kits (Jiangsu Meimian Industrial, Yancheng, Jiangsu, China).
2.5. Jejunal Morphology
The fixed jejunal tissues were embedded in paraffin and cut to 3 µm. Five discontinuous sections of each sample were selected for hematoxylin–eosin staining. Stained sections were observed using an Olympus BX51 microscope. The villus height, villus width, mucosal thickness, muscularis propria thickness, crypt depth, and villi height/crypt depth (VH/CD) were measured by Imagingeproplus 6.0 analysis software.
2.6. Quantitative PCR (qPCR)
The total RNA of jejunal tissues was extracted using Transzol Up (TRAN, Beijing, China) and reverse transcribed using the Universal SYBR Green qPCR Mix kit (Azaood, Beijing, China). The qPCR reaction procedure followed Zhu et al.’s (2024) method [21]. The relative expression of the gene was calculated using the 2^−ΔΔCt^ method. Information on the primer sequences is shown in Table S1.
2.7. Short-Chain Fatty Acid (SCFC) Composition
The concentration of SCFCs was detected by gas chromatography–mass spectrometry (7890B GC System, Aglient, Billerica, MA, USA) with an Agilent DB-FFAP capillary column (30 m × 250 μm × 0.25 μm) to separate the samples. The initial temperature was raised form 90 °C to 160 °C at a rate of 10 °C/min. Then, 40 °C temperature was raised to 160 °C and maintained for 5 min. The carrier gas used was helium, with a flow rate of 1.0 mL/min.
2.8. 16S rDNA Sequencing
The genomic DNA of jejunal contents was extracted using the HiPure Stool DNA kits (Magen, Guangzhou, China). The 16S rRNA genes targeted the V3-V4 hypervariable region and were amplified using the 341F-806R primers (5′CCTACGGGNGGCWGCAG3′) and 806R (5-GGACTACHVGGGTWTCTAAT-3′). The paired-end sequencing was accomplished using an Illumina MiSeq PE250 platform (Illumina Inc., San Diego, CA, USA). To obtain the high-quality clean tags, raw reads were filtered using the FASTP (Version 0.18.0). The abundance statistics of taxonomy was visualized using Krona (Version 2.6). Alpha diversity analysis was calculated in QIIME (Version 1.9.1). Principal coordinate analysis (PCoA) analysis was conducted using the package “vegan” in R software (Version 4.3.1).
2.9. Metabolome Sequencing
Non-targeted metabolomics was performed using an UHPLC system (1290 Infinity LC, Agilent Technologies) coupled with a quadrupole time of flight (AB Sciex TripleTOF 6600) in Shanghai Applied Protein Technology Co., Ltd. Chromatography was carried out with an ACQUITY UPLC BEH column (2.1 × 100 mm, 1.7 μm) (Waters, Ireland). Raw MS data were converted into mzXML format using ProteoWizard MSConvert (https://proteowizard.sourceforge.io/download.html, accessed on 7 November 2023). Orthogonal partial least squares discriminant analysis (OPLS-DA) was performed by the “ropls” package in R (Version 3.3.2). The differential metabolites were defined by variable importance projection (VIP) values above 1.0 and p < 0.05. The identified metabolites were mapped to the Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway database (https://www.kegg.jp, accessed on 23 November 2023).
2.10. Statistical Analysis
The results of jejunal morphology, antioxidant capacity, immune response, digestive enzyme data, relative gene expression, and SCFAs were analyzed using a general linear model (GLM, SPSS 22.0). Results were presented as means ± SEM. A p-value of <0.05 was considered to have a significant correlation. The correlation heat maps were drawn using the R package (Version 3.3.2).
3. Results
3.1. Antioxidant Capacity, Immune Response, and Digestive Enzyme Activity of Jejunal Contents
For the antioxidant capacity (Figure 1A), the RES-HMB group showed higher concentrations of catalase (CAT, p < 0.01), glutathione peroxidase (GSH-Px, p < 0.05), and superoxide dismutase (SOD, p < 0.01) compared to the C group. For the immune response (Figure 1B), dietary RES and (or) HMB supplementation significantly improved the levels of immunoglobulin (Ig) A, IgG, IgM, and tumor necrosis factor-α (TNF-α) compared to the C group (p < 0.01). The combination of RES with HMB exhibited an optimal effect. For digestive enzyme activity (Figure 1C), the contents of α-amylase and lipase in the RES-HMB group were greater than the C and RES groups (p < 0.05 or p < 0.01).
