Efficacy of salmon GnRHa, Ovaprim® and hCG for hormonal stimulation of spermiation in the Fowler’s toad (Anaxyrus fowleri)
Erin M Saylor, Andrew J Kouba, Melanie R Boudreau, Nucharin Songsasen, Carrie K Kouba

TL;DR
This study compares hormones to stimulate sperm production in Fowler’s toads and finds hCG more effective than fish-derived hormones.
Contribution
Demonstrates that fish hormones like salmon GnRHa and Ovaprim® are less effective than hCG for amphibian spermiation.
Findings
hCG outperformed salmon GnRHa and Ovaprim® in sperm production and quality in Fowler’s toads.
Ovaprim® caused higher sperm abnormalities and mass gain compared to hCG.
Fish-derived hormone mixtures may not be suitable for amphibian breeding programs.
Abstract
This study showed that hCG is better for stimulating sperm production, quantity and quality than two hormones commonly used in fish, including salmon GnRH and salmon GnRH + domperidone (Ovaprim). These results suggest that there may be limited transferability of fish hormones to amphibians within the bufonid Bufonidae family. Ex situ amphibian populations can experience reproductive dysfunction due to the absence of environmental cues that trigger reproductive events. Assisted reproductive technologies (ART) for amphibians, specifically exogenous hormone regimens, can circumvent these external signals to induce gametogenesis and gamete release. Currently, the use of the mammalian reproductive hormones gonadotropin-releasing hormone (GnRH) and human chorionic gonadotropin (hCG) are used in a species-specific manner to stimulate amphibian breeding. Hormones or hormone mixtures that are…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
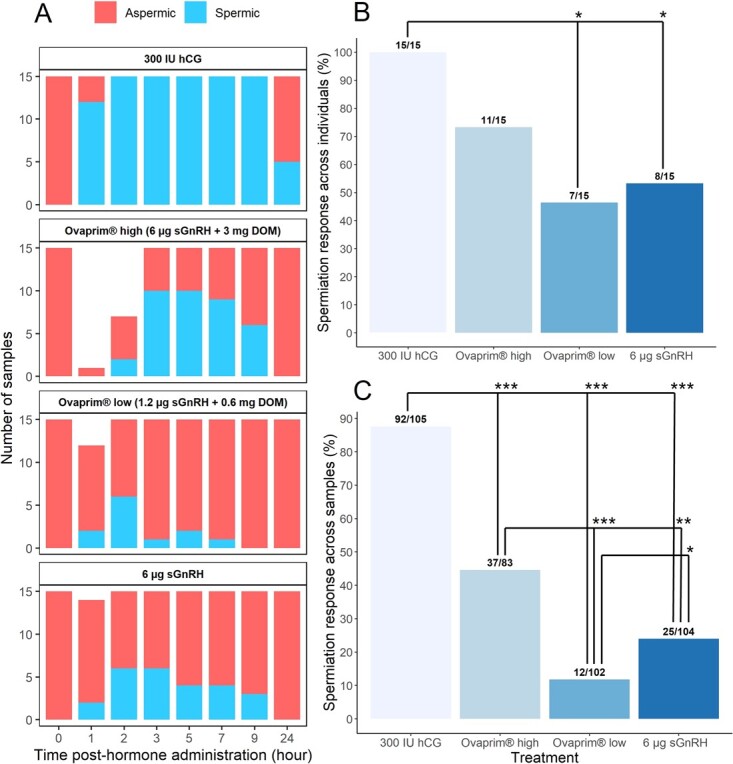 Figure 1
Figure 1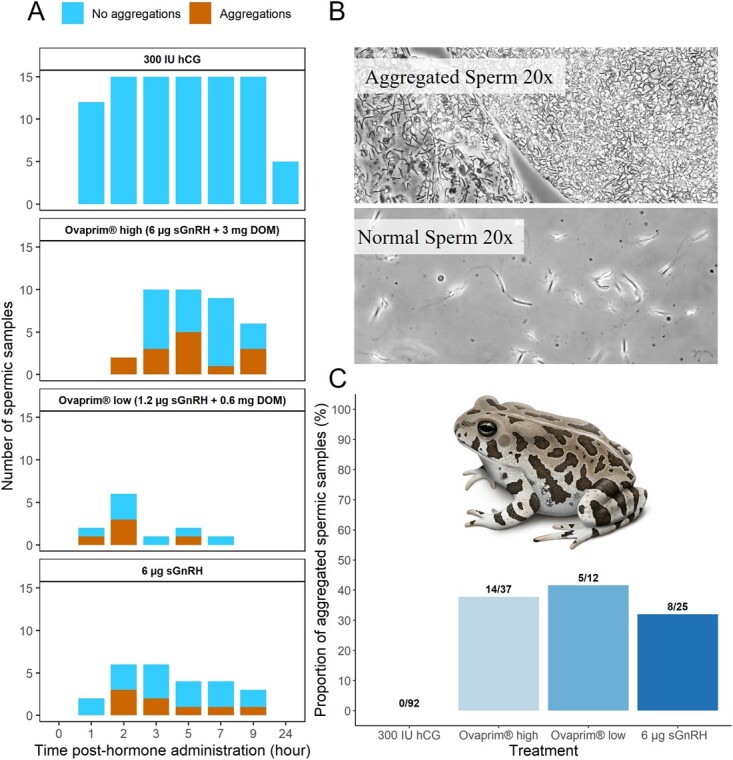 Figure 2
Figure 2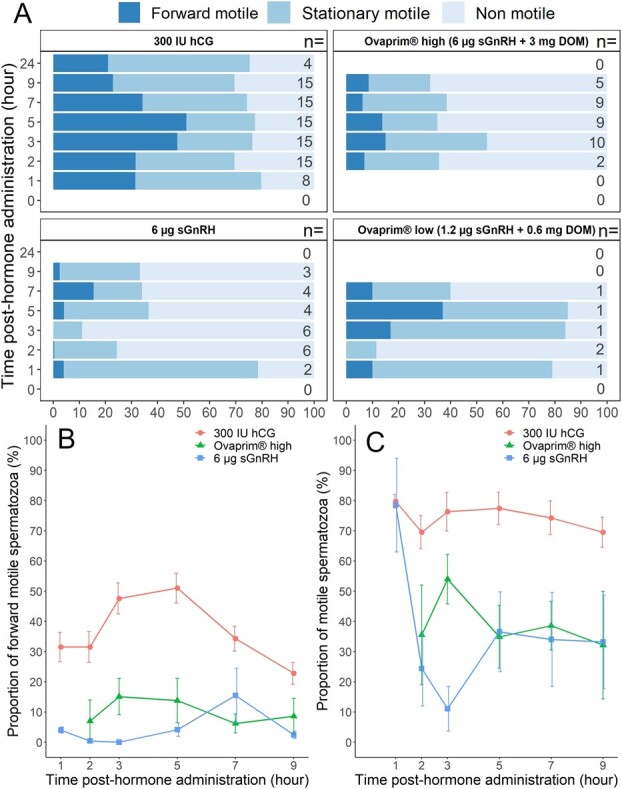 Figure 3
Figure 3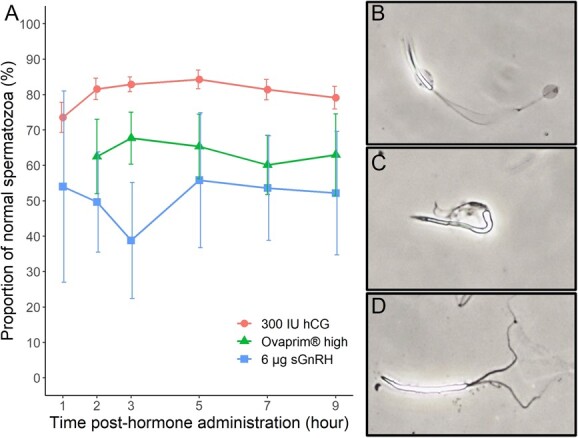 Figure 4
Figure 4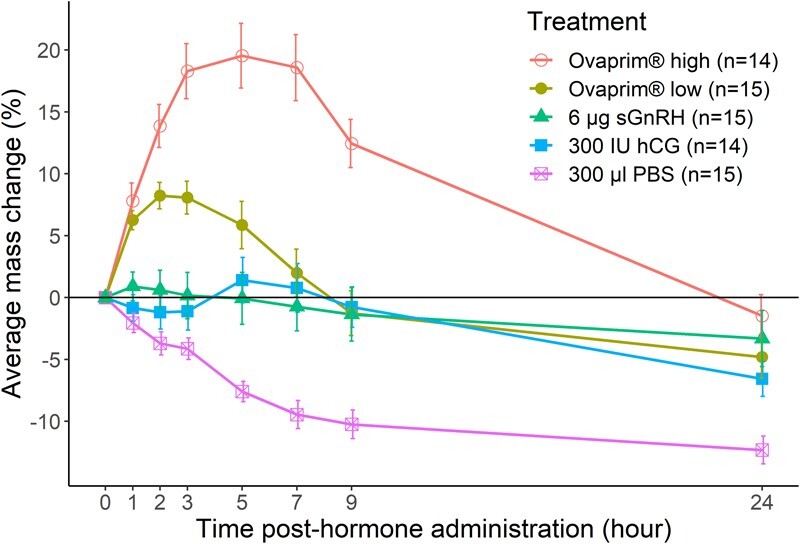 Figure 5
Figure 5|
|
|
|
|
|
|---|---|---|---|---|
|
| 300 IU | 24.0 ± 1.23 | 15/15 (100%) | 92/105 (88%) |
|
| 6 μg sGnRHa +3 mg DOM | 24.0 ± 0.80 | 11/15 (73%) | 37/83 (45%) |
|
| 1.2 μg sGnRHa +0.6 mg DOM | 24.0 ± 0.85 | 7/15 (47%) | 12/102 (12%) |
|
| 6 μg sGnRHa | 23.7 ± 0.84 | 8/15 (53%) | 25/104 (24%) |
|
| 300 μl PBS | 25.3 ± 1.01 | 0/15 (0%) | 0/105 (0%) |
| hCG | GnRH | Ovaprim® | sGnRHa | |
|---|---|---|---|---|
| Company | Sigma | Sigma | Syndel | Creative Peptides |
| Catalogue # | CG5 | L4513 | OVAM-M-ML-U010FN | OPO-011 |
| Per bottle price (USD) | $165.00 | $91.00 | $52.00 | $199.60 |
| Hormone quantity per bottle | 5000 IU | 1000 μg | 200 μg sGnRHa +100 mg DOM | 1000 μg |
| Dosage (for male | 300 IU | 0.4 μg/g BW | 6 μg sGnRHa +3 mg DOM | 6 μg |
| # | 16 | 104 | 33 | 166 |
