Unlocking fertility in the female gametophyte: a DEAD-box RNA helicase is essential for embryo sac development and seed setting
Asif Ali, Asad Riaz, Xianjun Wu

TL;DR
A DEAD-box RNA helicase is crucial for rice embryo sac development and fertility.
Contribution
Identifies OsRH52A as essential for functional megaspore specification and embryo sac development in rice.
Findings
OsRH52A is required for proper embryo sac development in rice.
Loss of OsRH52A leads to defective functional megaspore specification.
OsRH52A plays a key role in seed setting and female fertility.
Abstract
This article comments on: Huang J, Qiao Z, Yu H, Lu Z, Chen W, Lu J, Wu J, Bao Y, Shahid MQ, Liu X. 2024. OsRH52A, a DEAD-box protein, regulates functional megaspore specification and is required for embryo sac development in rice. Journal of Experimental Botany 75, 4802–4821. https://doi.org/10.1093/jxb/erae180
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
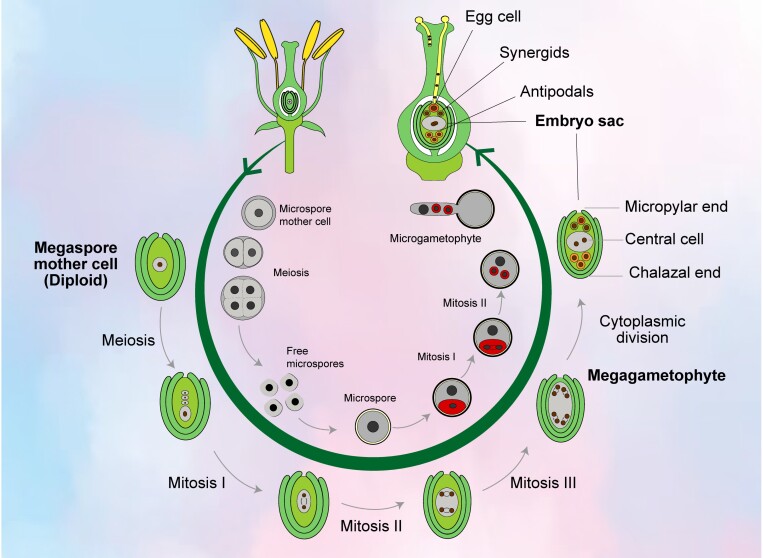 Figure 1
Figure 1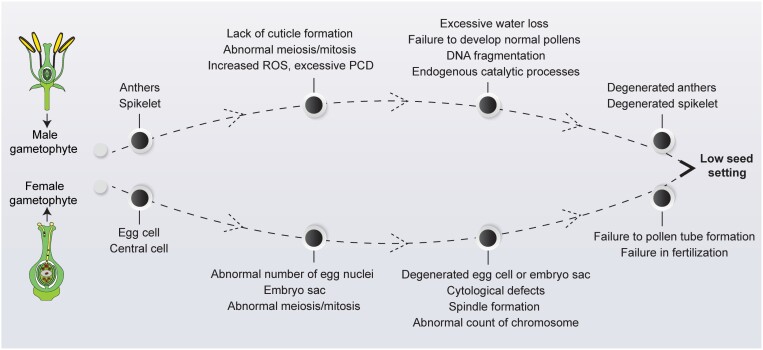 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Reproductive Biology · Plant Molecular Biology Research · Photosynthetic Processes and Mechanisms
Reproductive success and seed production rely on normal development and function of male and female organs. Previous studies reported that reproductive organs are prone to degeneration due to excessive reactive oxygen species (ROS), increased programmed cell death (PCD), developmental defects, and abnormal cell division. An embryo sac is a structure present in an ovule and is essential for developing an egg. Huang et al. (2024) revealed that loss of the DEAD-box RNA helicase OsRH52A in rice leads to abnormal development of the embryo sac including the appearance of double-female gametophyte structures and, as a consequence, to low seed setting.