3.2. Jejunal Morphology
H&E sections revealed the morphological alternations in the jejunal tissue of Tibetan sheep (Figure 2A). The mucosal thickness (p < 0.05) and VH/CD (p < 0.01) in the RES-HMB group was significantly increased compared with the C group. Compared to the C group, the RES group exhibited a significant increase in villi height and muscle layer thickness (p < 0.05). Additionally, the crypt depth of the C group was significantly increased compared with the HMB and RES-HMB groups (p < 0.01) (Figure 2B).
3.3. Jejunal Barrier-Related Genes Expression
The barrier-related genes expression were identified in jejunum tissues (Figure 3). The supplementation of RES and HMB alone or in combination significantly increased the mRNA expression of interleukin-10 (IL-10), mucin 2 (MUC-2), and zonula occludens-1 (ZO-1) (p < 0.01), whereas it decreased the mRNA expression of IL-1β (p < 0.01).
3.4. SCFA Concentration
As shown in Table 2, the concentrations of butyric acid were significantly increased in the RES-HMB group when compared to the C and HMB groups (p < 0.05). However, no significant difference was observed in the concentrations of other SCFAs among the four treatments (p > 0.05).
3.5. Bacterial Community Composition Analyses
A total of 530 OTUs, including common and unique, were identified as showed in Figure 4A. Of these, 332 OTUs belonged to the common type, and there were 40 unique OTUs in the C group, 61 unique OTUs in the RES group, 29 unique OTUs in the HMB group, and 68 unique OTUs in the RES-HMB group. When examining the community structure (Table 3), no significant differences in the Chao1, Ace, Shannon, and Simpson of jejunal bacteria among the four treatments were observed. The result of the PcoA analysis showed distinct differences among the treatments (Figure 4B). The Firmicutes, Proteobacteria, and Actinobacteria were the dominant phyla of the jejunal content, accounting for 82.00%, 15.72% and 1.48% (Figure 4C). At the genus level (Figure 4D), the predominantly abundant ones were Lysinibacillus (53.33%), Escherichia-Shigella (15.49%), and Bacillus (12.66%). LEfSe analysis identified eight genus and three phylum that were enriched in the four treatments (Figure 4E). The Patescibacteria, Euryarchaeota, Bacillus, Lachnospiraceae_NK3A20_group, Alistipes, Ruminococcus_gauvreauii_group, Methanobrevibacter, and Candidatus_Saccharimonas were significantly enriched in the RES-HMB group.
As seen via composition analysis (Figure 5), the relative abundance of Aeriscardovia, Firmicutes, Bacillus, Methanobrevibacter, and Actinobacteriota was significantly increased in the RES-HMB group compared with the C group. For the RES group, the relative abundance of Euryarchaeota, Firmicutes, Methanobrevibacter, Actinobacteriota, and Aeriscardovia was significantly increased compared to the C group. For the HMB group, the relative abundance of Actinobacteriota, Proteobacteria, and Escherichia-Shigella was significantly increased compared to the C group. Overall, the supplementation of RES and HMB alone or in combination significantly increased the relative abundance of Actinobacteriota.
3.6. Metabolite Profiles
The OPLS-DA analysis displayed a distinct separation between the inter-groups (Figure 6A–C). The diagrams of the permutation test of PLS-DA showed that the values of R2X, R2Y, and Q2 were 0.000, 0.860, and −0.010 (RES Vs. C, Figure 6D), 0.000, 0.840, and −0.260 (HMB Vs. C, Figure 6E), and 0.000, 0.850, and 0.070 (RES-HMB Vs. C, Figure 6F), which demonstrated that different dietary supplements in these treatments influenced the metabolic pathways of the Tibetan sheep.
According to the analysis of differential metabolites (DMs), a total of 300 DMs were identified (VIP > 1 and p < 0.05). Of these, 194 DMs (39 up-regulated and 155 down-regulated) were observed in the RES and C group (Figure 7A). Moreover, 65 DMs (20 up-regulated and 45 down-regulated) were observed in the HMB and C groups (Figure 7B). Additionally, 41 DMs (14 up-regulated and 27 down-regulated) were observed in the RES-HMB and C groups (Figure 7C).