| Cost/animal (USD) | $10.30 | $0.90 | $1.57 | $1.20 |
- —Mississippi Agricultural and Forestry Experiment Station, the National Institute of Food and Agriculture, US Department of Agriculture, Hatch project
- —US Department of Agriculture, Agricultural Research Service, Biophotonics project
- —USDA US Forest Service International Programs cooperative agreement
- —Institute of Museum and Library Services10.13039/100000208
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive biology and impacts on aquatic species · Amphibian and Reptile Biology · Animal Behavior and Reproduction
Introduction
Most temperate amphibians begin the process of reproduction in response to abiotic environmental cues such as temperature, photoperiod, barometric pressure or rainfall (Rastogi et al., 2011; Smaga et al., 2022), which are difficult to replicate in ex situ breeding programmes. Thus, captive populations of amphibians often exhibit suboptimal breeding behaviours such as calling or amplexus, poor synchronization of gamete release, low fecundity and depressed fertilization rates—all of which compromise reproductive output and the ability to maintain genetic and demographic sustainability of ex situ amphibian populations (Kouba et al., 2009, 2012, 2013; Kouba and Vance, 2009; Clulow et al., 2014; Silla and Byrne, 2019; Della Togna et al., 2020). The impact of low reproductive output and potential for inbreeding can have cascading effects on reintroduction and translocation strategies, decisions on whether to supplement captive stock with new wild founders, or even whether captive breeding programmes are a viable approach for recovery of threatened species (Silla and Kouba, 2022). Additionally, collection of gametes from both captive and wild amphibian populations is necessary for biobanking and in vitro fertilizations (IVF) for genetic management (Kouba and Julien, 2022). Because of difficulties in breeding captive amphibians and the need to obtain gametes for genetic management, the application of assisted reproductive technologies (ART) has become commonplace for many ex situ amphibian populations (Bronson and Vance, 2019; Lewis et al., 2019; Guy et al., 2020a; Bronson et al., 2021; Kouba and Julien, 2022; Burger et al., 2023). Amphibian ART utilizes exogenous hormone therapies to manipulate the hypothalamic–pituitary-gonadal axis to induce breeding or obtain gametes (Bronson and Vance, 2019; Trudeau et al., 2022). The most regularly used hormones in amphibian ART include the neuropeptide gonadotropin-releasing hormone (GnRH) and a mammalian gonadotropin, human chorionic gonadotropin (hCG), to stimulate sperm or egg release in both anurans and caudates (Kouba and Vance, 2009; Silla and Roberts, 2012; Marcec, 2016; Bronson and Vance, 2019; Graham and Kouba, 2022; Silla and Langhorne, 2022).