A flowering plant has two phases in its life cycle, diploid sporophytic generation and haploid gametophytic generation. Generally, both male and female gametophytes are present in the same plant and are essential for sexual reproduction. A female gametophyte is required to guide the pollen tube to the ovary for fertilization and induction of seed development (Yadegari and Drews, 2004). Development of the female gametophyte is a highly organized process that occurs within an ovule and has been well reviewed in terms of several aspects, such as zygote activation (Xie et al., 2022), zygote embryogenesis (Dresselhaus and Jürgens, 2021), and embryo–endosperm interaction (Doll and Ingram, 2022). However, how defects of the female gametophyte lead to sterility and low yield remained elusive.
Embryo sac formation begins with megasporogenesis, where a megaspore mother cell (a diploid cell) undergoes meiosis, resulting in four haploid megaspores (Fig. 1). Typically, three of these megaspores degenerate, and the remaining one undergoes three rounds of mitotic division, leading to an eight-nucleate structure called the mature embryo sac. A mature embryo sac consists of three antipodal cells at the chalazal end (opposite the egg cell), two synergids and an egg cell near the micropylar end (closest to the opening for pollen tube entry), and two polar nuclei in the central cell. The fusion of the central cell (2n) with a sperm cell (n) during double fertilization forms the triploid endosperm, which provides nourishment for the developing embryo. The formation of an egg apparatus, comprising an egg cell and two synergids, is essential for fertilization. The antipodal cells, whose function is not entirely clear, are believed to play a role in nutrient transfer or hormone production. This intricate process ensures successful sexual reproduction in angiosperms by preparing the female gametophyte for fusion of the egg cell with the male gamete (sperm cell).
Male and female gametes are prone to degeneration, also known as abortion, due to several factors, for example programmed cell death (PCD), excessive reactive oxygen species (ROS), lack of proper nutrition, abnormal mitosis and meiosis, and extreme environmental conditions (Yadegari and Drews, 2004; Ali et al., 2019; Sankaranarayanan et al., 2020). Similarly, cytological aberrations of chromosomes, cytoskeleton organization, spindle assembly, and an abnormal number of sporocytes result in the failure of normal gamete development (Zhao et al., 2008; Hafidh and Honys, 2021; Shi et al., 2022; Fábián et al., 2024). In a recent study, EMBRYO SAC DEVELOPMENT 1 (ESD1) was found to encode a protein of an ovate family that regulates seed setting by controlling the development of functional megaspores (Wang et al., 2021). Similarly, DEFECTIVE EMBRYO SAC1 (DES1) that encodes a nuclear envelope membrane protein is required for embryo sac development, and its absence leads to low seed setting (Hu et al., 2023). However, detailed physiological and mechanistic insights into fertility are as yet unavailable. Seed setting is susceptible to many physiological and environmental conditions, and we outline a potential mechanism in Box 1.
OsRH52A encodes a DEAD-box helicase and is essential for maintaining embryo sac fertility
RNA helicases of the DEAD-box family are highly conserved enzymes and are reported to perform diverse biological functions including suppression of PCD by regulating the expression of downstream genes (Linder and Jankowsky, 2011). In a previous study, Yu et al. (2020) discovered that a mutation in OsRH52A is responsible for low seed setting in neo-tetraploid rice; however, its biological function and the molecular mechanism were not explored. The expression spectrum showed that OsRH52A is preferentially expressed in anther and ovary development. Huang et al. (2024) used CRISPR/Cas9 [clustered regularly interspaced palindromic repeats (CRISPR)/CRISPR-associated protein 9] to target two sites in the genome to knock out OsRH52A and developed two mutants, rh52a-m1 and rh52a-m2, in which nucleotide insertions or deletions led to a premature stop codon and truncated proteins lacking the DEXDc and HELICs domains which show significant reductions of up to ~35% and ~29%, respectively, in seed yield. Phenotypic analysis revealed that there were no apparent defects in the morphology of floral organs.
