Structures and genetic information of control region in mitogenomes of Odonata
Bin Jiang, Yu Yao, Jia Li, Jiang Zhang, Yang Sun, Shulin He

TL;DR
This study examines the control region of Odonata mitogenomes to reveal structural patterns that may help understand their evolutionary relationships.
Contribution
The study identifies structural differences in the control region of Odonata mitogenomes and suggests its potential for phylogenetic analysis.
Findings
Anisoptera and Anisozygoptera species show a conserved stem-loop structure near the rrns gene.
Zygoptera lacks the conserved polyC-polyG region found in other Odonata groups.
Repetition patterns in the control region are similar among sibling species like Euphaea.
Abstract
Mitogenome data of Odonata is accumulating and widely used in phylogenetic analysis. However, noncoding regions, especially control region, were usually omitted from the phylogenetic reconstruction. In an effort to uncover the phylogenetic insights offered by the control region, we have amassed 65 Odonata mitogenomes and conducted an examination of their control regions. Our analysis discovered that species belonging to Anisoptera and Anisozygoptera exhibited a stem-loop structure, which was formed by a conserved polyC-polyG stretch located near the rrns gene (encoding 12S rRNA). Conversely, the polyC-polyG region was not a conserved fragment in Zygoptera. The length and number of repetitions within the control region were