When produced endogenously, GnRH from the hypothalamus is released into the hypophyseal-portal system within the median eminence of the anuran brain and subsequently binds to receptors of gonadotropic cells in the pituitary to stimulate the synthesis and release of the gonadotropins luteinizing hormone (LH) and follicle stimulating hormone (FSH). Exogenous hormone therapy regimens using GnRH analogues (mainly GnRH; [des -Gly10, D-Ala6]-LH-RH) and the mammalian gonadotropin hCG have been used to induce spermiation in male anurans with varying rates of success by species (Browne et al., 2006a, 2006b; Mann et al., 2010; Trudeau et al., 2010; Kouba et al., 2012; Silla and Roberts, 2012; Langhorne et al., 2021; Burger et al., 2021; Silla and Byrne, 2021; Naranjo et al., 2022). The spermiation response to these two exogenous hormones may be family- or genera-specific. For instance, Silla and Roberts (2012) showed that five Australian anuran species belonging to the family Myobatrachidae produced more sperm in response to GnRH administration, whereas three species belonging to the family Lymnodynastidae produced more sperm in response to hCG administration. Furthermore, the American toad (Anaxyrus americanus) produced a low concentration of sperm in response to GnRH compared to hCG (Kouba et al., 2012), yet the Panamanian golden frog (Atelopus zeteki) produced more sperm in response to GnRH compared to hCG (Della Togna et al., 2017).
In addition to optimizing gamete production, hormone therapies aim to elicit appropriate breeding behaviours, which is important for natural breeding in captivity. For most male bufonid species of the genus Anaxyrus, hormone treatment using GnRH more effectively elicits natural breeding behaviours, such as calling or amplexus, compared to treatments using hCG; however, hCG treatments typically result in more animals producing higher concentrations of sperm (Kouba et al., 2009, 2012). As a result, to maximize the effects of both gamete production and breeding behaviours, the two hormones are often administered together (Calatayud et al., 2015; Graham, 2015; Guy et al., 2020a; Trudeau et al., 2022). Incorporation of dopamine antagonists with GnRH treatment has been trialled in several amphibian species as dopamine has been shown to have an inhibitory effect on gonadotropin release in amphibians (Trudeau et al., 2013; Vu and Trudeau, 2016; Della Togna et al., 2017; Bronson et al., 2021). By preventing the inhibitory action of dopamine on gonadotropin secretion the target activity on gonads may be enhanced to elicit stronger responses to hypothalamic exogenous hormone therapies (Sotowska-Brochocka and Licht, 1992; Sotowska-Brochocka et al., 1994). Although blocking the action of dopamine may enhance response to hormone therapy, dopamine plays other roles in the body such as regulation of urinary flow and water trafficking in cells (Zelenina et al., 2002; Pravikova and Ivanova, 2022). Thus, physiological responses to dopamine antagonists such as osmoregulatory changes are an important consideration when developing hormone regimens.
For the genus Anaxyrus, hCG has been established as the standard hormone used for inducing spermiation as it results in greater sperm output compared to GnRH when performing IVF procedures or when sperm is needed for biobanking genetic lineages. If the goal is to induce natural breeding, hCG is combined with GnRH because it stimulates normal reproductive behaviours and promotes spermiation leading to successful breeding events. When utilizing both hCG and GnRH in hormone therapies multiple hormones must be purchased, reconstituted, administered separately. Moreover, each hormone has strict storage requirements which can be costly and difficult for institutions to implement in breeding programmes. One potential alternative is to use commercially pre-prepared products such as Ovaprim®, which is used frequently in the aquaculture and fisheries industries for breeding brood stock.
Ovaprim®, containing salmon GnRHa (Glp-His-Trp-Ser-Tyr-DArg-Trp-Leu-Pro-NHet) plus the dopamine antagonist domperidone (DOM), was developed to enhance reproduction in captive fish brood stock. Like amphibians (Vu and Trudeau, 2016), teleost fish experience dopaminergic inhibition of reproduction (Barsagade, 2020); thus, addition of DOM to sGnRHa blocks the inhibitory action of dopamine and theoretically bolsters the response to sGnRHa. For example, male longspine scraper (Capoeta trutta) fish had an increased gonadosomatic index, higher volumes of released sperm and longer motility duration of sperm in response to Ovaprim® compared to treatments using hCG (Zadmajid, 2016). While the use of Ovaprim® in fish has been widely studied (Acharjee et al., 2017; Pawar et al., 2019; Kucharczyk et al., 2020; Yeganeh et al., 2022; Nargesi et al., 2023) its potential applicability to amphibian breeding programmes is relatively unexplored.