A total of eight common DMs including Ala-Asp, His-Lys, 2′,6′-dihydroxy-4′, 4-dimethoxychalcone, 1-oleoyl-2-myristoyl-sn-glycero-3-phosphocholine, Pg 36:2, 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine, gamma-aminobutyric acid and diacetyl were observed in these four treatments.
Metabolic pathway enrichment analyses showed that the DMs in the C and RES groups were mainly enriched in the “Biosynthesis of amino acids”, “ABC transporterse”, “2-Oxocarboxylic acid”, “Butanoate metabolism”, and “Secondary bile acid biosynthesis” (Figure 7D). The DMs in the C and HMB groups were mainly enriched in “Metabolic pathways”, “Butanoate metabolism”, and “Lysine degradation” (Figure 7E). The DMs in the C and RES-HMB groups were mainly enriched in “Microbial metabolism in diverse environments”, “Butanoate metabolism”, and “Carbon metabolism” (Figure 7F).
3.7. Correlation Analysis
As showed Figure 8A, the concentration of butyric acid was positively related to the antioxidant capacity (GSH-Px, T-AOC, CAT, and SOD), immune response (IgG, IgM, and IgA), digestive enzyme activity (trypsin, α-amylase, and lipase), morphological indicators (VH/CD), and barrier-related genes expression (IL-10, MUC-2, and ZO-1), while it was negatively correlated with crypt depth and the expression of IL-1β.
Additionally, the butyric acid concentration was positively related to the abundance of Aeriscardovia, Actinobacteriota, and Methanobrevibacter, while it was negatively correlated with the contents of diacetyl (Figure 8B).
A correlation network was carried out to reflect the correlation between differential bacteria and differential metabolites (Figure 8C). The abundance of Aeriscardovia was positively correlated with 2′,6′-dihydroxy-4′,4-dimethoxychalcone and His-Lys. Furthermore, the abundance of Actinobacteriota was positively correlated with 2′,6′-dihydroxy-4′,4-dimethoxychalcone, Ala-Asp, and His-lys, while it was negatively correlated with gamma-aminobutyric acid, diacetyl, and Pg 36:2. The abundance of Escherichia-Shigella was positively correlated with gamma-aminobutyric acid, and diacetyl.
4. Discussion
It was pointed out that antioxidant enzymes constitute the first-line defense of an antioxidant defense system (CAT, GSH-Px, SOD, T-AOC) [18,22,23]. In an investigation, Ding et al. found that dietary RES increased the activities of SOD, GSH-Px, and T-AOC in the jejunum of laying hens [24]. Yang et al. also identified that dietary supplementation with 400 mg/kg RES alleviated oxidative injury by improving the activity of SOD in the duck jejunum [25]. Similarly, supplementing HMB increased the number of CD3+ and CD8+ cells and reduced the pro-inflammatory cytokine levels [26,27]. As a type of polyphenol, RES exhibited antioxidant properties similar to other polyphenols [28]. HMB is a derivative of the essential amino acid leucine metabolite a-ketoisohexanoic acid, which activates antioxidant stress via mediating ketoisohexanoic acid and isovaleryl CoA [29,30]. Our results indicated that both the RES and HMB groups significantly increased the activities of CAT and SOD in the jejunal tissues, while the RES-HMB group significantly increased the concentration of CAT, GSH-Px, SOD, and T-AOC. It is speculated that the combination of RES and HMB could enhance antioxidant enzyme activity through eliminating excess reactive oxygen (ROS) in the jejunum.
Previous research has established that RES supplementation in weaned piglets’ diets increased the concentrations of IgA and IgM in the jejunum [31]. Similarly, the levels of IgA, IgG, and IgM in piglets were increased to varying degrees with different doses of RES supplementation [32]. Moreover, a study reported RES alleviated the equine metabolic syndrome of horse via reducing the concentrations of TNF-α [33]. Evidence suggested that HMB reduced inflammatory cytokine production, thereby regulating excessive inflammation [34,35]. Using 0.2% functional amino acids (AAs) reduced weaning-induced stress and inflammation in the early stages of the growth of pig [36]. Additionally, functional AAs and polyphenols reduced dietary corticosterone-induced intestinal damage [37]. In this study, the concentrations of IgA, IgG, and IgM were significantly increased in the RES-HMB group. We speculated that the complex effect of RES and HMB regulated the immune status of the jejunum by interfering with immune cell regulation and the synthesis of pro-inflammatory cytokines.