identified as the primary determinants of its overall length. Further, sibling species within Odonata, particularly those in the genus Euphaea, displayed similar…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2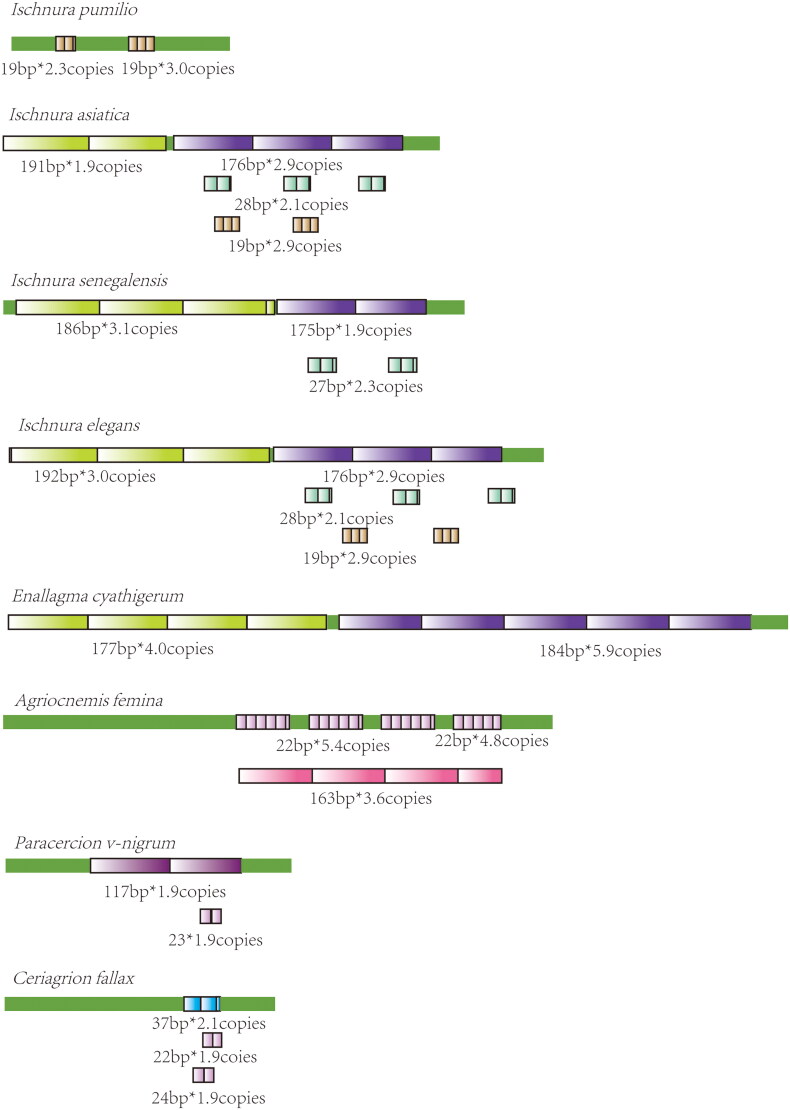 Figure 3
Figure 3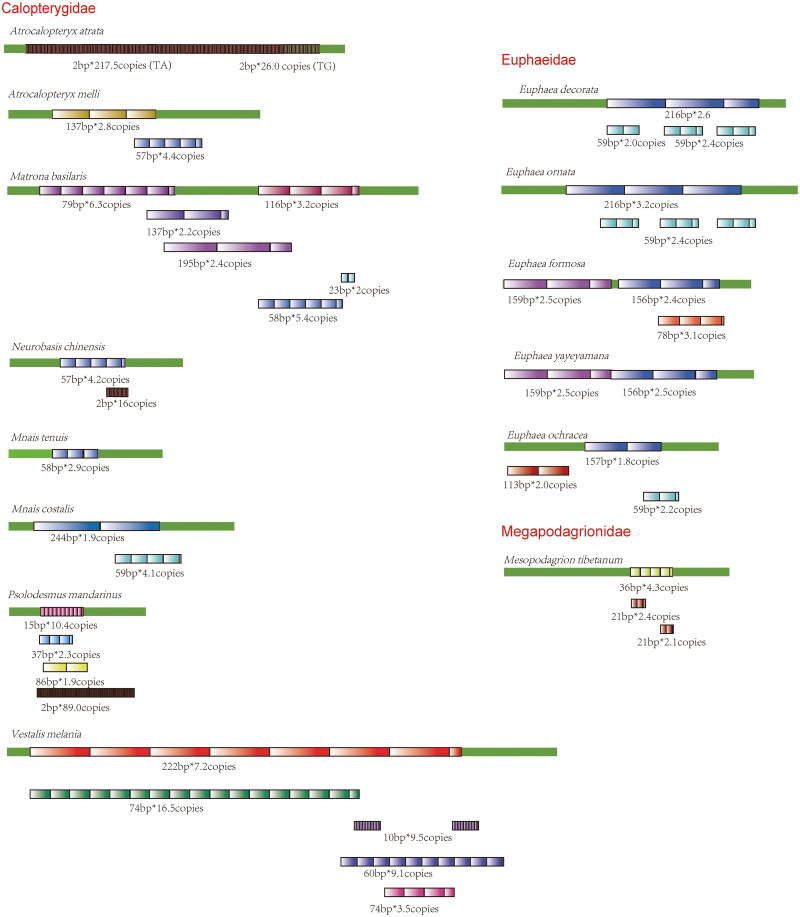 Figure 4
Figure 4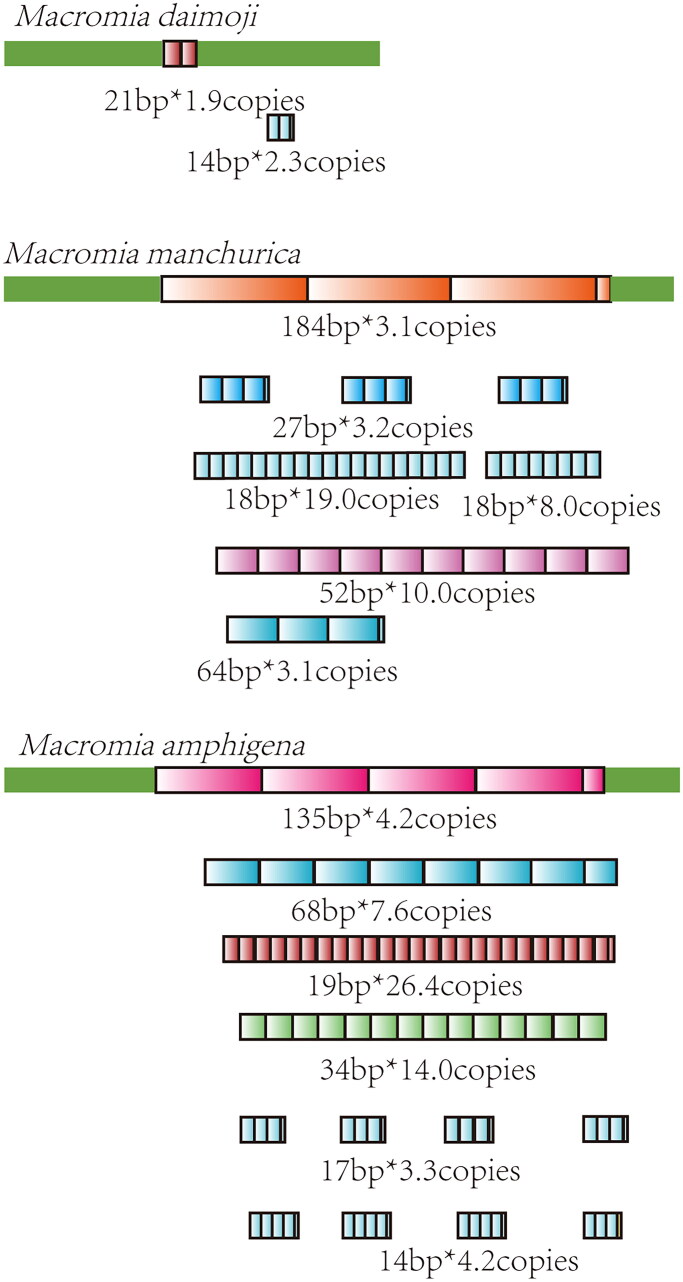 Figure 5
Figure 5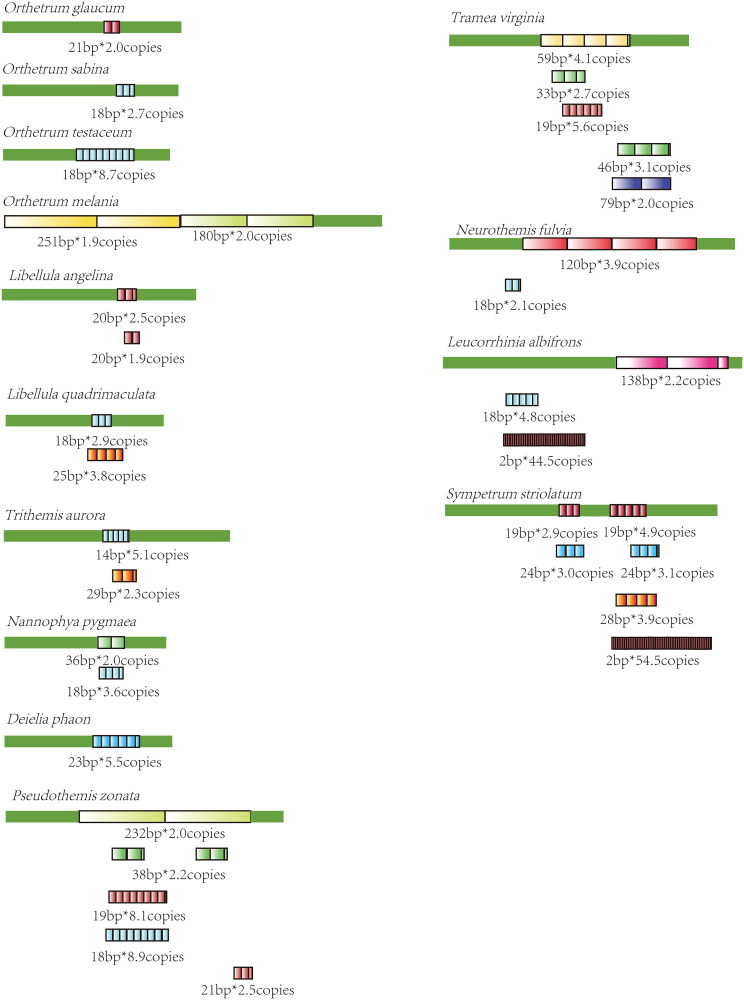 Figure 6
Figure 6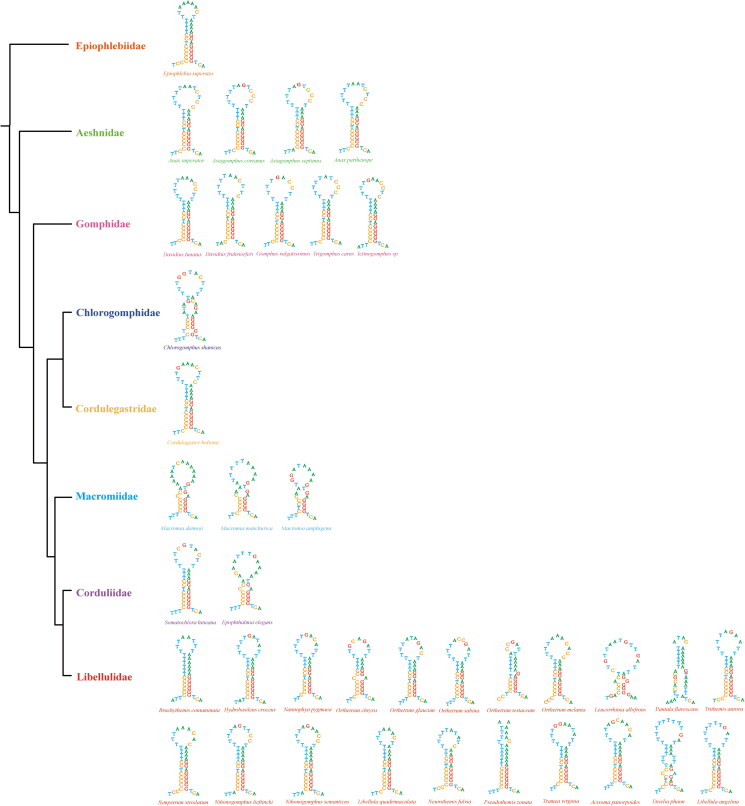 Figure 7
Figure 7| Family | Species | GenBank Acc. No. | AT (%) | References |
|---|---|---|---|---|
| Epiophlebiidae |
| 87.78 | Wang, Chen, et al. ( | |
| Aeshnidae |
| 93.53 | Feindt, Osigus, et al. ( | |
|
| 81.32 | Park et al. ( | ||
|
| 83.94 | Liao et al. ( | ||
|
| 91.78 | Ma et al. ( | ||
| Gomphidae |
| 85.18 | Lee et al. ( | |
|
| 88.24 | Liao et al. ( | ||
|
| 83.05 | Unpublished | ||
|
| 83.74 | Guan et al. ( | ||
|
| 80.68 | Tang et al. ( | ||
| Chlorogomphidae |
| 88.70 | Wang et al. ( | |
| Cordulegastridae |
| 85.55 | Unpublished | |
| Macromiidae |
| 86.38 | Kim et al. ( | |
|
| 89.19 | An et al. (2023) | ||
|
| 88.59 | An et al. ( | ||
| Corduliidae |
| 85.90 | Jackson et al. ( | |
|
| 91.91 | Wang, Wang, et al. ( | ||
| Libellulidae |
| 83.28 | Yu et al. ( | |
|
| 85.93 | Tang et al. ( | ||
|
| 84.58 | Jeong et al. ( | ||
|
| 82.93 | Yong et al. ( | ||
|
| 83.57 | Yong et al. ( | ||
|
| 83.16 | Yong et al. ( | ||
|
| 89.25 | Yong et al. ( | ||
|