To investigate the reason for low seed setting, whole-mount eosin B-staining confocal laser scanning microscopy (WE-CLSM) was performed to search for defects associated with embryo sac fertility. In a recent study, Hu et al. (2023) also found that a mutation in DES1, which interacts with LONELY GUY (LOG), caused defective development of pistils and provided evidence that low seed setting in rh52 mutants is caused by a degenerated embryo sac. Indeed, cytological observations revealed that in contrast to the wild type (WT) with only 5% abnormal embryo sacs, rh52a-m1 and rh52a-m2 had 35% and 41% abnormal embryo sacs, respectively (Huang et al., 2024). Further analyses revealed that four different types of embryo sacs, namely the double female gametophyte (12%), degenerated embryo sacs (21%), embryo sacs without the female germline (6%), and embryo sacs without an egg and synergids (4%), were present in rh52a-m1 and rh52a-m2. The results of Huang et al. (2024) revealed the following (but not limited to) types of aberrations in embryo sacs of rh52a.
(i) Double functional megaspores: under normal (WT) conditions, usually one megasporocyte divides into two cells and then into four cells following mitosis. Among these four cells, three of them undergo degeneration and only one survives, forming only one functional megaspore (Fig. 1). However, in rh52a, two cells were present, forming a double functional megaspore.(ii) Double mono-nucleate embryo sacs: in the WT, one cell survives near the chalazal end; however, in rh52a, two cells survived, one near the chalazal end and the other near the micropylar end, and each of them developed into mono-nucleate embryo sacs forming double mono-nucleate embryo sacs.(iii) Double bi-, tetra-, and eight-nucleate embryo sacs: in WT, the functional megaspore near the chalazal end is mono-nucleate. However, in rh52a, double mono-nucleate embryo sacs undergo one, two, or three mitotic cell divisions, forming bi-, tetra-, and octa-nucleate embryo sacs, respectively.
As a result of these cytological aberrations, the fertilized embryo contained two sacs (one near the chalazal end and one near the micropylar end). Among them, the one near the chalazal end could not undergo cellularization on 1 day after fertilization (DAF) and 3 DAF. Mutants defective in female gametophyte development are helpful tools for studying the complexity of sexual reproduction, as it is affected by many factors, stages, and the position of the egg cell (Yadegari and Drews, 2004; Sun et al., 2021). Cytological aberrations observed in rh52a can be used to gain further insights into how exactly the degenerated embryo sac fails fertilization, and whether the activation, recognition, and delivery of the sperm cell were normal in rh52a. One possible reason for low fertility can be attributed to the fact that rh52a has an abnormal egg and synergids, which are essential for guiding the pollen tube. Moreover, it remains to be shown whether there are any maternal effects associated with its functional role in reproductive development and whether its overexpression can increase the seed setting. Additional studies could aim to obtain more insights into the practical implications of hybrid rice development.
Interplay of OsRH52A with fertility-associated genes OsMFS1 and ZIP4
Genetic analysis confirmed that OsRH52A is essential for functional megaspore and embryo sac development. Yeast two-hybrid (Y2H) screening was conducted to find potential interactions with other proteins involved in reproductive development. Y2H and bimolecular fluorescence complementation results revealed that OsRH52A interacts with MALE AND FEMALE STERILITY 1 (OsMFS1) and ZMM protein ZIP4. OsMFS1 encodes a coiled-coil domain-containing protein, and dysregulation of its protein causes complete sterility in male and female organs due to abnormal meiotic recombination (Lu et al., 2020). ZIP4 also plays a role in crossover formation, and its knockdown may reduce the frequency of chiasma formation that leads to sterility (Shen et al., 2012). The study of Huang et al. (2024) not only highlights the molecular pathway of embryo sac development but also advocates for more detailed investigations of meiotic recombination and synopsis for a better understanding of the ZIP4–OsRH52A–OsMFS1 complex.