Despite GnRH treatment in some amphibian species resulting in poor sperm production, the GnRH analogue [des -Gly10, D-Ala6]-LH-RH has been the most widely used GnRH for sperm and egg release in anurans yet other forms of GnRH have not been thoroughly investigated. Multiple GnRH molecules and several GnRH receptor types have been found in the anuran brain (Licht and Papkoff, 1974; Muñoz-Cueto et al., 2020) at varying levels depending on the species, likely performing differing roles in regulation of reproduction. Additionally, the most abundant GnRH form found in a species is not necessarily the most potent form to endogenous receptors (Licht and Papkoff, 1974; McCreery et al., 1982; Licht et al., 1987, 1994; Millar, 2003, 2005; Muñoz-Cueto et al., 2020). Relatively low response rates and low sperm concentration in some anuran species in response to GnRH treatment may be the result of low binding affinity of GnRH to receptor types found in these species. As sGnRHa-like molecules have been found in anuran brains (Sherwood et al., 1986; King and Millar, 1995; Ebersole and Boyd, 2000), Ovaprim® or sGnRHa alone may provide more effective alternatives to GnRH when attempting hypothalamic stimulation of reproduction in amphibians. Ovaprim® may also provide a simpler, cost-efficient hormone combination that does not require reconstitution and storage of hCG and GnRH, simplifying personnel training for implementation within amphibian conservation programmes. Early application of Ovaprim® successfully induced spawning and fertilization in the tiger frog, Hoplobatrachus occipitalis (Godome et al., 2021) but failed to induce a response in the Australian southern bell frog (Litoria raniformis) (Mann et al., 2010). In neither case was the effect of sGnRHa or Ovaprim® on sperm quality or quantity assessed. Moreover, these studies suggest that the spermiation response to Ovaprim® may differ between species, as observed for other hormone treatments in anurans.
Considering the similarities in neuroendocrine control of reproduction between teleost fish and amphibians, and the differential response of anurans to Ovaprim® in previous studies, we investigated if sGnRHa and Ovaprim® administered to Fowler’s toads (Anaxyrus fowleri) would stimulate spermiation and produce sperm samples of higher quality and quantity compared to male toads treated with hCG. Additionally, we explored change in mass of treated animals in response to Ovaprim® and sGnRHa to investigate potential osmoregulatory changes due to the action of DOM in Ovaprim®. To assess the efficacy of these hormones for stimulating spermiation, Fowler’s toads were administered one of five treatments: (1) Ovaprim® high dose; (2) Ovaprim® low dose; (3) sGnRHa; (4) hCG; and (5) phosphate-buffered saline (PBS) control. We measured spermiation response rate, sperm quality and quantity parameters and recorded toad mass at eight time points over a 24-h period after hormone administration.
Materials and Methods
Collection and care of animals
Fowler’s toads (n = 30) were collected in 2021 from Oktibbeha County, Mississippi, and housed at Mississippi State University Conservation Physiology Lab Mississippi State University, MS USA (Permit #; 0504202; 0415221) in accordance with IACUC protocols (IACUC #19–345; #22–335). Animals were acclimated to captive conditions for 4–6 months prior to start of experimental trials. Toads were housed in plastic polycarbonate tanks (30 cm H × 46 cm W × 66 cm L) furnished with sheet moss (Moss Unlimited, Washington USA), refugia and a water tub. Animals were fed a rotating diet of gut-loaded crickets, mealworms and dubia roaches dusted with vitamins (Rep-Cal® Herptivite and Calcium with vitamin D_3_) three times per week. Tanks were kept under UVB lamps on a 12 h light cycle and the temperature of enclosures was ~21°C throughout the year. Animals were identified as male based on the presence of dark vocal sacs, nuptial pads and sperm production and individually identified based on their unique dorsal spot patterns.
Hormone preparation and treatment groups
All trials were performed outside of the peak breeding season (April–June) to avoid any confounding factors. Ovaprim® was obtained from Syndel company (catalogue NDC# 50378–012-01, Washington, USA) and stored at 4°C until use. Commercial Ovaprim® is marketed for intraperitoneal or intramuscular injection as a spawning aide for finfish brood stock and comes in a stock at 20 μg/ml sGnRHa +10 mg/ml DOM in a 10-ml bottle. Lyophilized sGnRHa (Glp-His-Trp-Ser-Tyr-DArg-Trp-Leu-Pro-NHet) was purchased from Creative Peptides Inc. (catalogue # OPO-011, New York, USA) and was reconstituted in PBS, aliquoted and stored at −20°C until used. Lyophilized hCG was obtained from Sigma-Aldrich (catalogue #CG5, Missouri, USA) and stored in the powder form at 4°C until needed, then reconstituted with PBS prior to administration. Males (n = 15/treatment) were randomly assigned to one of five treatment groups: (1) Ovaprim® high dose consisting of 6 μg sGnRHa + 3 mg DOM (= 300 μl Ovaprim®); (2) Ovaprim® low dose consisting of 1.2 μg sGnRHa + 0.6 mg DOM (= 60 μl Ovaprim®); (3) sGnRHa alone (6 μg sGnRHa), which is equivalent to the concentration of sGnRHa in the high-Ovaprim® treatment; (4) 300 IU hCG; and (5) a PBS control (300 μl) (as detailed in Table 1). In our design, we used a crossover experimental set-up, where a single animal can provide several experiment units. In this design, each animal is used as its own control and was randomly assigned two or more distinct hormone treatments, separated by washout periods of 2 months before another trial. As toads can be exposed to different treatments in different test periods, the experimental unit is the animal for a set period of time; this design helps to address the three Rs of experimental treatments on animals: Reduce, Refine and Reuse.
Toad mass was measured prior to hormone injection. All hormones were administered intraperitoneally using syringes with sterile 30-gauge needles. Following hormone administration, toads were placed into 19.0 × 29.5 × 15.5 cm plastic holding tubs with 1 cm of aged tap water to encourage hydration through their ventral drink patch.