| 84.30 | Guan et al. ( | ||
|
| 83.18 | Ožana et al. ( | ||
|
| 80.70 | David et al. ( | ||
|
| 84.79 | Unpublished | ||
|
| 82.09 | Feng et al. ( | ||
|
| 89.00 | Liao et al. ( | ||
|
| 87.84 | Liao et al. ( | ||
|
| 85.05 | Unpublished | ||
|
| 84.72 | Peng et al. ( | ||
|
| 88.67 | Peng et al. ( | ||
|
| 87.71 | Guan et al. ( | ||
|
| 91.78 | Guan et al. ( | ||
|
| 89.06 | Guan et al. ( | ||
|
| 82.99 | Kim et al. ( | ||
| Pseudolestidae |
| 74.20 | Unpublished | |
| Euphaeidae |
| 83.03 | Cheng et al. ( | |
|
| 81.28 | Lin et al. ( | ||
|
| 83.16 | Cheng et al. ( | ||
|
| 80.06 | Cheng et al. ( | ||
|
| 80.03 | Miga et al. ( | ||
| Megapodagrionidae |
| 79.27 | Song et al. ( | |
| Calopterygidae |
| 87.14 | Unpublished | |
|
| 77.86 | Xu et al. ( | ||
|
| 74.20 | Wang, Lin, et al. (2019) | ||
|
| 74.31 | Chen et al. ( | ||
|
| 78.03 | Song et al. ( | ||
|
| 79.11 | Okuyama and Takahashi ( | ||
|
| 80.59 | Unpublished | ||
|
| 77.45 | Lan et al. ( | ||
| Platycnemididae |
| 81.76 | Guan et al. ( | |
|
| 80.74 | Song et al. ( | ||
|
| 65.71 | Song et al. 92019) | ||
| Coenagrionidae |
| 83.78 | Feindt, Herzog, et al. ( | |
|
| 86.30 | Lorenzo-Carballa et al. ( | ||
|
| 85.44 | Jeong et al. ( | ||
|
| 81.78 | Jiang et al. ( | ||
|
| 88.40 | Jiang et al. ( | ||
|
| 88.12 | Song et al. ( | ||
|
| 85.38 | Shao et al. ( | ||
|
| 84.99 | Zhang et al. ( | ||
| Pseudostigmatidae |
| 87.65 | Feindt, Osigus, et al. ( |
