Increased expression of metabolism and lysosome-associated genes in a C. elegans dpy-7 cuticle furrow mutant
Aiden Fong, Michael Rodriguez, Keith Patrick Choe

TL;DR
This study shows that a C. elegans cuticle mutant increases specific genes related to metabolism and lysosomes, possibly through mechanisms separate from SKN-1.
Contribution
The study confirms RNAseq findings using RT-qPCR and reveals that gene activation in cuticle mutants is not dependent on SKN-1.
Findings
Five genes related to redox balance, ATP homeostasis, and lysosome function are upregulated in dpy-7 mutants.
SKN-1 RNAi does not reduce the expression of these genes, indicating alternative regulatory pathways.
The study highlights a potential new regulatory mechanism for gene expression in response to cuticle damage.
Abstract
The collagen-based epidermal ‘cuticle' of Caenorhabditis elegans functions as an extracellular sensor for damage that regulates genes promoting osmotic balance, innate immunity, and detoxification. Prior studies demonstrate that SKN-1 , an ortholog of the mammalian Nrf transcription factors, activates core detoxification genes downstream from cuticle damage. Prior RNAseq data suggested that expression of five genes with functions in redox balance, ATP homeostasis, and lysosome function ( gst-15 , gst-24 , cyts-1 , argk-1 , and mfsd-8.4 ) were increased in a cuticle collagen mutant; this study employed RT-qPCR to verify this observation and to test the role of SKN-1 . Activation of all five genes was verified in dpy-7 mutants, but none were reduced by skn-1 (RNAi) suggesting parallel or distinct regulatory mechanisms.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
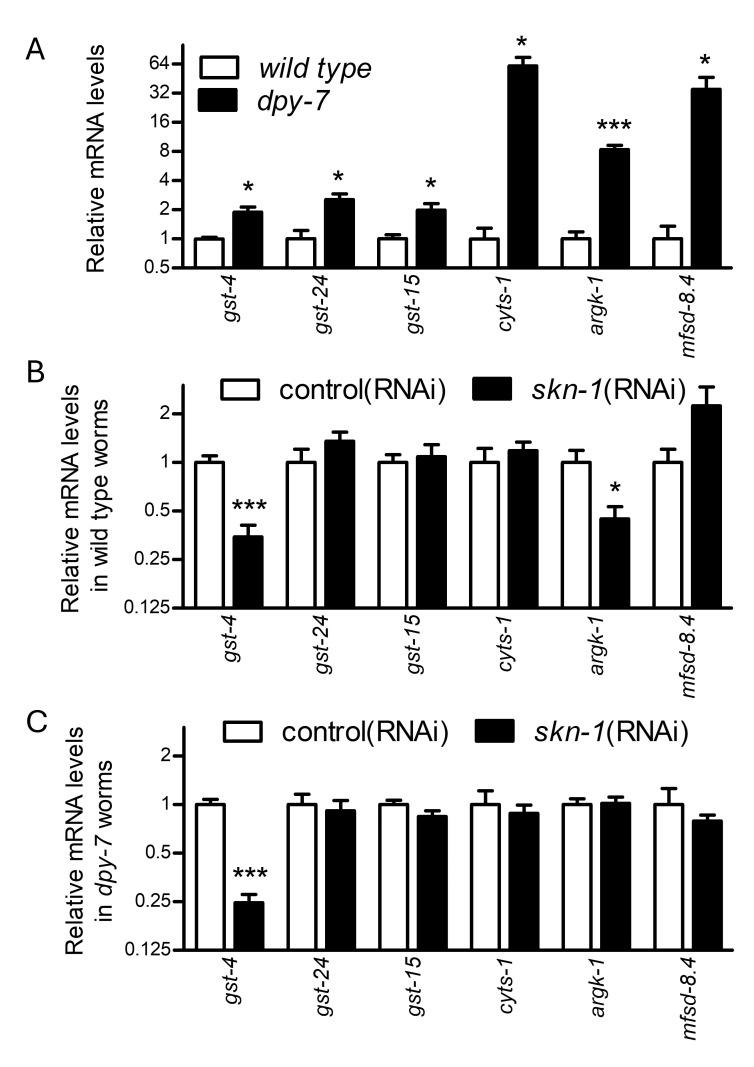 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBiochemical and Molecular Research
Description
Animal cells rely on conserved signaling mechanisms to sense adverse environmental conditions and modulate expression of cytoprotective genes. Intracellular sensing and signaling pathways that regulate cytoprotective genes are well-studied (Choe et al. 2009, Blackwell et al. 2015, Dietrich et al. 2017, Urso and Lamitina 2021, Pujol and Ewbank 2022) , but mechanisms outside of cells in the tissues that interact directly with the environment are poorly understood.