Sperm collection and analysis
Urine collections occurred at 0 (prior to hormone administration), 1, 2, 3, 5, 7, 9 and 24 h post-hormone administration. To collect urine, males were quickly removed from their container, patted dry with Kimwipes® and held cloaca facing down into a sterile petri dish with their legs held apart. Toads typically urinated into dishes within 1 min of restraint, as part of their natural defence mechanism. Urine was immediately assessed for sperm presence, motility and the proportion of morphologically normal sperm determined under a phase contrast microscope (Olympus CX43; 40× magnification), and the remaining spermic urine was stored at 4°C. All toads were aspermic prior to hormone treatment (Hour 0). After each urine collection, toads were weighed to monitor any changes in water retention over time.
The presence of sperm in a urine sample indicated a spermiation response. The latency to spermiation and duration of spermiation varied considerably across individuals and across treatment groups, with some males releasing sperm at multiple hours but others only released sperm at a single hour. The spermiation response was analysed in two ways: 1) by assessing the proportion of total individuals with a spermic response at any collection point (herein called ‘spermiation across individuals’); and 2) the total number of spermic urine samples to total urine samples (herein called ‘spermiation across samples’) collected post-hormone administration (after Hour 0), within each treatment. If an individual did not provide a urine sample, the individual was excluded from the analysis for that hour. Peak response was determined by assessing the time points at which the greatest number of males responded within each treatment. Sperm motility was measured immediately after collection. Sperm were categorized as forward motile (FM) if flagellar movement was observed and the sperm progressed forward, stationary motile (SM) if there was flagellar movement but no forward progression or non-motile (NM) if no flagellar movement was observed. Sperm morphology was measured by counting the number of sperm exhibiting an abnormal head or tail (broken, bent, missing or double heads or tails). To measure sperm concentration, 10 μl of each sperm sample was diluted with hyperosmotic PBS (275–304 mOs/kg) for arresting sperm motility and counted on a Neubauer hemacytometer. Due to frequently observed aggregation of sperm in some treatments, select samples (n = 2) were stained with SYBR and propidium iodide (Burger et al., 2022) to determine the presence of live sperm in the aggregated sperm.
Panel A shows the proportion of urine samples containing spermatozoa over time for treatments: 1) 300 IU hCG; 2) 300 μl Ovaprim®; 3) 60 μl Ovaprim®; and 4) 6 μg sGnRHa. Panel B shows the spermiation response rate for the total number of individuals in each treatment, whereas Panel C shows the sperm production response rate for the total number of samples collected for each treatment ( * P < 0.05, ** P < 0.01, *** P < 0.001).
Statistical analysis
Fischer’s exact tests were used to assess overall spermiation response across individuals and samples between treatments; *P-*values with Odds Ratios (OR) are reported. Due to lack of samples collected at certain time points for some treatments, and non-normal distributions of data, a Generalized Additive Mixed Effects Model with Location Space and Scale (GAMLSS) was used to assess effects between treatments and time on both sperm motility and abnormality. A zero-inflated beta (BEZI) distribution was used with individual animal run as a random factor to eliminate the effect of individual variation in response. The changes in mass were assessed for normality by visual inspection of residual plots and a Shapiro–Wilk test and checked for heteroskedasticity using Levene’s test. Due to heterogeneity of variances and slight non-normality, a weighted least squares regression model was used to analyse the effect of treatment over time on changes in mass, with individual as a random factor. Data are presented as Means ± SEM, significance was set at P < 0.05 and statistical analyses were performed using R statistical programming software v. R 4.2.3 (R Core Team, 2023).
Results
Spermiation response across treatments
All males provided a urine sample at every hour in the hCG (Fig. 1A) and control treatments. In the Ovaprim®-high treatment, urine could not be collected from 14 and 8 males at the 1 and 2 h time point, respectively (Fig. 1A). In the Ovaprim®-low group, 3 males could not be collected at the 1-h time point (Fig. 1A). In the sGnRHa treatment, 1 male did not provide a sample at the 1 h time point (Fig. 1A). The greatest number of males producing sperm occurred at the: 1) 3 and 5 h time points (67%; n = 10/15) for the Ovaprim® high treatment; 2) 2 h time point (40%; n = 6/15) for the Ovaprim® low treatment; and 3) 2 and 3 h time points (40%; 6/15) for the sGnRHa treatment (Fig. 1A). In contrast, 80% (n = 12/15) of the hCG treated males were producing sperm at the 1 h time point and 100% of the animals were producing sperm between the 2 and 9 h time points (Table 1).
Spermiation response across individuals (i.e. total number of males producing sperm throughout trials to total number of males in treatment) with Fischer’s exact tests revealed no significant difference between hCG and the Ovaprim® high treatment (Fig. 1B). Spermiation across individuals for the Ovaprim® low and sGnRHa treatments were significantly lower compared to the hCG treatment (OR < 0.001, P < 0.001 and 0.001, respectively; Fig. 1B), but not significantly different compared to the Ovaprim® high treatment (Fig. 1B). When spermiation response was analysed as spermiation across samples (i.e. total number of spermic samples to total samples per treatment) with Fischer’s exact tests, spermiation was significantly lower in Ovaprim® and sGnRHa treatments (OR = 0.02–0.11, P < 0.05 across treatment comparisons; Fig. 1C) compared to the hCG group. In addition, spermiation across samples was significantly greater in the Ovaprim® high treatment compared to both the sGnRHa (OR = 2.40, P < 0.001) and the Low Ovaprim® treatments (OR = 5.19, P < 0.001), with the sGnRHa significantly greater (OR = 0.46, P = 0.04) than the Ovaprim® low treatment.
Sperm aggregation
Sperm aggregation occurred in both Ovaprim® treatments and the sGnRHa group (Fig. 2). The highest frequency of samples exhibiting sperm aggregation occurred in the Ovaprim® low treatment (41.6%), followed by the Ovaprim® high (37.8%) and the sGnRHa cohorts (32.0%). All sperm that were aggregated appeared non-motile. However, live/dead staining with propidium iodide and SYBR14 revealed many sperm were alive despite aggregation (data not shown). Analysis by Fisher’s exact test showed no difference (OR = 0.8–1.2, P > 0.7) in frequency of samples exhibiting sperm aggregation amongst the Ovaprim® high, Ovaprim®low and sGnRHa treatments (Fig. 2). It is also notable that aggregated sperm was not observed in any samples collected from the hCG treatment (Fig. 2).
