Microglial process convergence onto injured axonal swellings, a human postmortem brain tissue study
Amanda L. Logan-Wesley, Karen M. Gorse, Audrey D. Lafrenaye

TL;DR
This study investigates how microglia interact with injured axons in human brains after traumatic brain injury, confirming a previously observed phenomenon in animals.
Contribution
The study is the first to quantitatively measure microglial process convergence onto injured axons in human postmortem brain tissue.
Findings
Microglial process convergence onto injured axonal swellings was increased in TBI cases.
Findings are consistent with previous observations in micropigs, suggesting clinical relevance.
MPC was not observed on intact myelinated fibers, highlighting specificity to injury.
Abstract
Traumatic brain injury (TBI) affects millions globally, with a majority of TBI cases being classified as mild, in which diffuse pathologies prevail. Two of the pathological hallmarks of TBI are diffuse axonal injury and microglial activation. While progress has been made investigating the breadth of TBI-induced axonal injury and microglial changes in rodents, the neuroinflammatory progression and interaction between microglia and injured axons following brain injury in humans is less well understood. Our group previously investigated microglial process convergence (MPC), in which processes of non-phagocytic microglia directly contact injured proximal axonal segments, in rats and micropigs acutely following TBI. These studies demonstrated that MPC occurred on injured axons in the micropig, but not in the rat, following diffuse TBI. While it has been shown that microglia co-exist and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
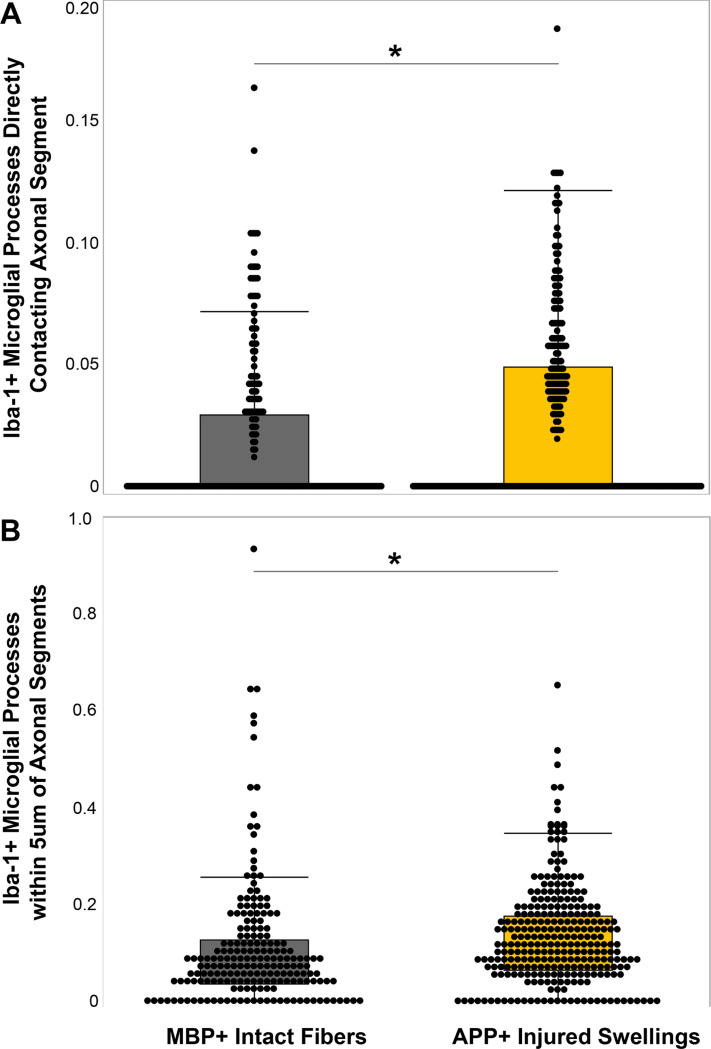 Figure 1
Figure 1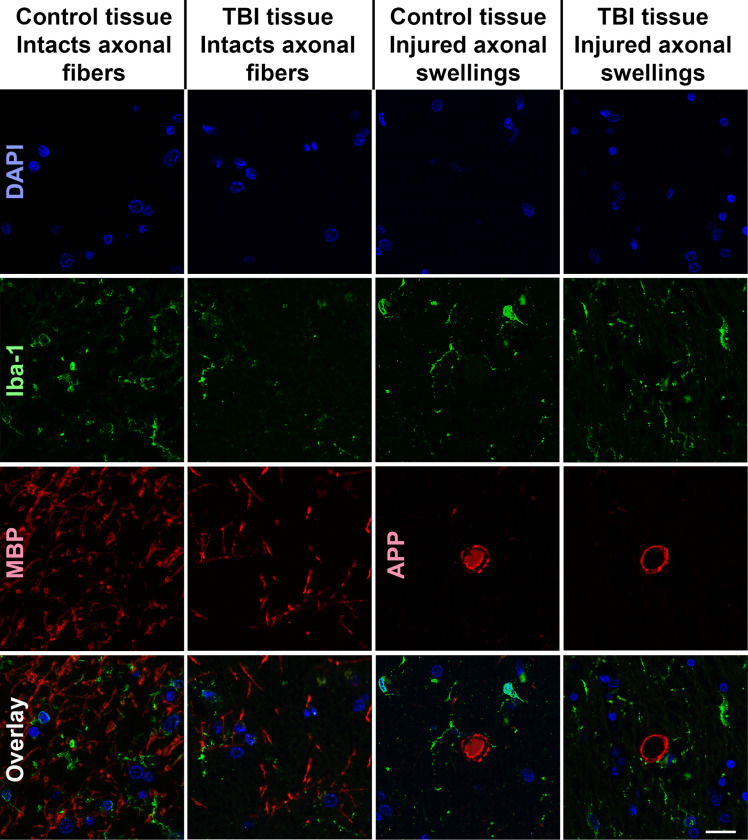 Figure 2
Figure 2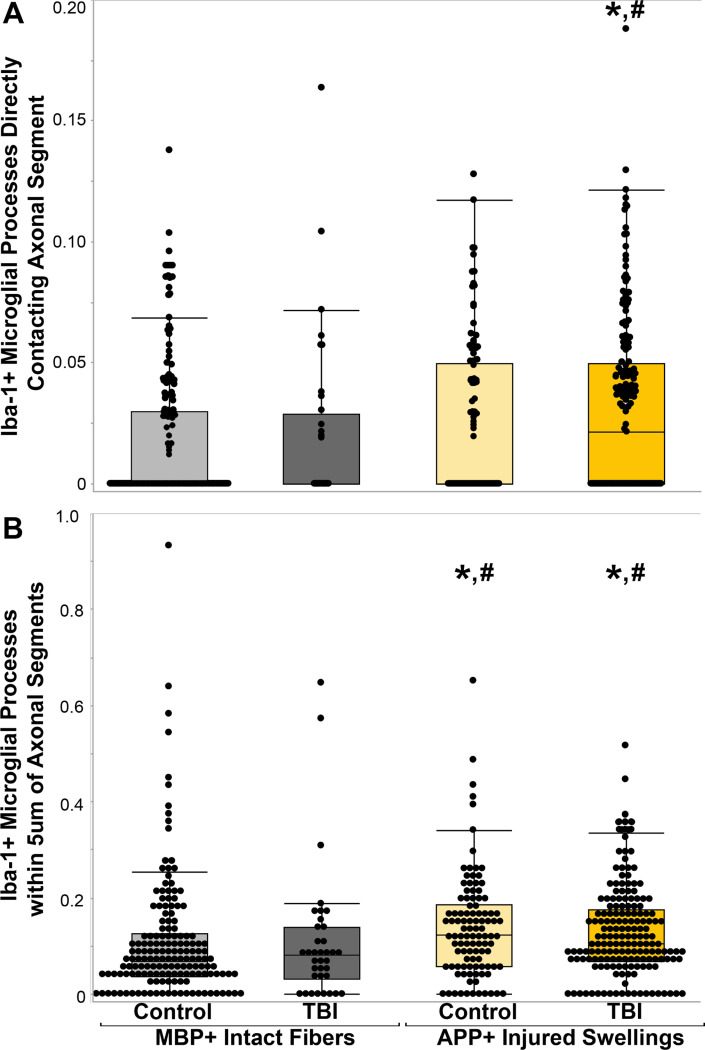 Figure 3
Figure 3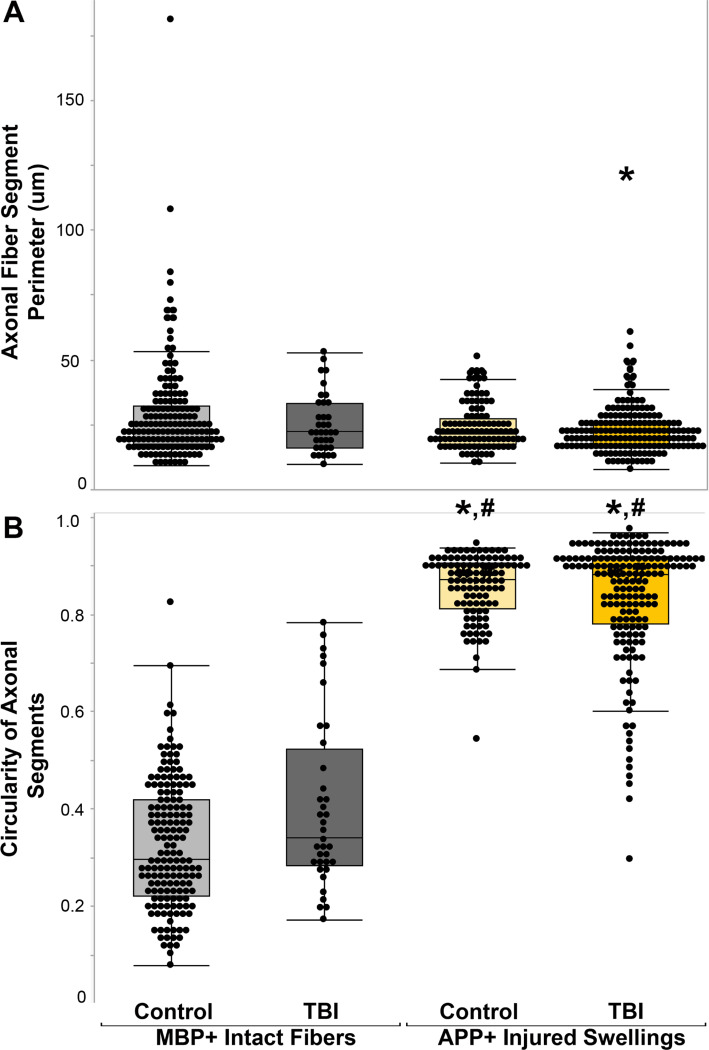 Figure 4
Figure 4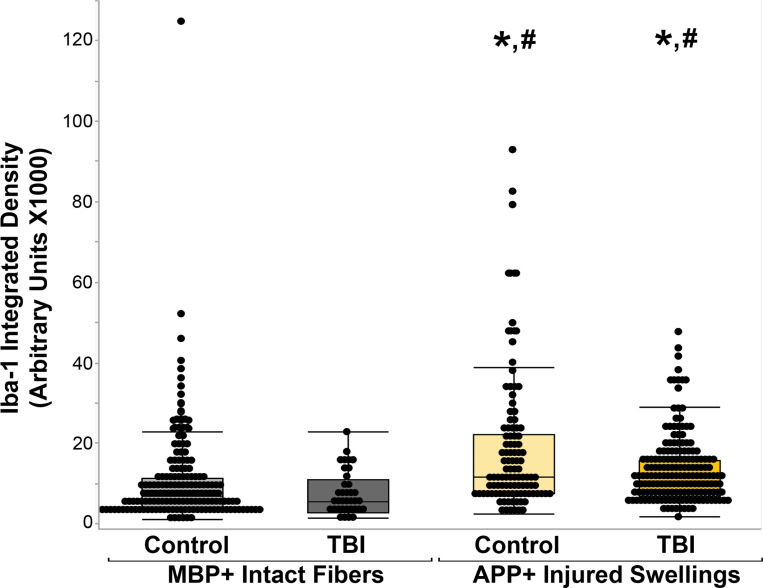 Figure 5
Figure 5- —NINDS
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTraumatic Brain Injury and Neurovascular Disturbances · Neuroinflammation and Neurodegeneration Mechanisms · S100 Proteins and Annexins
Introduction
Traumatic brain injury (TBI) affects an estimated sixty-nine million people globally each year[1]. In 2022 alone, over twenty thousand United States service members from the Army, Navy, Air Force, and Marines, suffered from a TBI[2]. Approximately 80% of all TBI cases are classified as mild, in which diffuse pathologies that are difficult to discern via molecular imaging prevail. One of the pathological hallmarks of mild TBI is diffuse axonal injury, wherein axons are disrupted over time and progress to disconnection resulting in a proximal axonal swelling that is still connected to the neuronal soma and a distal axonal segment that degenerates via Wallerian degradation[3]. Additionally, microglia, the innate immune cell of the central nervous system, have been shown to be activated following TBI in both humans[4–9] and animals[10–14] and have been linked to cognitive changes following TBI[5]. Activated microglia fall on a spectrum from pro-inflammatory to anti-inflammatory with functions that can promote tissue neurodegeneration or neuroprotection[15–21].