- —Natural Science Foundation of Anhui Province10.13039/501100003995
- —Anhui Provincial Key Laboratory for Conservation and Utilization of Important Biological Resources
- —the Outstanding Innovative Research Team for Molecular Enzymology and Detection in Anhui Provincial Universities
- —The University Synergy Innovation Program of Anhui Province
- —Special Funds for Supporting Innovation and Entrepreneurship for Returned Oversea-students in Anhui Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhysiological and biochemical adaptations · Neurobiology and Insect Physiology Research · Genetic diversity and population structure
Introduction
Odonata is comprised three suborders, Anisoptera (dragonflies), Zygoptera (damselflies), and Anisozygoptera (including just two species of Epiophlebiidae). Odonates are exemplary of non-model organisms, extensively employed for probing pivotal issues in ecology and evolution (Bybee et al. 2016, 2021; Cordobaaguilar 2008; Jiang and Mikolajewski 2018; Stoks et al. 2003). Notably, odonates are among the most ancient of insect groups, occupying a pivotal role in the evolutionary narrative of winged insects (Lin et al. 2010). Furthermore, the diversity within the Odonata group, encompassing a multitude of species that inhabit a wide array of environmental niches, renders them invaluable as bioindicators for environmental health and the impacts of climate change (Benchalel et al. 2017; Golfieri et al. 2016; Hassall 2015; Herzog and Hadrys 2017).