Panel A shows the proportion of spermic samples that contained aggregated sperm over time for each treatment: 1) 300 IU hCG; 2) 300 μl Ovaprim®; 3) 60 μl Ovaprim®; and 4) 6 μg sGnRHa. Panel B shows a comparison of sperm aggregated in the sample from the Ovaprim® high treatment group to normal sperm in a sample from a male treated with hCG. Panel C shows the frequency of samples exhibiting sperm aggregation for each treatment group.
Sperm Motility, Concentration, and Morphology
Sperm aggregation in both Ovaprim® treatments and the sGnRHa group (Fig. 2) prevented accurate calculation of sperm concentration. Due to the low number of males producing sperm in the Ovaprim® low treatment (Fig. 2A), this treatment was excluded from the GAMLSS analysis of sperm motility and morphology. There was no difference in forward or total motility between the hCG and the Ovaprim® high treatments. However, compared to the hCG treatment, sGnRHa resulted in significantly lower total motility (F_29,113_ = 0.39, P < 0.001) and forward motility (F_23,119_ = 3.67, P < 0.001) across time (Fig. 3). Total motility in the sGnRHa group was also significantly lower (F_30,113_ = 20.16, P < 0.001) compared to the Ovaprim® high group over time, and there was also a marginal difference (F_23,119_ = 20.16, P = 0.08) in forward motility between these two treatments, with forward motility being slightly higher in the Ovaprim® high group (Fig. 3B and C). We also observed a significant effect (F_29,113_ = 6.04, P = 0.01) of time on total motility in the Ovaprim® high treatment, with variation in motility across time points (Fig. 3B and3C). There were fewer morphologically normal sperm in the Ovaprim® high and sGnRHa treatments compared to the hCG treatment (F_29,106_ = 5.1 and 12.32, respectively, P < 0.001 both cases; Fig. 4) over time. Although the Ovaprim® high treatment resulted in more normal sperm on average throughout time compared to sGnRHa alone (Fig. 4), this difference was not significant and there were no significant effects of time on sperm morphology in any treatment, nor was there an interaction between treatment and time.
Panel A shows the proportions of forward motile sperm (FM), stationary motile sperm (SM) and non-motile sperm (NM) in urine samples over time (0–24 h) within each treatment, with the number of samples analysed listed on the right. Panel B shows the average proportion of sperm with forward motility (FM) over time between treatments, whereas Panel C shows the average proportion of sperm exhibiting motility (FM + SM) in samples over time between treatments. All data are presented as Means ± SEM.
Panel A shows the average proportion of morphologically normal spermatozoa produced over time (0–24 h) across treatments. Data are presented as Means ± SEM. Panel B shows a morphologically normal sperm under 20× magnification. Panel C and D show an abnormal spermatozoa with misshapen head and tail or split tail, respectively, under 20× magnification.
Change in Mass
There was very little change in the mass of males treated with hCG, which remained close to 0, or baseline, throughout the collection period (Fig. 5) and is comparable to previous data (McDonough et al., 2016). However, males treated with the high dose of Ovaprim® gained 19.5% mass on average, peaking at Hour 5 and then returning to normal (0%) at 24 h post-hormone administration (Fig. 5). The change in mass for the Ovaprim® high treatment was significantly greater (F_39,544_ = 11.94–41.86, P < 0.001 across time) than change in mass for the hCG treatment across all hours, except Hour 24. Increases in mass were also observed in the Ovaprim® low treated toads (+8.2%), which peaked earlier at Hour 2 and diminished to −4.8% of their initial mass after 24 h (Fig. 5). The Ovaprim® low treatment also caused significantly greater (F_39,544_ = 0.02–9.39, P < 0.05 across time points) changes in mass compared to the hCG treatment at Hour 1, 2 and 3 (Fig. 5). Treatment with sGnRHa did not result in a significant change in mass at any time point compared to the hCG treatment (Fig. 5). Control males treated with PBS showed significant (F_39,544_ = 4.63–13.06, P < 0.05) loss of mass at Hour 5, 7, 9 and 24 compared to males treated with hCG (Fig. 5).
Average proportional mass change of individuals over time (0–24 h) across the five treatments: 1) Ovaprim® high; Ovaprim® low; 3) sGnRHa; 4) hCG; and 5) control. Data are presented as Means ± SEM.
Discussion
Effects of Ovaprim® on spermiation response and sperm quality
Understanding the effect of different hormones and dopamine antagonists for induction of spermiation and optimizing quantity and quality of sperm produced is important for the successful and efficient propagation of amphibian brood stock when using ART. Both Ovaprim® and its component sGnRHa induced spermiation yet resulted in lower spermiation rates and produced sperm with greater abnormalities and lower motility compared to hCG in the bufonid species A. fowleri. Additionally, due to many toads not providing sperm at all collection hours and latency to spermiation differing between individuals within treatments, peak spermiation response in non-hCG treatments differed widely between groups and between individuals within the same cohort.
The effects of hCG on spermiation seen in this study align with effects previously documented for certain taxa including Bufonidae and lymnodynastidae, where hCG treatments are generally more effective compared to GnRHa ([des-Gly10, D-Ala6]-LH-RH) (Kouba et al., 2012; Silla and Roberts, 2012; Uteshev et al., 2012). Other studies investigating hormonal induction of spermiation in Anaxyrus sp. (e.g. American toads; A. americanus) show that animals administered GnRHa have lower rates of spermiation and lower sperm concentration, but similar sperm motility, compared to hCG (Kouba et al., 2012). While tThe Ovaprim® high and sGnRHa treatments in this study elicited lower response rates and poorer sperm quality compared to hCG, these treatments resulted in greater spermiation response rates in A. fowleri compared to treatments of GnRHa documented in A. americanus (73.3% and 53.3%, respectively, vs. 35% with GnRHa; Kouba et al., 2012). The lower number of males responding to GnRHa in another Anaxyrus species suggests that the sGnRHa isoform may produce a greater response rate in this genus.