While previous studies have identified various pro and anti-inflammatory pathways upregulated following TBI, non-phagocytic physical interactions between activated microglia and adjacent neurons have only recently begun to be investigated[10, 22–29]. Previous studies from our group using a micropig model of TBI found microglial processes converging onto the injured proximal axonal segment in a phenomenon called microglial process convergence (MPC)[27, 30]. This MPC does not appear to involve phagocytosis[27] and was not found in our rat model of TBI[26]. Specifically, in pigs, the number of activated microglial processes contacting injured proximal axonal swellings was nearly twice that observed for non-injured myelinated fibers at 6hrs following a diffuse TBI generated using the central fluid percussion injury model[27]. This MPC significantly increased from 6hrs to 1 day post injury[26].
However, in rats, there were far fewer microglial processes contacting injured axonal swellings compared to non-injured myelinated fibers following TBI, indicating that MPC might be a phenomenon associated with higher order gyrencephalic brains[26]. To investigate the potential that TBI-induced MPC onto injured axons occurs in the human brain, in the current study we quantitatively assessed the prevalence of MPC onto injured axonal swellings and intact axonal segments in human postmortem brain tissue.
Results
Microglial Process Convergence Increases Following Traumatic Brain Injury and Axonal Injury
To investigate the potential for microglial process convergence occurring on injured axonal swellings or intact myelinated fibers in the human brain, multiplexed immunohistochemistry against APP to visualize injured axons, MBP to visualize intact myelinated axons, and Iba-1 to visualize microglia processes was done on human postmortem tissue from the DoD/USU tissue repository. When all fibers that were analyzed across all cases were collated as APP + injured axonal swelling or MBP + intact myelinated fibers, it was found that more Iba-1 + microglial processes/um of the perimeter were in direct contact with APP + axonal swellings compared to MBP + intact myelinated fibers (Fig. 1; U = 32,240, p = 2.4×10^−4^). The paraffin sections were ~ 5μm thick, precluding the ability to perform 3D reconstructions of the axonal swellings, as we had done for our previous studies[26, 27]. As a single 2D image of the axonal segments is likely to be missing processes that are out of the plan of section, we also investigated the number of Iba-1 + microglial processes that were within 5um of the axonal segments. More microglial processes were found within 5um of APP + axonal swellings compared to MBP + intact myelinated fibers (Fig. 1; U = 34,260, p = 2.99×10^−6^), indicating that more microglial processes are close to the injured axonal swellings.
After completing this initial analysis of the overall comparison between APP + swellings and MBP + myelinated fibers, the cases were un-blinded. Following case unblinding, it was discovered that some APP + axonal swellings were identified in control individuals and some MBP + myelinated fibers were analyzed from individuals that suffered a TBI (Fig. 2). Therefore, the analyzed fibers were organized into four groups: 1) MBP + intact myelinated fibers in control tissue, 2) MBP + intact myelinated fibers in TBI tissue, 3) APP + axonal swellings in control tissue, and 4) APP + axonal swellings in TBI tissue. When the data was stratified by axonal injury and TBI the finding of more microglial process convergence occurring directly onto APP + injured axonal swellings, specifically within TBI tissue (χ^2^(3) = 15.53, p = 0.001; Fig. 3). There were also significantly more microglia processes within 5μm of APP + injured swellings in both control and TBI tissue as compared to MBP + fibers in either injured or control postmortem samples was maintained (χ^2^(3) = 21.97, p = 6.61×10^−5^; Fig. 3).