To date, complete mitochondrial genomes of various animal groups have been extensively sequenced (Cameron 2014; Kenechukwu et al. 2018; Wang, Zhang, et al. 2019). Complete mitochondrial genomes provide a common and easy way for phylogeny reconstruction and population genetic analyses (Boore and Fuerstenberg 2008; Jiang et al. 2023; Saccone et al. 1999). In odonates, protein-coding genes of mitogenomes were commonly chosen for determining phylogeny reconstructions (Feindt, Osigus, et al. 2016; Yong et al. 2016). During phylogenetic analysis, the intergenic sequences were discarded in most cases, however, because of great variation, this region has gradually drawn attentions in solving phylogeny among closely related species (Barbosa et al. 2020; Bondarenko et al. 2019; Bronstein et al. 2018; Vila and Björklund 2004). Among the intergenic regions, the control region is the longest non-coding region in mitochondrial genome. In early studies, with limited number of insect species (15 species from Lepidopteran, Orthopteran, Dipteran, Hymenopteran, and Coleopteran), control regions were classified into two groups based on their structures (Zhang and Hewitt 1997). Group 1 structure contains a conserved G + A-rich sequence close to the rrns (encoding 12S rRNA) and another conserved domain, including a poly-A stretch, [TA(A)]n-like stretch and a stem-loop structure, close to trnI (tRNA^ile^). Group 2 contains no distinct conserved sequence block. With the accumulation of mitochondrial genomes, structural elements shared among certain group of species were characterized (Amaral et al. 2016; Li and Liang 2018). A prominent character of control region is the common occurrence of poly-T stretch and tandem repetition (Amaral et al. 2016), which could vary the length of control region via various copy number and different length of repeats. For example, among 12 Caliscelidae planthoppers, 1–4 repeat regions are present and the largest repeat unit is 186 bp (Gong et al. 2021). Variation in the number of repeat units could happen among species (Omote et al. 2013) and populations (Dueñas et al. 2006; Wang, Liu, et al. 2015).
In Odonata, control regions were studied sporadically. Control regions from Ephemera orientalis and Davidius lunatus exhibit different tandem repeat sequences, within which stem-loop structures may potentially form (Lee et al. 2009). Comparisons of mitogenomes from seven Zygoptera species indicates that control regions contain two conserved elements, poly-A stretches and microsatellite-like elements (TTA)3 (Juen et al. 2023). However, structural and characteristic diversity within control regions is notable and the presence of shared features across the extensive range of Odonata species is yet to be thoroughly investigated and substantiated.
Variations in the control region have been applied in evolutionary analyses, especially in intraspecific genetic variation and fast-evolving genera (Vila and Björklund 2004). The control region is widely used in studying population genetic structures (Lai et al. 2006; Robalo et al. 2021; Wang et al. 2014). Researches indicate that the combination of control region and COX1 gene is effective in elucidating the phylogeny of fast-evolving lepidopteran genera (Vila and Björklund 2004). In the damselfly genus Euphaea, the control region exhibits a relatively high substitution rate, offering a wealth of informative sites that enhance the resolution of phylogenetic analyses (Cheng et al. 2018). Utilizing the control region as a genetic marker, Parnassius glacialis has been found to possess 239 distinct haplotypes across 13 geographical populations, indicating that this region can serve as a valuable source of evidence for assessing population genetic differentiation and conducting phylogeographical studies (Wang, Pan, et al. 2019).