In current study, cellulase and chymotrypsin in the contents of the jejunum were increased in the RES group. Moreover, α-amylase, lipase, and trypsin activities increased in all experimental groups, while there were more remarkable effects in the RES-HMB group. It was reported that adding 800 mg/kg RES in the diet activated lipase activity in rainbow trout [38]. Other studies have found that RES supplementation improved digestive capacity by enhancing the activity of digestive enzymes in the duodenum of Siberian sturgeon [39]. Foye et al. found that HMB significantly promoted the interaction of digestive enzymes in the jejunum of turkeys, thereby enhancing the digestion and absorption of nutrients [40]. One study found that Hawthorn, rich in acid phenols and amino acids, is endowed with the ability to promote the digestive ability by increasing the activity of digestive enzymes in the small intestine [41]. Therefore, supplementing RES and HMB simultaneously may promote the digestive ability of the jejunum in Tibetan sheep by increasing the digestive enzyme activity in the jejunal contents.
Villus height, villus width, muscle layer thickness, and crypt depth are important indexes for evaluating intestinal integrity and absorptive and digestion capacity [42,43,44]. In the current experiment, the permeability of the jejunum was improved with the RES-HMB supplement. The VH/CD of the jejunum exhibited significant increases in all treatments. The crypt depth demonstrated significant decreases in the HMB and RES-HMB groups. Apart from this, RES treatment significantly increased the muscle layer thickness and villus height of the jejunum. The HMB supplement and RES-HMB supplement significantly increased the villus width and mucosal thickness, respectively. Chen et al. confirmed that dietary RES supplementation contributed to the development of villus height in the jejunum of weaning piglets [45]. Another study reported that adding 400 mg/kg of RES improved jejunal morphology when exposing the heat stress in black bone chickens [46]. In addition, the supplementation of HMB significantly increased the value thickness of the mucosa, height, and thickness of villi in the jejunum of broiler [47]. HMB supplementation in weaned piglets increased villus high in the jejunum while decreasing crypt depth in the ileum [48].
The intestinal morphology, intestinal mucosal epithelium, and intestinal epithelium, among others, constitute the physical barrier of the gut. MUC-2 plays an important role in mucins by labeling goblet cells in the intestine [49]. As the first epithelial tight junction protein, ZO-1 is critical for efficient mucosal repair following chemical injury and mucosal healing after immune-mediated injury in intestinal epithelial cells. The abundant immune cells and related lymphoid tissue in intestinal epithelial cells jointly constitute the immune barrier of the intestine. Pro-inflammatory cytokine (IL-1β) and anti-inflammatory cytokine (IL-10) are essential cytokines for activating the immune system [50]. Prior research indicates that resveratrol treatment not only enhanced the expression of MUC-2 in mice induced by HFD [51] but also reversed intestinal damage induced by IL-1β [52]. A study on the heat-stress-induced impairment of the jejunal mucosa in black-boned chicken reported that RES increased the expression of ZO-1. RES supported the mRNA expression of IL-10 in the jejunal mucosa of weaning piglets [45]. Zhou et al. confirmed that HMB repaired deoxynivalenol-induced intestinal injury by modulating the expression of MUC-2 [53]. In young grass carp, HMB promoted anti-inflammatory cytokines and down-regulated pro-inflammatory cytokine mRNA levels to execute immunological barrier function in the small intestinal mucosa [54]. There is also a study that indicates that adding AAs and polyphenols to a diet can enhance the physical and immune barrier of the jejunum in broiler chickens [37]. In this experiment, MUC-2, ZO-1, and IL-10 were significantly increased, while IL-1β decreased by RES and HMB treatments. In general, all results represented here were consistent with those of previous studies, which suggested that the addition of RES and HMB alone or combination to Tibetan sheep diets contributed to jejunal permeability and the jejunal barrier.