The morphology between MBP + intact axonal segments and APP + axonal swellings was significantly different (χ^2^(3) = 12.2, p = 0.007), with the APP + axonal swellings within TBI tissue having smaller perimeters compared to the MBP + intact axonal fibers within control tissue (p = 0.003). The APP + axonal swellings in both control and TBI tissue had significantly higher circularity indices compared to the MBP + axonal fibers in either control or TBI tissue (χ^2^(3) = 332.35, p = < 0.001; Fig. 4). While the MBP + intact axonal fibers within TBI tissue were more circular than the MBP + axonal fibers in control tissue (p < 0.001), the morphology of APP + axonal swellings within control cases, however, were consistent with the APP + axonal swellings in TBI cases (Fig. 4; Perimeter p = 0.08; circularity p = 0.83).
To validate the by eye counts of Iba-1 microglial process puncta onto the APP + axonal swellings, the integrated density/intensity of Iba-1 microglial processes in the region of the APP + axonal swelling or MBP + myelinated fiber was assessed. The intensity of Iba-1 labeling was significantly higher in APP + injured axonal swellings compared to control MBP + axons (χ^2^(3) = 56.217, p = 3.78×10^−12^; Fig. 5). Specifically, there were not differences in the intensity of Iba-1 between APP + injured axons (p = 0.72) nor between MBP + axonal segments (p = 0.99) within control tissue compared to TBI tissue. There was significantly higher Iba-1 integrated density within APP + axonal swellings compared to MBP + intact axonal segments regardless of TBI status of the case (MBP + intact fiber in control cases vs. APP + swellings in control cases p < 0.001; MBP + intact fiber in TBI cases vs. APP + swellings in TBI cases p < 0.001; Fig. 5). This finding validated the counts of microglial process convergence onto injured axonal swelling, demonstrating increases in microglial process convergence onto APP + injured axonal swellings in the human brain.
Discussion
The current study demonstrates that microglial processes converge onto injured axonal swellings in the human brain following TBI. The perimeters of the axonal segments were relatively consistent across non-injured MBP + myelinated fibers in both control and TBI tissue as well as APP + injured axonal swellings within control tissue. The APP + injured axonal swellings within TBI tissue had significantly lower perimeters than the MBP + intact myelinated axonal segments within control tissue. The number of Iba-1 + microglial processes in direct contact with the axonal segment, however, was significantly higher onto APP + axonal swellings compared to MBP + non-injured axonal segments, despite the lower perimeter available for contact. There were also significantly more microglial processes within 5μm of the injured axons compared to the non-injured MBP + axonal segments. Our group previously observed MPC in the micropig brain acutely following a central fluid percussion diffuse TBI[27]. This phenomenon; however, was not recapitulated following TBI in the rat[26], indicating that MPC onto injured axons might be species specific. The current findings that MPC onto injured axonal swellings occurs in human postmortem tissue, validates our previous findings in the micro pig and indicates that MPC following TBI might primarily manifest in higher order gyrencephalic brains. These findings show that MPC is a phenomenon that occurs in the human population, necessitating further investigation.
Diffuse axonal injury (DAI) or traumatic axonal injury (TAI) is one of the hallmark pathologies of mild TBI[31–36]. It was originally thought that DAI occurs following TBI due to the mechanical shearing of the axons. However, this is only true for a subset of axons that are observed to be injured within minutes of TBI, which is referred to as primary axotomy. Rather, secondary axotomy/axonal injury, which occurs sub-acutely after injury in rodent models of TBI is the phenomenon that is typically investigated. Secondary axonal injury involves accumulation of the cytoskeleton and organelles within the axons[37–39]. Specifically, the tensile forces of the TBI cause axonal alterations that allow an influx of calcium into the axon. This calcium influx leads to activation of cysteine protease pathways, which leads to degradation of the cytoskeleton and ultimately lead to a reactive axonal swelling as anterogradely transported proteins and organelles pool at the end of the proximal axon[38–40]. In the early 1990s immunohistochemistry against the anterogradely transported protein, APP, was found to efficiently label the proximal injured axonal segment where it pooled[41, 42]. Immunohistochemical labeling of APP has since become the gold standard for identifying DAI pathologically. Secondary axonal injury is typically studied in rodents 6–24 hours following injury, when it is most prevalent[43, 44]. Within higher order animals DAI appears to be prevalent starting hours following injury and peaking at 1w post-TBI[13], however DAI has been shown to last up to 6 months in a pig model of TBI[45] and has been observed in human postmortem tissue from people several years following a TBI[46, 47].