Conserved structural elements of control region may reflect the shared evolutionary history among species (Schultheis et al. 2002). Our study is focused on exploring the structural attributes of the control region both within and across genera of Odonata order. Thus, we have examined the structure patterns of control regions from the downloaded Odonata mitogenomes. Further, tandem repeat patterns of control regions in Zygoptera and Anisoptera were compared and analyzed.
Material and methods
Mitogenomes of Odonata (65 mitogenomes in total) were downloaded from GenBank (Table 1). Mitogenomes of Cordulia aenea, Tanypteryx hageni, and Orthetrum melania were excluded from further analyses because they are partial genomes without control regions. Control regions were initially identified based on mitogenome annotations, and the range of control regions was further confirmed based on the neighboring genes (rrns and trnI). More specifically, control regions in this study were extracted fragments from the end of rrns to the beginning of trnI in each mitogenomes.
In order to investigate conserved sequence blocks of control region in Odonata mitogenome, we aligned the control region in clustal X v2.1 with default setting (Gap opening penalty is 15 and gap extension penalty is 6.66) (Ramu et al. 2003). We explored the tandem repeats in control region by using Tandem Repeat Finder web service (Benson 1999). The distribution of AT and GC content were calculated using the online GC content calculator from VectorBuilder Tools, applying the segment window size of 70 base pairs (Lahn et al. 2023).
Results
polyC-polyG blocks in control region
3.1.
In Anisoptera and Anisozygoptera species, we found a polyC-polyG block in the control region, which mostly start around 90 bp position near rrns in the control region (28 out of 38 species in Anisoptera, Figure 1). However, we did not find similar polyC-polyG region in Pantala flavescens.
The polyC-polyG block in the control regions of Anisoptera and Anisozygoptera. the regions with brown background represent the specific polyC-polyG block. The regions with red background indicate the segments from the rrnS coding sequences near control region. Number in front of each sequence indicates the starting position for the polyC-polyG blocks. Phylogenetic relationships were modified from Bybee et al. (2021).
Analogous to the polyC-polyG blocks observed in Anisoptera, we identified a remarkable conservation of a specific fragment (characterized by the sequence GGGGTCA, as depicted in Figure 2) within the polyC-polyG blocks of 13 out of the 28 species belonging to the Zygoptera. This includes notable species such as Pseudolestes mirabilis, Euphaea decorate, E. Formosa, E. ornata, E. yayeyamana, Mesopodagrion tibetanum, Coeliccia cyanomelas, Platycnemis foliacea, Ischnura asiatica, Ischnura elegans, Ischnura senegalensis, Enallagma cyathigerum, and Agriocnemis femina. Although polyC-polyG blocks were not ubiquitous, GC-rich blocks were discovered in the remaining Zygopteran species, as revealed by the GC content distribution across the entire control region (Supplementary materials 1, Figures A1–A65). For example, a (GT)n region was discovered in Atrocalopteryx atrata (Figure 2).
Highest GC-content blocks in the control region of Zygoptera. Sequences with yellow background indicate the ‘GGGGTCA’ fragments which also occur in polyC-polyG blocks in Anisoptera. G and C were highlighted with different colors (red and blues, respectively) and underlines. Phylogenetic relationships were modified from Bybee et al. (2021).
Patterns of tandem repeats
3.2.
Nine species (Trigomphus carus, Ictinogomphus sp., Brachythemis contaminate, Hydrobasileus croceus, Orthetrum chrysis, P. flavescens, P. mirabilis, C. cyanomelas, and Platycnemis phyllopoda,) contain no repeats in their control regions. For the control regions containing tandem repeats, patterns of tandem repeats were examined.
In Coenagrionidea, I. elegans, I. asiatica, I. senegalensis, and E. cyathigerum contain two long (∼190 and ∼180 bp) repetitive elements in their control region (Figure 3, highlighted in greenish brown and purple). A short consensus sequence (ATATAAATATTTAA) in the repetitive elements was identified in P. foliacea, Ischnura species, and E. cyathigerum (Supplementary materials 2, Figure B1). In contrast, for the remaining species in Coenagrionidae (A. femina, Paracercion v-nigrum, and Ceriagrion fallax), only fragments of the consensus sequence were detected (highlighted in orange in Supplementary materials 2, Figure B1). In Calopterygidae, Matrona basilaris, Atracalopteryx melli, and Neurobasis chinensis share a 23 bp consensus sequence (ATTAAACATTTATATATATATAA, Supplementary materials 2, Figure B2); however, A. atrata features only TA or TG repeats in its control region. In Euphaeidae, E. decorata, and E. ornata display identical repeating pattern characterized by 216 bp repetitive elements (highlight in blue in Figure 4), while the other two Euphaea species only possess truncated 156–157 bp repetitive elements (highlight in blue in Figure 4). Furthermore, E. formosa and E. yayeyamana were found to share an additional 159 bp repetitive element (highlighted in purple in Figure 4).
