Non-ovarian Wolbachia pipientis titer correlates with fertility rescue of a Drosophila melanogaster bag of marbles hypomorph
Catherine H. Kagemann, Gabriela M. Colocho, Charles F. Aquadro

TL;DR
A study finds that the fertility rescue in fruit flies with a partial mutation is linked to the overall level of a bacteria called Wolbachia in the flies, not just in their ovaries.
Contribution
The study reveals that Wolbachia titer in non-ovarian tissues, not ovarian titer, correlates with fertility rescue in a Drosophila melanogaster germline stem cell mutant.
Findings
Ovarian Wolbachia titer does not differ between variants with low and high fertility rescue.
Carcass Wolbachia titer differs between variants during peak fertility rescue.
Combined ovarian and carcass Wolbachia titer positively correlates with fertility rescue.
Abstract
Bag of marbles ( bam ) is an essential gene that regulates germline stem cell maintenance and germline stem cell daughter cell differentiation in Drosophila melanogaster . When bam is partially functional (hypomorphic), the introduction of Wolbachia pipientis rescues the mutant fertility phenotype that would otherwise result in partial sterility. Infection by different W. pipientis variants results in differential rescue of the bam hypomorph fertility phenotype. We were intrigued by the varying degrees of rescue exhibited in the bam hypomorph when exposed to different W. pipientis variants, prompting us to investigate whether this phenomenon is attributable to variations in the titers of W. pipientis variants. We found no significant difference in ovarian titer between two W. pipientis variant groups, w Mel-like (low bam hypomorph fertility rescue) and w MelCS-like…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
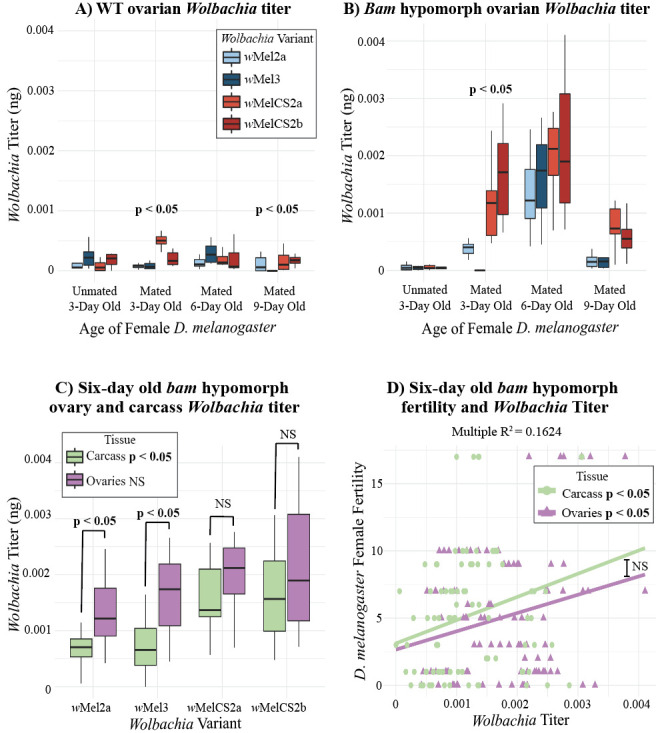 Figure 1
Figure 1- —National Institutes of Health (United States)https://ror.org/01cwqze88
- —National Institutes of Health (United States)https://ror.org/01cwqze88
- —New York State Stem Cell Science (NYSTEM)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOvarian function and disorders
Description
Bag of marbles (Bam) is a protein necessary for oogenesis and spermatogenesis in Drosophila melanogaster (McKearin and Spradling 1990) . Bam triggers germline stem cell (GSC) daughter cell differentiation during *D. melanogaster * oogenesis and regulated GSC maintenance (McKearin and Spradling 1990) . It was previously determined that the introduction of a maternally inherited bacterial endosymbiont, *Wolbachia pipientis, * rescues the mutant fertility phenotype in bam partial loss-of-function (hypomorph) mutants (Flores et al. 2015) . Without W. pipienti s, bam hypomorph flies are partially sterile (Flores et al. 2015) . The interaction between *Wolbachia pipientis * and the bam hypomorph prompted us to study the mechanisms that could play a role in the mutant fertility rescue.