Many recent studies have demonstrated the impact of inflammatory cascades in regulating behavioral morbidities, general pathology, and neuronal function in both the normal brain and in various disease states, including TBI[21, 48, 49]. Neuroinflammation has been demonstrated in various brain regions in the human population chronically following TBI[5, 9, 20, 50]. Microglia, the innate immune cells of the brain, are critical mediators of these TBI-induced neuroinflammatory processes[16, 51–57]. Microglia have been shown to contact specific areas of the axon in the mouse brain during homeostasis including, the nodes of Ranvier[58], the axon initial segment[10], synapses[59], and neuronal soma[60]. Many studies using rodents have indicated that reduction or elimination of activated microglia and/or targeting various neuroinflammatory signaling pathways ameliorates downstream pathology and behavioral morbidity[24, 61–64]. Conversely, other studies have also found that anti-inflammatory microglial activation is necessary and potentially advantageous[17, 54, 65–70]. These studies demonstrated activated microglia can secrete neurotrophic factors[15, 71–73], which would suggest a potential ameliorative effect of microglia following injury in some cases. Additionally, recent studies have shown that microglia physical contacts play a role in regulating neuronal activity, either increasing activity following anesthesia[74, 75] or decreasing activity following epileptiform activity[60, 67].
A study by Schirmer et. al. investigating human post-mortem tissue found a significant positive correlation between the density of microglia and axonal outgrowth as well as the duration of patient survival following TBI, however, they did not quantitatively investigate the physical interactions between microglia and injured axons[76]. Another study done in vitro and in rats following a spinal cord injury found that exosomes from anti-inflammatory microglia increased neurite outgrowth in vitro and increased GAP43 expression in vivo, indicating that microglia could play a role in axonal outgrowth[17].
Microglia have been observed within physical proximity to injured axonal swellings in human postmortem tissue[76–78]. Oehmichen et al. observed an increase in CD-68 + microglial cells areas of axonal injury in the white matter at least five days post-TBI in human postmortem tissue, however, they only observed limited physical interactions between the CD68 + cells and the APP + axonal segments[77]. Ryu et al. qualitatively identified areas in which Iba-1 + microglia were in proximity to APP + injured axons in postmortem tissue from individuals following both motor vehicle accident and blast induced TBI[78]. Although, these previous studies indicated that there could be direct physical interactions between microglia and injured axonal segments, our current study is the first to quantitatively show that MPC occurs onto injured proximal axonal segments in human brain tissue following TBI.
We do appreciate that there are limitations to the current study, mainly, that all cases were from male doners. There is evidence that males and females respond to TBI differently[79]. A recent study also found that the burden of axonal injury following a diffuse TBI in a pig model was significantly higher in females compared to age matched males[80]. Further, microglia have been shown to be different in males and females[81–85]. Therefore, investigations into MPC in both the male and female population should be done to fully appreciate the prevalence of MPC onto injury axons in the human brain. Additionally, these sections were only 15μm thick, precluding a 3D investigation of MPC, as was done in our previous micro pig studies[26, 27]. It is likely that the numbers of microglial processes we found converging onto the axonal segments in the current study were artificially lower than they might actually be due to the section thickness. Therefore, investigations using thicker tissue in which 3D reconstructions could be done, would be warranted to glean a better appreciation of the degree of MPC onto injured axonal swellings in human postmortem tissue of both males and females. Despite these limitations, the current study is the first to quantitatively demonstrate MPC onto injured axons in the human brain following TBI. These findings indicate that MPC is a component of human TBI and that further studies exploring the phenotypes and overall roles of the microglia involved in MPC following TBI should be investigated.