As important fuels for intestinal epithelial cells (IECs), SCFAs are the main carbon flux in the diet and the main metabolite produced by the breakdown of the gut microbiota [55]. Previous research showed that RES restored intestinal bacteria to a stable level and increased the production of i-butyric acid in a murine model [56]. Baghbanzadeh-Nobari et al. found that supplementing ewe diets with 2-hydroxy-4 (methylthio) butaoic acid isopropyl ester (HMBi) increased the concentration of total VFA in the rumen [57]. These studies are consistent with our research results showing that RES-HMB treatment significantly increases the level of butyric acid in the jejunum. Although butyric acid cannot be used as a free radical scavenger, it can serve as a secondary antioxidant by affecting the DNA repair system and the levels of enzymatic antioxidants [58]. It can also induce mitochondrial autophagy by activating AMPK, thereby alleviating oxidative stress [59]. As a histone deacetylase inhibitor (HDAC), butyric acid inhibits nuclear factors-κB (NF- κB), thereby down-regulating the expression of IL-1β, IL-6, and TNF-α. Meanwhile, butyric acid acted as a ligand to activate G protein-coupled receptors GPR 41, GPR 43, and GPR 109, promoting the expression of anti-inflammatory factors [60]. In this study, butyric acid was positively related to GSH-Px, SOD, T-AOC, CAT, IgG, IgM, IgA, IL-10, MUC-2, and ZO-1. Therefore, butyric acid fermented with RES and HMB, thereby improving the antioxidant function, immune system, intestinal development, and intestinal barrier.
In this study, the relative abundance of Actinobacteriota was increased in the three treatments compared to the C group. As a key modulator in the maintenance of gut barrier homeostasis, Actinobacteria regulated the mucin biosynthesis and catabolism [61]. Besides this scenario, Actinobacteriota promoted intestinal digestion and absorption [62]. A study related to wild-type mice displayed that Actinobacteria interrelated with the SCFA in the cecum [63]. In the present study, the relative abundance of Methanobrevibacter and Bacillus was increased in the RES-HMB group. Methanobrevibacter can be translocated from the ileum and cecum to lymphatic tissues via immune cells, thereby regulating the immune system of pigs [64]. The deficiency of Methanobrevibacter disrupted the homeostasis inside the colon, leading to a decrease in mitochondrial SCFA oxidation. The gene encoding NOX was identified in Methanobrevibacter smithii to catalyze the oxidation of NADH and convert O_2_ to H_2_O, playing an important role in protecting organisms from oxidative stress and maintaining NAD+/NADH balance [65,66]. It has been observed that Bacillus has multiple effects in preventing animal intestinal infections, stimulating the immune system, reducing oxidative stress and breaking down mucins to convert them into short-chain fatty acids [67,68,69]. In our study, butyric acid was positively related to the abundance of Actinobacteriota, Methanobrevibacter, and Bacillus. Therefore, these results indicated that immune response, oxidative stress, digestive enzyme activity, intestinal barrier, and SCFA were associated with intestinal microbial composition in RES and HMB alone or in combination.
We chose pathways and important metabolites with significant differences to describe and explain their relationship with phenotypes and screened differential genera. In the present study, the concentrations of alpha-ketoglutarate and succinic semialdehyde related to butanoate metabolism are increased when fed RES and HMB. Butanoate metabolism is transported using butyric acid as a substrate, which has the same function with butyrate [70]. Alpha ketoglutarate is an essential metabolite of the mitochondrial TCA cycle, which can activate mitochondrial autophagy to inhibit ROS production [71], thereby slowing down oxidative stress and lowering the expression of IL-6 and TNF-α to inhibit chronic low-grade inflammatory response [72]. Moreover, it contributes towards reducing the activity of myosin light-chain kinase (MLCK), enhancing the concentration of the tight junction (TJ) protein, regulating endoplasmic reticulum stress, and activating Wnt/β-. The proliferation and differentiation of intestinal stem cells using catenin drugs can restore barrier function [73,74]. Butyrate mediates DNA demethylation by up-regulating acetyl CoA and alpha ketoglutarate, thereby inhibiting the colon cancer transformation index [75]. Succinic semialdehyde (SSA) is the important metabolite of γ-aminobutyric acid (GABA) [76]. GABA can suppress immune responses by reducing T cell activity and levels of inflammatory mediators, making it a classic guardian of intestinal immune homeostasis [77]. It can also inhibit the signal transduction of hydrogen peroxide (H_2_O_2_) and superoxide anions to enhance the antioxidant mechanism in cells [78]. A previous study on the Nile tilapia also reported that supplementing GABA in the diet significantly increased the levels of protease, amylase, and lipase in the intestine [79]. Diacetyl plays a critical redox role in the cycle of oxygen to produce ROS in the body [80]. Butyric acid reduced the contents of diacetyl by the inhibition of diacetyl reductase [81]. Therefore, butanoate metabolism may have a positive effect on the antioxidant capacity, immune response, digestive enzyme activity, intestinal development, and intestinal barrier of Tibetan sheep jejunum.