W. pipientis , exhibits a diversity of variants, including well-known variants like w Mel, w Yak, and w Ri (Riegler et al. 2005) . Among these, the w Mel strain, infecting D. melanogaster , displays various variants (e.g. w Mel2a, w Mel3, w MelCS2a, and w MelCS2b) each influencing distinct host phenotypes (Bubnell et al. 2021; Christensen et al. 2019; Chrostek et al. 2013; Truitt et al. 2019) . These phenotypes encompass characteristics such as the intensity of cytoplasmic incompatibility, protection against viruses, and the host's temperature preference (Bubnell et al. 2021; Christensen et al. 2019; Chrostek et al. 2013; Truitt et al. 2019) .
There is a correlation between W. pipienti s titer and variant genotype that results in specific host phenotypes (Chrostek et al. 2013; Ilinsky and Zakharov 2011) . For example, w Mel-like variants which have a lower titer, confer lower viral protection but a longer host lifespan in males (Chrostek et al. 2013) . W MelCS-like variants cause higher viral protection and a shorter lifespan in male D. melanogaster (Chrostek et al. 2013) . Bubnell et al. (2022) determined that w MelCS-like infection in the bam hypomorph results in higher mutant fertility rescue than flies infected with w Mel-like variants. As there is evidence that *Wolbachia * genotype and titer increase specific host phenotypes, we questioned whether there is an association between W. pipientis titer and the differential bam hypomorph fertility rescue.
Kagemann et al. (2023) determined that there is a general correlation between ovarian W. pipientis titer and increases in *bam * hypomorph rescue. However, they did not address whether the differential rescue of bam hypomorph by different W. pipientis variants was driven by differences in W. pipientis variant titer or by other mechanisms specific to each variant (Kagemann et al. 2023) . While an increased *W. pipientis * titer alone may not necessarily lead to a direct increased fertility rescue in the *bam * hypomorph, an elevated titer, particularly influenced by specific strains of W. pipientis , has the potential to broaden the spectrum of host modifications. This, in turn, may enhance the effectiveness of fertility rescue in the *bam * hypomorph, especially when infected with W.
- pipientis* variants.
W. pipienti s titer has traditionally been measured using relative qPCR which utilizes the ratio of a W. pipienti s-specific gene, such as wsp or arm , relative to a host gene, such as *rpl32 * in D. melanogaster (Christensen et al. 2019; Cogni et al. 2021; Gnainsky et al. 2021; Kagemann et al. 2023) . However, relative qPCR assumes that host gene copy number is stable across conditions that are tested (Christensen et al. 2019) . Absolute quantification qPCR of W. pipienti s titer overcomes the limitations associated with relative quantification (Christensen et al. 2019) . This is achieved by comparing the copy number of W. pipienti s samples to a standard curve, allowing for the extrapolation of the total W. pipienti s copy number. As the W. pipientis infected *bam * hypomorph contains a mix of over proliferating GSC-like cells and WT GSC daughter cells, it is important to use absolute quantification to measure titer. This necessity arises from the potential variation in host gene copy numbers attributable to the diverse cell types present in the population. Our study is the first to use absolute quantification to measure ovarian and carcass W. pipientis titer of w Mel2a, w Mel3, w MelCS2a, and w MelCS2b variants infecting wildtype D. melanogaster and bam hypomorph genotypes of different ages/mating statuses.
Our results show that ovarian titer of all W. pipientis variants remained low as the female, wildtype flies aged, which contrasts with the increase that has been reported in whole male flies ( Fig. 1A ). The w MelCS-like variants infecting wildtype D. melanogaster had a higher ovarian titer than the w Mel-like variants infecting D. melanogaster in mated three-day old and nine-day old flies, but not in unmated three-day old or mated six-day old flies ( Fig. 1A linear regression model, p < 0.05).
Individual *W. pipientis * variant titer revealed that bam hypomorph flies infected with the w MelCS-like ( w MelCS2a and w MelCS2b) variants had a significantly higher titer than flies infected with the w Mel-like ( w Mel2a and w Mel3) variants in three and nine-day old flies, but not in six-day old flies ( Fig. 1B, linear regression model, p < 0.05). Kagemann et al. (2023) observed that the largest increase in *bam * hypomorph fertility rescue occurred in six-day old flies (peak fertility). However, we did not observe a statistically significant difference in ovarian titer between the W. pipientis groups ( w Mel-like and w MelCS-like) in six-day old flies suggesting another factor drives the difference in fertility rescue. We then asked whether bam hypomorph fertility rescue could be influenced by W. pipientis present in tissues outside of the ovaries during peak fertility rescue. To do this, we repeated the experiment using ovaries and carcass (whole flies excluding ovaries) from six-day old female *bam * hypomorph flies and discovered that the w MelCS-like variant carcass titer is significantly higher than the w Mel-like variant carcass titer ( Fig. 1C, linear regression model, p < 0.05). W Mel-like variant titer in the ovaries is higher than in the carcass (ANOVA, R).