Methods
Samples
Human brain samples were acquired from the Department of Defense (DoD)/Uniform Services University (USU) Tissue Repository. All cases were from males between 26–69 years old (median age of 36 years) with a maximum postmortem interval of 1day. All identifiers were removed from the samples. Tissue was paraffin embedded and sectioned at 15um. Slides containing areas demonstrating axonal injury regardless of brain region were used for this study. A total of 11 human brain samples were used. Of these samples, 6 were confirmed TBI cases with demonstrated areas of axonal injury and 5 were controls. All study staff was blinded to case group throughout the labeling, imaging, and analysis.
Immunohistochemistry
To visualize the interactions between microglia and injured or intact axonal segments in the human brain multiplexed fluorescent immunohistochemistry was performed. To identify injured axons, an antibody against amyloid precursor protein (APP) was used, which indicates axonal transport issues indicative of axonal injury [13, 14, 58]. To visualize microglia, an antibody against ionized calcium-binding adaptor molecule 1 (Iba-1) was used. Intact, non-injured axonal fibers were visualized using an antibody against myelin basic protein (MBP).
In this procedure, sections were deparaffinized by incubating slides in progressively more concentrated alcohols. Antigen retrieval was done by steaming the tissue in pH 6.0 citric acid buffer for 30min. Tissue was then blocked and permeabilized at room temperature in 5% normal goat serum (NGS), 2% bovine serum albumin (BSA) and 1.5% triton in phosphate buffered saline for 2hr followed by overnight incubation with a rabbit antibody against microglial Iba-1 (1:200; Cat.#019–19741 Wako; Richmond, VA, USA) at 4C° in 5% NGS/2% BSA/0.5% triton. Tissue was washed with 1%NGS/1%BSA in PBS at least six times prior to secondary antibody incubation with Alexa Fluor 488-conjugated goat anti-rabbit IgG (1:700; Cat.# A- 11008, Life Technologies, Carlsbad, CA, USA) in 1%NGS/1%BSA/PBS at room temperature for 2hr. Tissue was washed in PBS at least four times prior to overnight incubation with a mouse antibody against the 22C11 clone of APP (1:200; Cat.#14–9749-82, ThermoFisher Scientific, Waltham, MA, USA) in 5% NGS/2% BSA/0.5% triton at 4C°. Tissue was washed with 1%NGS/1%BSA in PBS at least six times prior to the next secondary antibody incubation with Alexa Fluor 647-conjugated goat anti-mouse IgG (1:700; Cat.# A- 21237, Life Technologies, Carlsbad, CA, USA) in 1%NGS/1%BSA/PBS at room temperature for 2hr. Iba-1 and APP labeled tissue was washed in PBS at least four times prior to overnight incubation with a rat antibody against MBP (1:200; Novus) at 4C° in 5% NGS/2% BSA/0.5% triton. Tissue was washed with 1%NGS/1%BSA in PBS at least six times prior to the third secondary antibody incubation with Alexa Fluor 568-conjugated goat anti-rat IgG (1:700; Cat.# A- 11077, Life Technologies, Carlsbad, CA, USA) in 1%NGS/1%BSA/PBS at room temperature for 2hr. Tissue was washed in PBS at least four times. Multiplex labeled tissue was coversliped with Vectashield hard-set mounting medium with Dapi (Cat.#H-1500; Vector Laboratories, Burlingame, CA, USA).
Microglial Process Convergence Analysis
The fluorescently immunolabeled slides were imaged on the Keyence BZ-X800 microscope (Keyence Corporation of America, Itasca, IL, USA) at 40X magnification. One section was analyzed for each case by an investigator blinded to group. A navigation super-image was generated using the far-red channel in which the APP + injured axonal swellings could be visualized. Images containing at least 1 APP + injured axonal swellings were captured or no APP + swellings but clean MBP labeling were captured. Fewer MBP only images were captured, as there were several analyzable MBP + intact myelinated fibers in each captured image, whereas there were few analyzable APP + axonal swellings in each captured image. At least 25 images were taken for most cases, however, only 13 images were captured for 1 case as no APP + swellings were identified. Across all samples a total of 161 non-injured axonal segments from controls, 36 non-injured axonal segments from injured samples, 105 injured axonal swellings from controls, and 173 injured axonal swellings from injured samples were analyzed for the current study. Fiji Image J software (National Institute of Health, Bethesda, MD, USA) was used to evaluate the 2D images. Image scales were set to 5.3 pixels/um
To assess the interaction between microglia and injured axonal swellings, the APP + axonal swelling was traced using the freehand tool and measured for perimeter, area, shape descriptors (aspect ratio, circularity, round, solidity), integrated density, and mean grey value. The number of microglial processes and puncta that were directly touching the APP + axonal swelling was counted by hand. Then, the region encircling the APP + axonal swelling was enlarged by 5um and the microglial processes and puncta within the enlarged region was counted by hand.