The Spearman correlation between microbiome and metabolome showed that diacetyl was negatively correlated with Methanobrevibacter, Actinobacteriota, and Bacillus. It is speculated that butanoate metabolism in the RES, HMB and RES-HMB trial groups was related to the changes in Methanobrevibacter, Actinobacteriota, and Bacillus.
5. Conclusions
Our results indicated that the supplementation of RES and HMB alone or in combination efficiently improved butyrate formation, thus promoting intestinal morphological development and barrier function in the jejunum of Tibetan sheep. These changes may be closely related to the microbiota, and metabolites increased the contents of butyric acid in the jejunum. The proposed mechanism is represented in Figure 9. Therefore, these findings provide insights into detailed strategies for improving jejunal health in Tibetan sheep.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Shifflett D.E. Clayburgh D.R. Koutsouris A. Turner J.R. Hecht G.A. Enteropathogenic E. coli disrupts tight junction barrier function and structure in vivo Lab. Investig.2005851308132410.1038/labinvest.370033016127426 · doi ↗ · pubmed ↗
- 2Ma Y. Yang X. Hua G. Deng X. Xia T. Li X. Feng D. Deng X. Contribution of gut microbiomes and their metabolomes to the performance of Dorper and Tan sheep Front. Microbiol.202213104774410.3389/fmicb.2022.104774436519177 PMC 9742522 · doi ↗ · pubmed ↗
- 3Bi Y. Tu Y. Zhang N. Wang S. Zhang F. Suen G. Shao D. Li S. Diao Q. Multiomics analysis reveals the presence of a microbiome in the gut of fetal lambs Gut 20217085386410.1136/gutjnl-2020-32095133589511 PMC 8040156 · doi ↗ · pubmed ↗
- 4Su S. Wang L. Fu S. Zhao J. He X. Chen Q. Belobrajdic D.P. Yu C. Liu H. Wu H. Effects of oat (Avena sativa L.) hay diet supplementation on the intestinal microbiome and metabolome of Small-tail Han sheep Front. Microbiol.202213103262210.3389/fmicb.2022.103262236590432 PMC 9801518 · doi ↗ · pubmed ↗
- 5Shoukry M.M. El-Nomeary Y. Salman F.M. Shakweer W.M.E. Improving the productive performance of growing lambs using prebiotic and probiotic as growth promoters Trop. Anim. Health Prod.20235537510.1007/s 11250-023-03752-837875638 PMC 10598149 · doi ↗ · pubmed ↗
- 6Shaito A. Posadino A.M. Younes N. Hasan H. Halabi S. Alhababi D. Al-Mohannadi A. Abdel-Rahman W.M. Eid A.H. Nasrallah G.K. Potential Adverse Effects of Resveratrol: A Literature Review Int. J. Mol. Sci.202021208410.3390/ijms 2106208432197410 PMC 7139620 · doi ↗ · pubmed ↗
- 7Corrêa M.G. Absy S. Tenenbaum H. Ribeiro F.V. Cirano F.R. Casati M.Z. Pimentel S.P. Resveratrol attenuates oxidative stress during experimental periodontitis in rats exposed to cigarette smoke inhalation J. Periodontal Res.20195422523210.1111/jre.1262230346038 · doi ↗ · pubmed ↗
- 8Farrokhi E. Ghatreh-Samani K. Salehi-Vanani N. Mahmoodi A. The effect of resveratrol on expression of matrix metalloproteinase 9 and its tissue inhibitors in vascular smooth muscle cells ARYA Atheroscler.20181415716210.22122/arya.v 14i 4.148430627191 PMC 6312567 · doi ↗ · pubmed ↗