Tandem repeats in the control region of Coenagrionidae. Repetition elements were showed with colored boxes. Length * copy numbers of each repetitive element were showed.
Tandem repeats in the control region of Calopterygidae. Repetition elements were showed with colored boxes. Length * copy numbers of each repetitive element were showed.
In Macromiidae, M. manchurica and M. amphigena were notable for their long repetitive elements (184 and 135 bp, respectively). These elements all encompass a 39 bp consensus sequence (Figure 5). Across all three Macromia species, a common sequence (ATAAATAATTTATTATAT) was discernible within the 21 bp repetitive element of M. daimoji, 52 and 64 bp repetitive elements of M. manchurica, and the 19 and 68 bp repetitive elements of M. amphigena (highlighted in red in Supplementary materials 2, Figure B3). In Libellulidae, 11 species were found to share a short sequence (ATATAAATA, highlighted with underline in Supplementary materials 2, Figure B4) in their repetition elements except two species (Trithemis aurora and Leucorrhinia albifrons) exhibiting with only fragments of the above sequence. In genus Orthetrum, O. sabina, and O. testaceum share the same 18 bp repetitive element (Figure 6). Among four Orthetrum species, a shared sequence (ATATAAATA or its complementary sequence TATATTTAT) was found in their repetitive elements (highlighted with underline or double-underline in Supplementary materials 2, Figure B4). In Gomphidae, the long repetitive elements of Davidius lunatus (261 bp2.8 copies) and Davidius fruhstorferi (260 bp2.2 copies) share high sequence similarity (Supplementary materials 2, Figure B5).
Tandem repeats in the control region of Macromiidae. Repetition elements were showed with colored boxes. Length * copy numbers of each repetitive element were showed.
Tandem repeats in the control region of Libellulidae. Repetition elements were showed with colored boxes. Length * copy numbers of each repetitive element were showed.
Discussion
Universal features in control region
4.1.
The control region exhibits significant diversity among Odonata species. The total length of control region varies not only within but also across different genera. Therefore, no general pattern of control regions was found in Odonata, which is also true in Lepidoptera (Vila and Björklund 2004) and Hemiptera (Li and Liang 2018). In Ischnura, length of control region differs between Ischnura pumilio (489 bp) and the other two Ischnura species (I. elegans: 1196 bp and I. senegalensis: 1032 bp). Length differences within genus are also found in Euphaea, Macromia, and Orthetrum. Length differences in control regions are mainly due to the various patterns of tandem repeats.
Within the control region, a polyC-polyG block was found in most of the species in our study. This block is much conserved in Anisoptera, which could form stem-loop structures (Figure 7). In mosquito and Heteroptera, a GC-rich block was also found (Duenas et al. 2006; Li and Liang 2018). A GC-rich segment was also identified as one of conserved areas in five Callitettixini species (Liu et al. 2014). Whether this kind of GC-rich blocks is widely distributed among species is still need to be investigated. GC-rich sequence element in yeast was thought to contain an origin of replication (Chen and Clark-Walker 2018). However, the role of this GC-rich block in Odonata is still unknown. However, the polyC-polyG block was not well conserved in Zygoptera. This distinctive feature between Anisoptera and Zygoptera likely reflects their divergent evolutionary histories.
Predicted stem-loop structures in the polyC-polyG blocks of Anisoptera and Anisozygoptera. Phylogenetic relationships were modified from Bybee et al. (2021).
Tandem repeats in control region
4.2.
Tandem repetitive patterns were extensively found in control regions of insect species (Zhang and Hewitt 1997). This is also true in Odonata. However, no overall repetitive pattern across Odonata species was found. Closely related species did show similar repetitive patterns. For example, in Ischnura, a highly similar repetitive pattern was found in I. asiatica, I. elegans, and I. senegalensis (all contain two long repeats), while I. pumilio did not have any of these long repetitive units. Actually, I. elegans and I. senegalensis are very closely related species on phylogeny, which was corroborated by their interspecific hybridization in the lab (Okude et al. 2020). Based on divergence age estimates, I. pumilio diverged earliest among those four Ischnura species (Blow et al. 2021). This suggested that Ischnura species have undergone length expansion in control region after separation of I. pumilio. Surprisingly, E. cyathigerum has the similar repetitive pattern as the above three Ischnura species. E. cyathigerum has overlapped distribution with I. elegans; both species contain several subspecies (Boudot and Kalkman 2015), which indicated that the radiation between these two genera was quite recent.