In order to visualize the interaction between microglia and intact axonal segments, a random number generator was used to generate x,y coordinates to choose a MBP + axonal segment on the image. The axonal segment was traced with the freehand tool and measured for perimeter, area, shape descriptors (aspect ratio, circularity, roundness, and solidity), integrated density, and mean grey value. The number of microglial processes and puncta that were directly touching the MBP + intact axonal segment was counted by hand. Then, the region encircling the axonal segment was enlarged by 5um and the number of microglial processes and puncta within the enlarged region were counted by hand.
Statistical Analysis
The statistics were run using IBM SPSS software (IBM Corp., Armonk, NY). A Shapiro-Wilk test was conducted to test for normality of the data. As the data was not normally distributed, a Mann-Whitney U test was used to test differences between all APP + injured axonal swellings and all MBP + intact axonal segments. A Kruskal-Wallis test was run to assess differences across multiple groups. A Bonferroni post hoc was used to correct for multiple pairwise comparisons. Statistical significance was set to a p-value of < 0.05. Data is presented as means and standard error of the mean. All raw data is included in Supplemental table 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dewan MC, Rattani A, Gupta S, Baticulon RE, Hung Y-C, Punchak M, Estimating the global incidence of traumatic brain injury. Journal of Neurosurgery. 2018;130:1080–97.29701556 10.3171/2017.10.JNS 17352 · doi ↗ · pubmed ↗
- 2DOD TBI Worldwide Numbers [Internet]. Military Health System. [cited 2024 May 28]. Available from: https://www.health.mil/Military-Health-Topics/Centers-of-Excellence/Traumatic-Brain-Injury-Center-of-Excellence/DOD-TBI-Worldwide-Numbers
- 3Kelley BJ, Farkas O, Lifshitz J, Povlishock JT. Traumatic axonal injury in the perisomatic domain triggers ultrarapid secondary axotomy and Wallerian degeneration. Exp Neurol. 2006;198:350–60.16448652 10.1016/j.expneurol.2005.12.017 · doi ↗ · pubmed ↗
- 4Coughlin JM, Yuchuanwang Y, Minn I, Bienko N, Ambinder EB, Xu X, Imaging of glial cell activation and white matter integrity in brains of active and recently retired national football league players. JAMA Neurology. 2017;74:67–74.27893897 10.1001/jamaneurol.2016.3764 PMC 5504689 · doi ↗ · pubmed ↗
- 5Coughlin JM, Wang Y, Munro CA, Ma S, Yue C, Chen S, Neuroinflammation and brain atrophy in former NFL players: An in vivo multimodal imaging pilot study. Neurobiology of Disease. 2015;74:58–65.25447235 10.1016/j.nbd.2014.10.019PMC 4411636 · doi ↗ · pubmed ↗
- 6Little DM, Kraus MF, Joseph J, Geary EK, Susmaras T, Zhou XJ, Thalamic integrity underlies executive dysfunction in traumatic brain injury. Neurology. 2010;74:558–64.20089945 10.1212/WNL.0b 013e 3181 cff 5d 5PMC 2830915 · doi ↗ · pubmed ↗
- 7Boche D, Perry VH, Nicoll J a R. Review: Activation patterns of microglia and their identification in the human brain. Neuropathology and Applied Neurobiology. 2013;39:3–18.23252647 10.1111/nan.12011 · doi ↗ · pubmed ↗
- 8Neumann KD, Seshadri V, Thompson XD, Broshek DK, Druzgal J, Massey JC, Microglial activation persists beyond clinical recovery following sport concussion in collegiate athletes. Front Neurol [Internet]. 2023 [cited 2024 May 1];14. Available from: 10.3389/fneur.2023.1127708/full PMC 1008013237034078 · doi ↗ · pubmed ↗