We measured the fertility of the parent female flies in which ovary and carcass titer were measured in six-day old bam hypomorph flies. Due to our sample sizes (N=5 for each W. pipientis variant), we were unable to make comparisons between w Mel-like or w MelCS-like variant titer and fertility. Our correlation analyses between fertility and combined *W. pipientis * (all variants) titer show a positive correlation between both ovarian and carcass titer and fertility (Fig 1D). While the correlation between carcass titer and fertility seems higher than the correlation between ovarian titer and fertility, there is not a statistically significant difference (linear regression mode, p > 0.05, Fig 1D). With increased sample sizes, we hypothesize that w MelCS-like carcass titer would correlate with higher bam hypomorph fertility rescue compared to w Mel-like variant infected *bam * hypomorph flies.
There are at least two possible mechanisms by which W. pipientis titer outside of the ovaries could be influencing
- bam* hypomorph fertility rescue. W. pipientis have a type IV secretion system (T4SS) that allows proteins to move from cell to cell (Rancès et al. 2008). It is possible that the W. pipientis variant groups secrete different amounts of proteins, peptides, or RNA into the ovaries that could be contributing to differential rescue of the bam hypomorph by different W. pipientis variants. This is reminiscent of the impact of gut bacteria on *D. melanogaster oogenesis * (Gnainsky et al. 2021) . Secondly, the regulatory mechanisms governing W. pipientis titer in the carcass may dictate fertility rescue in the flies, drawing parallels with the interplay between Notch and Wnt signaling—crucial components in gametogenesis—and their involvement in immune cell differentiation (Rogan et al. 2019) .
Building upon existing knowledge of *W. pipientis * strain diversity and its impact on host phenotypes, our investigation, employing absolute quantification qPCR, provides novel insights into the ovarian and carcass titers of w Mel-like and w MelCS-like variants in WT and bam hypomorph D. melanogaster. While the observed differences in ovarian titer between these variants in specific age and mating conditions align with prior findings, our revelation of significantly higher carcass titers for w MelCS-like variants prompts further inquiry into their potential influence on fertility rescue. The positive correlation between combined ovarian and carcass titers and fertility suggests a broader impact beyond ovarian infection alone.
Methods
** Fly Strains and Absolute Quantification of W. pipientis Titer **
The bam ^L255F^ hypomorph mutation we used was recently remade using the same single amino acid change as the original bam ^BW^ hypomorph but in a w ^1118^ isogenic background (Bubnell et al., 2021; Bubnell et al. 2022; Flores et al. 2015) . The female bam hypomorph ( w ^1118^ ; bam ^L255F^ / bam ^[3xP3dsRed])^ that we used was generated by crossing a bam ^L255F^ /TM6 female to a bam null ( w ^1118^ ; bam ^[3xP3dsRed]/^ TM6) male (McKearin and Spradling 1990; Bubnell et al 2021) . An uninfected bam hypomorph control was used along with WT bam w ^1118^ fly lines containing each
- W. pipientis * variant. The four *W. pipientis * variants and uninfected control were generously provided by Luis Teixeira and are described in Chrostek et al. (2013). All D. melanogaster lines were maintained on yeast glucose food and placed in an incubator at 25°C with a 12-hour light-dark cycle.
W. pipientis titer was first measured in WT and bam hypomorph ovaries in unmated three-day old, mated three-day old, mated six-day old, and mated nine-day old flies and is described in Kagemann et al. (2023). Five biological replicates were used per W. pipientis variant infecting the WT or bam hypomorph along with three technical replicates per biological replicate. We did not make a direct comparison of ovarian titers between the Bam hypomorph and WT because their DNA extractions were performed on different 96-well plates and diluted separately.
Subsequently, the experiment was repeated in six-day old bam hypomorph flies to measure both ovary and carcass W. pipientis titer. Flies were dissected from flash frozen female parents from our fertility assays using absolute quantification qPCR. Methods for absolute quantification of *W. pipientis * in the ovaries and carcass (whole flies without ovaries) are described in Kagemann et al. (2023). QPCR was run on a QuantStudio 7 Pro provided by Cornell’s Biotechnology Resource Center.