In Euphaea, five species showed three different repetitive patterns. One long repeat (216 bp in length) was found in E. decorata and E. ornata only; while two long repeats (159 and 156 bp in length) were found in both E. formosa and E. yayeyamana; a 157 bp repeat (similar to both E. formosa and E. yayeyamana) and another 113 bp repeat was found in E. orchracea. E. formosa, and E. yayeyamana, two island-dwelling species in east Asia, are sibling species in subtropical islands (Lee and Lin 2012b). E. decorata and E. ornata are two tropical siblings, which inhabit southeast Asian mainland and Hainan Island, respectively (Lee and Lin 2012a). Therefore, the similar pattern in control region could indicate a very close relationship on phylogeny (Jiang et al. 2021). Contrastingly, our findings revealed minimal resemblance in the tandem repeat patterns among species within the genus Orthetrum. Prior phylogenetic analyses have demonstrated that the Orthetrum species studied herein are distributed across disparate clades within phylogenetic trees (Yong et al. 2014), potentially suggesting a divergence in their evolutionary histories.
In conclusion, our findings contribute to the growing understanding of the complexity and diversity of control regions in insects, particularly in Odonata. The unique patterns and conservation levels of tandem repeats and GC-rich blocks within and across species offer valuable insights into their potential functional roles in evolutionary analysis. Future studies exploring the functional significance of these sequences and their application in phylogenetic reconstructions will further elucidate the intricate relationships within the diverse world of insects. However, the accuracy of mitogenome sequences is influenced by various methodological factors. Discrepancies are often observed in the noncoding control region due to the use of different sequencing, assembly, and annotation techniques (Velozo Timbó et al. 2017). Consequently, it is essential to prioritize the verification of mitogenome sequences in future research.
Supplementary Material
Supplimentary materials 1.pdf
Supplimentary materials 2.pdf
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1An CH, Cheon KS, Jang JE, Choi JK, Lee HG. 2022. Complete mitochondrial genome of large dragonfly (Macromia amphigena). Mitochondrial DNA B Resour. 7(2):377–378. doi:10.1080/23802359.2022.2039082.35187235 PMC 8856081 · doi ↗ · pubmed ↗
- 2An CH, Cheon KS, Jang JE, Lee HG. 2023. Complete mitochondrial genome of Macromia manchurica Asahina, 1964 (Odonata: macromiidae). Mitochondrial DNA B Resour. 8(1):10–12. doi:10.1080/23802359.2022.2157197.36620309 PMC 9815220 · doi ↗ · pubmed ↗
- 3Amaral, DT, Mitani, Y, Oliveira, G, Ohmiya, Y, Viviani, VR. 2016. Revisiting Coleoptera a + T-rich region: structural conservation, phylogenetic and phylogeographic approaches in mitochondrial control region of bioluminescent Elateridae species (Coleoptera). Mitochondrial DNA Part A, 28: 671–680.10.3109/24701394.2016.117422027159725 · doi ↗ · pubmed ↗
- 4Barbosa JTV, Barbosa MS, Morais S, Santana AEG, Almeida C. 2020. Mitochondrial genomes of genus Atta (Formicidae: myrmicinae) reveal high gene organization and giant intergenic spacers. Genet Mol Biol. 42(4):e 20180055. doi:10.1590/1678-4685-GMB-2018-0055.31188925 PMC 7197989 · doi ↗ · pubmed ↗
- 5Benchalel W, Merah S, Bouslama Z, Ramdani M, Elmsellem H, Flower R. 2017. Odonata as indicators of environmental impacts in rivers, case of wadi El-Kébir-East (northeastern Algeria). Moroccan J Chem. 5(2017):610–621.
- 6Benson G. 1999. Tandem repeats finder: a program to analyze DNA sequences. Nucleic Acids Res. 27(2):573–580. doi:10.1093/nar/27.2.573.9862982 PMC 148217 · doi ↗ · pubmed ↗
- 7Blow R, Willink B, Svensson EI. 2021. A molecularphylogeny offorktail damselflies (genus Ischnura) reveals a dynamic macroevolutionary history of female colour polymorphisms. Mol Phylogenet Evol. 160:107134. doi:10.1016/j.ympev.2021.107134.33677008 · doi ↗ · pubmed ↗
- 8Bondarenko N, Bondarenko A, Starunov V, Slyusarev G. 2019. Comparative analysis of the mitochondrial genomes of Orthonectida: insights into the evolution of an invertebrate parasite species. Mol Genet Genomics. 294(3):715–727. doi:10.1007/s 00438-019-01543-1.30848356 · doi ↗ · pubmed ↗