The differential response to GnRH isoforms and analogues may be due to the potency of the molecules varying by genera and between receptor types due to ligand selectivity. For example, in the bullfrog (Lithobates catesbeianus), chicken GnRH has a higher potency for all three GnRH receptors found in the bullfrog brain compared to sGnRHa and mammalian GnRH (Wang et al., 2001). Many species of anurans that do not respond well to either GnRHa or hCG, such as the southern bell frog (Litoria raniformis; (Mann et al., 2010), may respond better to other forms of GnRH, including sGnRHa or chicken GnRH. In addition to poor receptor–ligand specificity of some hormones, sub-optimal routes of hormone administration may play a role in poor response to hormone treatment. For example, intraperitoneal administration of GnRH elicits low spermiation rates compared to hCG in bufonids , but intranasal administration of GnRH to male A. fowleri elicits spermiation in up to 95% of males—likely due to the proximity of hormone administration to receptors in the brain (Julien et al., 2019). Assessment of spermiation response to intranasal administration of sGnRHa would provide more insight into the applicability of sGnRHa for hormonal induction of spermiation in anurans. In both A. americanus and A. fowleri, intraperitoneal administration of hCG elicits a greater response rate and higher quality sperm compared to intraperitoneal administration of sGnRHa (shown here), GnRHa (Kouba et al., 2012) and intranasal administration of GnRHa (Julien et al., 2019).
In addition to the lower spermiation response, sperm motility and poorer morphology found in our Ovaprim® and sGnRHa treatments, sperm aggregation was also frequently observed in these groups. In anurans, sperm develop in spermatogenic cysts, which travel through the testes lobule and are released by Sertoli cells through pathways dependent on NA+/K+ ion pumps (Volonteri and Ceballos, 2010). Aggregations were seen in both the Ovaprim® and sGnRHa groups but not in the hCG treatment, and aggregations have not been reported for GnRHa. In fish and mammals, GnRH peptides binding to receptors in the gonads have been implicated in paracrine signalling pathways in regulation of gonadal function (Gazourian et al., 1997; Ramakrishnappa et al., 2005), with sGnRHa causing germinal vesicle breakdown in zebrafish ovaries (Fallah and Habibi, 2020). In the amphibian, Rana esculenta, GnRH applied to testes in vitro enhanced primary spermatogonia multiplication (Minucci et al., 1986). Here, administration of exogenous sGnRHa binding to receptors on the gonads may cause a dysfunction in the release of sperm from spermatogenic cysts; however, more research is needed to confirm the exact cause of sperm aggregation.
Despite being viable, aggregated sperm produced through Ovaprim® or sGnRHa may not have optimal fertilizing capacity, as motility of sperm has been shown to be important for fertilization success (Dziminski et al., 2009). Additionally, if sperm is being collected for the purpose of cryopreservation, genetic management and IVF procedures, samples must contain an adequate concentration of spermatozoa. Because concentration of aggregated samples cannot be measured, sperm aggregations present in samples impede the use of spermic urine for both cryopreservation and IVF. Considering the lower response rate, lower sperm motilities, higher sperm abnormalities and aggregation of sperm obtained from Fowler’s toads treated with Ovaprim® compared to sperm obtained using hCG, Ovaprim® does not provide a better alternative to hCG for exogenous hormone treatment of A. fowleri to elicit spermiation.
Influence of DOM
Significantly greater spermiation response, higher sperm motility and greater % of normal sperm were obtained in the Ovaprim® high treatment compared to treatment with sGnRHa alone. These findings support existing evidence that dopamine antagonists combined with hypothalamic hormones elicits a more effective spermiation response and the release of higher quality sperm compared to treatments of hypothalamic hormones alone in anurans (Trudeau et al., 2010; Vu et al., 2017). Consistent with our results, combinations of GnRH with either DOM or a different dopamine antagonist (i.e. metoclopramide) were both effective for inducing spawning in Lithobates pipiens (Trudeau et al., 2013).
Considering Ovaprim® is designed for use in fish aquaculture, the ratio of the sGnRHa and dopamine antagonist in the mixture is specifically formulated for exogenous hormone treatment in fish. Manipulation of the ratio of sGnRHa to dopamine antagonist for amphibian hormone treatment to stimulate a hormone cascade more specific to anuran endocrine systems may increase the efficacy of spermiation treatments using sGnRHa, particularly in species that may respond well to the salmon GnRH isoform. Results from this study show that Ovaprim®, as formulated, has low transferability from fish to amphibians. In particular, the 53% response rate to sGnRHa alone warrants further investigation at higher concentrations for Anaxyrus and application to other genera that do not respond well to GnRHa. Furthermore, adjusting the concentration of dopamine antagonists when combined with sGnRHa may reduce the osmoregulatory changes observed here.