OsRH52A affects the expression of downstream genes involved in megaspore development
To obtain a deeper understanding of downstream genes involved in megaspore development, a comparative transcriptome analysis of the WT and rh52a was carried out at meiosis and functional megaspore development stages. A total of 594 and 787 differentially expressed genes (DEGs) were up- and down-regulated at each developmental stage. Gene Ontology (GO) enrichment analysis indicated that the most enriched GO term was ‘microtubule-related processes’, which suggests that down-regulated DEGs were mainly responsible for defective embryo sac development. Notably, among the down-regulated DEGs, MULTIPLE SPOROCYTE1 (OsMSP1) encodes a LEUCINE-RICH PROTEIN KINASE that controls the number of sporocytes in ovule development (Zhao et al., 2008). Similarly, HYBRID STERILITY-A1 (HSA1) encodes a protein of DOMAIN OF UNKNOWN FUNCTION (DUF1618) and controls the sterility of both F_1_ and F_2_ hybrids (Kubo et al., 2016). Further studies on single and double mutants of rh52a with hsa1 and msp1 will help to understand why there is a sudden loss in the seed setting in F_2_ compared with F_1_, which remains open to study. In addition, it is thus far not known whether OsRH52A has the potential to be utilized for induction of synthetic apospory (development of an embryo from the nucellus without meiosis), which is considered an alternative mechanism to avoid heavy losses in F_2_ during hybrid seed production.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ali A , Wu T, Zhang H, et al. 2022. A putative SUBTILISIN-LIKE SERINE PROTEASE 1 (SUB Sr P 1) regulates anther cuticle biosynthesis and panicle development in rice. Journal of Advanced Research 42, 273–287.36513418 10.1016/j.jare.2022.01.003PMC 9788943 · doi ↗ · pubmed ↗
- 2Ali A , Xu P, Riaz A, Wu X. 2019. Current advances in molecular mechanisms and physiological basis of panicle degeneration in rice. International Journal of Molecular Sciences 20, 1613.30939720 10.3390/ijms 20071613 PMC 6479839 · doi ↗ · pubmed ↗
- 3Doll NM , Ingram GC. 2022. Embryo–endosperm interactions. Annual Review of Plant Biology 73, 293–321.10.1146/annurev-arplant-102820-09183835130443 · doi ↗ · pubmed ↗
- 4Dresselhaus T , Jürgens G. 2021. Comparative embryogenesis in angiosperms: activation and patterning of embryonic cell lineages. Annual Review of Plant Biology 72, 641–676.10.1146/annurev-arplant-082520-09411233606951 · doi ↗ · pubmed ↗
- 5Fábián A , Péntek BK, Soós V, Sági L. 2024. Heat stress during male meiosis impairs cytoskeletal organization, spindle assembly and tapetum degeneration in wheat. Frontiers in Plant Science 14, 1314021.38259921 10.3389/fpls.2023.1314021 PMC 10800805 · doi ↗ · pubmed ↗
- 6Hafidh S , Honys D. 2021. Reproduction multitasking: the male gametophyte. Annual Review of Plant Biology 72, 581–614.10.1146/annurev-arplant-080620-02190733900787 · doi ↗ · pubmed ↗
- 7Hu X , Yu P, Zhang Y, Gao Z, Sun B, Wu W, Deng C, Abbas A, Hong Y, Sun L. 2023. Mutation of DEFECTIVE EMBRYO SAC 1 results in a low seed-setting rate in rice by regulating embryo sac development. Journal of Experimental Botany 74, 1501–1516.36651501 10.1093/jxb/erac 506PMC 10010608 · doi ↗ · pubmed ↗
- 8Huang J , Qiao Z, Yu H, Lu Z, Chen W, Lu J, Wu J, Bao Y, Shahid MQ, Liu X. 2024. Os RH 52A, a DEAD-box protein, regulates functional megaspore specification and is required for embryo sac development in rice. Journal of Experimental Botany 75, 4802–4821.10.1093/jxb/erae 180PMC 1135008338642102 · doi ↗ · pubmed ↗