** Six-Day Old *Bam * Hypomorph Fertility Assay **
Our experimental design allowed us to quantify fertility and titer from the same bam hypomorph female flies (Fig 1D). We aged female *bam * hypomorph flies infected with each W. pipientis variant for five days and mated them with two three-day old Canton-S males for 24 hours in vials containing yeast glucose food. Six-day old *bam * hypomorph females were subsequently collected for qPCR while the vials were kept at 25°C with a 12-hour light-dark cycle. The progeny from the vials were counted every two days for eight days in total and the sum of the progeny were used for our analyses.
Statistical Analyses
To assess correlations between D. melanogaster fertility found in Kagemann et al. (2023) and *W. pipientis * variant titer, we used a linear regression model. A linear regression model was conducted in R (v. 4.1.0) at each age/mating status to determine statistically significant differences in W. pipientis titer between D. melanogaster infected with different W. pipientis groups ( w Mel-like and w MelCS-like) and between the uninfected control (Fig 1A and 1B). The equation used was as follows: W. pipientis titer (response variable) ∼ W. pipientis Group * Day (Fig 1A and 1B). A two-way ANOVA was used to find differences in W. pipientis variant ovary and carcass titer (Fig 1C). To find correlations between D. melanogaster fertility and W. pipientis group titer in the ovaries and carcass we used a linear regression model (Fig 1D). The equation used was as follows: D. melanogaster Fertility ~ W. pipientis Group Titer (Fig 1D).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bubnell Jaclyn E Fernandez-Begne Paula Ulbing Cynthia K S Aquadro Charles F 202197 Diverse w Mel variants of Wolbachia pipientis differentially rescue fertility and cytological defects of the bag of marbles partial loss of function mutation in Drosophila melanogaster G 3 Genes|Genomes|Genetics 11122160-183610.1093/g 3journal/jkab 312PMC 866447134580706 · doi ↗ · pubmed ↗
- 2Christensen Steen Camacho Moises Sharmin Zinat Momtaz A. J. M. Zehadee Perez Laura Navarro Giselle Triana Jairo Samarah Hani Turelli Michael Serbus Laura R. 201993 Quantitative methods for assessing local and bodywide contributions to Wolbachia titer in maternal germline cells of Drosophila BMC Microbiology 1911471-218010.1186/s 12866-019-1579-3PMC 672436731481018 · doi ↗ · pubmed ↗
- 3Chrostek Ewa Marialva Marta S. P. Esteves Sara S. Weinert Lucy A. Martinez Julien Jiggins Francis M. Teixeira Luis 20131212 Wolbachia Variants Induce Differential Protection to Viruses in Drosophila melanogaster: A Phenotypic and Phylogenomic Analysis P Lo S Genetics 9121553-7404 e 1003896 e 100389610.1371/journal.pgen.100389624348259 PMC 3861217 · doi ↗ · pubmed ↗
- 4Cogni Rodrigo Ding Shuai Dominique Pimentel André C. Day Jonathan P. Jiggins Francis M. 20211125 Wolbachia reduces virus infection in a natural population of Drosophila Communications Biology 412399-364210.1038/s 42003-021-02838-z PMC 861717934824354 · doi ↗ · pubmed ↗
- 5Flores Heather A. Bubnell Jaclyn E. Aquadro Charles F. Barbash Daniel A. 2015820 The Drosophila bag of marbles Gene Interacts Genetically with Wolbachia and Shows Female-Specific Effects of Divergence PLOS Genetics 1181553-7404 e 1005453 e 100545310.1371/journal.pgen.100545326291077 PMC 4546362 · doi ↗ · pubmed ↗
- 6Gnainsky Yulia Zfanya Nofar Elgart Michael Omri Eman Brandis Alexander Mehlman Tevie Itkin Maxim Malitsky Sergey Adamski Jerzy Soen Yoav 202111 Systemic Regulation of Host Energy and Oogenesis by Microbiome-Derived Mitochondrial Coenzymes Cell Reports 3412211-124710858310858310.1016/j.celrep.2020.10858333406416 · doi ↗ · pubmed ↗
- 7Grobler Yolande Yun Chi Y. Kahler David J. Bergman Casey M. Lee Hangnoh Oliver Brian Lehmann Ruth 20181113 Whole genome screen reveals a novel relationship between Wolbachia levels and Drosophila host translation PLOS Pathogens 14111553-7374 e 1007445 e 100744510.1371/journal.ppat.100744530422992 PMC 6258568 · doi ↗ · pubmed ↗
- 8Ilinsky Yu. Yu. Zakharov I. K. 201191 Cytoplasmic incompatibility in Drosophila melanogaster is caused by different Wolbachia genotypes Russian Journal of Genetics: Applied Research 152079-059745846210.1134/s 2079059711020031 · doi ↗