Physiological effects of hormone treatment
Difficulties in obtaining urine samples from Ovaprim® treated males within the first 2 h post-administration prevented the acquisition of spermic samples and presents a disadvantage of using Ovaprim® in A. fowleri. This observation may be related to the rapid increase in mass of the treated individuals. Additionally, mortality was seen in two males in the Ovaprim® high treatment group, and necropsy of deceased animals revealed large amounts of fluid in the coelomic cavity, indicating a strong anti-diuretic effect of Ovaprim®. Because mass gain was not seen in animals treated with sGnRHa alone, DOM may be responsible for the fluid retention. Osmoregulation in anurans is controlled by complex interactions of cellular signalling and water channel proteins embedded in cellular membranes called aquaporins (Suzuki et al., 2015). Fowler’s toads possess a drink patch located on the pelvic portion of their ventral area—an area wherein several types of aquaporins have been found in other anuran species. For example, in Xenopus laevis, an aquaporin 3 homologue exists within granular and mucous glands of skin (Mochida et al., 2008), and a homologue to mammalian aquaporin 2 is found in the ventral skin of Hyla japonica (Hasegawa et al., 2003). There are also specific aquaporins specialized for osmoregulation within the kidneys, yet other types are specialized for osmoregulation via water movement through the skin, such as aquaporin -h2 and aquaporin -h3 (Mochida et al., 2008). Thus, changes in osmoregulation seen in Fowler’s toads may occur through aquaporins in the skin or in other areas of the body such as the kidneys. Aquaporin activation pathways in anuran cells have been shown to be regulated by hormones such as arginine vasotocin, which acts as an anti-diuretic in the kidneys (Bentley, 1969) and increases water permeability in anuran skin (Saitoh et al., 2014). In previous research, use of arginine vasotocin for exogenous hormone treatment resulted in a similar retention of fluid and some mortality in A. fowleri (Julien, 2021). While the influence of dopamine on osmoregulation via aquaporins has not been researched in amphibians, in mammalian renal cells, dopamine and protein kinase C can decrease the water permeability of aquaporin 4 (Zelenina et al., 2002). Considering the possibility that dopamine may influence aquaporin activation pathways, administration of dopamine antagonists may result in changes in osmoregulation of treated animals through aquaporin regulation in the drink patch or within the kidneys. The anti-diuretic effect of the Ovaprim® high treatment further validates the recommendation that if DOM and sGnRHa were to be used together, the ratios would need to be adjusted to maintain the higher sGnRHa concentration, whilst lowering the DOM component.
Economic cost comparison of hormone treatments
To evaluate the economic viability of the Ovaprim® and sGnRHa treatments tested in this study as options for hormone therapy in anuran breeding compared to GnRHa and hCG, the cost of hormones purchased for this study are outlined in Table 2. Salmon GnRHa has the highest cost per unit, followed by hCG, Ovaprim® and GnRH. The cost was further assessed by dividing the per unit cost by the approximate number of A. fowleri that can be treated per unit to obtain a cost per animal for each hormone. hCG has the highest cost per animal (0.90). The cost per animal for Ovaprim® (1.20). Not included in this cost breakdown are the costs of reagents, laboratory equipment and personnel training required for reconstitution and storage of lyophilized hCG, GnRH and sGnRHa, which could present limitations for application in captive breeding efforts, but are otherwise similar across all these treatments.
Conclusions
The Amphibian Conservation Action Plan lists ex situ conservation initiatives such as captive breeding and biobanking as imperatives to address the amphibian extinction crisis (IUCN SSC Amphibian Specialist Group, 2005; 2022). Amphibian ex situ conservation relies on successful breeding, genetic management and transfer of genetic material between captive and wild populations, which are often difficult or impossible to achieve without the development and optimization of exogenous hormone therapies. Development of hormone regimens that successfully elicit responses in both male and female amphibians across species is an important research objective to provide a cost-effective, easy approach to amphibian assisted breeding or as a method of obtaining gametes for biobanking genetic lineages. This study is the first to find that exogenous sGnRHa alone elicits spermiation in anurans and one of the first to assess Ovaprim® as a hormone for amphibian ART in a bufonid. Although Ovaprim® or sGnRHa alone are less costly than hCG, the low sperm motility, abnormal sperm, anti-diuretic effect of DOM and aggregation of sperm present trade-offs to the lower cost per animal of Ovaprim® compared to hCG at the concentrations tested here. While sGnRHa and Ovaprim® are less costly than hCG, the latter is still the more effective hormone for sperm production, quantity and quality in bufonid species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Acharjee A , Chaube R, Joy KP (2017) Ovaprim, a commercial spawning inducer, stimulates gonadotropin subunit gene transcriptional activity: a study correlated with plasma steroid profile, ovulation and fertilization in the catfish Heteropneustes fossilis. Gen Comp Endocrinol 251: 66–73. 10.1016/j.ygcen.2016.10.001.27720753 · doi ↗ · pubmed ↗
- 2Barsagade VG (2020) Dopamine system in the fish brain: a review on current knowledge Vikas G Barsagade. J Entomol Zool Stud 8: 2549–2555. 10.22271/j.ento.2020.v 8.i 5ag.9098. · doi ↗
- 3Bentley PJ (1969) Neurohypophyseal hormones in amphibia: comparison of their actions and storage. Gen Comp Endocrinol 13: 39–44. 10.1016/0016-6480(69)90219-6.5821087 · doi ↗ · pubmed ↗
- 4Bronson E , Guy EL, Murphy KJ, Barrett K, Kouba AJ, Poole V, Kouba CK (2021) Influence of oviposition-inducing hormone on spawning and mortality in the endangered Panamanian golden frog (Atelopus zeteki). BMC Zool 6: 17. 10.1186/s 40850-021-00076-8.37170365 PMC 10127361 · doi ↗ · pubmed ↗
- 5Bronson E , Vance CK (2019) Anuran reproduction. Chapter 13 In Fowler’s Zoo and Wild Animal Medicine Current Therapy, Volume 9. Editors: RE Miller, N Lamberski and PP Calle, Elsevier Inc., St. Louis MO, USA,pp. 371–379. ISBN: 978-0-323-55228-8
- 6Browne RK , Li H, Seratt J, Kouba A (2006 a) Progesterone improves the number and quality of hormone induced Fowler toad (Bufo fowleri) oocytes. Reprod Biol Endocrinol 4: 3. 1-7. 10.1186/1477-7827-4-3.PMC 137363316451718 · doi ↗ · pubmed ↗
- 7Browne RK , Seratt J, Vance C, Kouba A (2006 b) Hormonal priming, induction of ovulation and in-vitro fertilization of the endangered Wyoming toad (Bufo baxteri). Reprod Biol Endocrinol 4.34: 1-11 10.1186/1477-7827-4-34.PMC 152477816790071 · doi ↗ · pubmed ↗
- 8Burger ID , Julien AR, Kouba AJ, Counsell KR, Barber D, Kouba(Vance) CK(2021) Linking in-situ and ex-situ populations of the endangered Puerto Rican crested toad (Peltophryne lemur) through biobanking of hormonally induced sperm. Conservation Science and Practice. e 525. 10.1111/csp 2.525 · doi ↗