Collagenous extracellular matrices (ECMs) are ubiquitous in animal organs and serve as barriers to the environment in epidermal tissues. Although originally hypothesized to be inert physical scaffolds, ECMs are now understood to be dynamic structures that regulate organogenesis and tissue remodeling (Maquart et al. 2004, Rozario and DeSimone 2010, Clause and Barker 2013) . In mammalian lungs, peptide fragments of digested collagen and other ECM components are sensed by cell receptors and regulate immune responses, wound repair, and cell proliferation (Gaggar and Weathington 2016, Patel and Snelgrove 2018) .
Nematodes are enclosed in a collagen-rich exoskeleton called the ‘cuticle' (Chisholm and Hsiao 2012) . We and others have identified the cuticle as a putative extracellular sensor for damage that regulates three stress responses (Lamitina et al. 2006, Wheeler and Thomas 2006, Pujol et al. 2008, Dodd et al. 2018, Wimberly and Choe 2022) . This ECM damage response is induced by disruption of circumferential bands of collagen in the cuticle known as annular furrows; silencing or mutation of any one of six collagens required for furrow formation activates the responses (Dodd et al. 2018) . Understanding this ECM damage response will help define a novel mode of stress-response signaling and relevant homeostasis mechanisms. The mechanism for sensing cuticle damage is not known, but recent studies provide insights. Full activation of stress responses requires atypical membrane-associated kinase DRL-1 (Wimberly and Choe 2022) . Plasma membrane folds named ‘meisosomes' were recently identified and shown to be associated with furrows in epidermal cells and could be involved in signaling (Aggad et al. 2023) .
Candidate genes activated by furrow disruption have been identified with microarrays and RNAseq; they are highly enriched for functions in canonical osmotic, detoxification, and innate immune responses and largely exclude other core stress responses (Pujol et al. 2008, Rohlfing et al. 2010, Dodd et al. 2018, Scolaro et al. 2019) . DPY-7 is a collagen localized to furrows and is required for periodic organization of the cuticle and epidermal cortical cytoskeleton and attachment of cuticle to the epidermal plasma membrane (Cox et al. 1980, McMahon et al. 2003, Thein et al. 2003, Dodd et al. 2018, Chandler and Choe 2022, Aggad et al. 2023) . In the current study, we used RT-qPCR to independently verify activation of genes predicted to function in detoxification, redox balance, and energy metabolism in * dpy-7 ( e88 ) * mutant worms; we also used RNAi to test the requirement of transcription factor SKN-1 , a master regulator of detoxification that we previously showed to mediate activation of * gst-4 * and * gst-10 * in the same strain (Dodd et al. 2018) . Sequencing of the * skn-1 * ORFeome clone that we used confirmed that it covers exons 1-4 of * skn-1 c * , which overlaps at least 227 bases of all predicted transcript variants (i.e., * skn-1 a, b, c, * and d) .
As shown in Figure 1A, * gst-15 * , * gst-24 * , * cyts-1 * , * argk-1 * , and * mfsd-8.4 * were all verified to be induced in * dpy-7 * worms; direct SKN-1 target gene * gst-4 * was previously studied and is included here as a positive control (Dodd et al. 2018) . * cyts-1 * is predicted to encode a cysteine synthase and was induced 61.2-fold; cysteine is a precursor for glutathione, a major cellular redox buffer (Lapenna 2023) . * gst-15 * and * gst-24 * are predicted to encode glutathione S-transferase enzymes and they were induced 1.8-2.5-fold; glutathione S-transferases conjugate glutathione to small molecules reducing toxicity and increasing solubility (Salinas and Wong 1999) . Activation of these detoxification and redox homeostasis genes is expected to help compensate for a compromised barrier ECM that is permeable to xenobiotics (Dodd et al. 2018) . Surprisingly, only expression of positive control gene * gst-4 * was reduced by * skn-1 * RNAi in wild type and * dpy-7 * worms (Figures 1B-C).
argk-1 * is predicted to encode a creatine kinase and was induced 8.3-fold in * dpy-7 * worms ( Figure 1A ); creatine kinases function to buffer and transport energy and are enriched in muscle and neurons (Sumien et al. 2018) . In human cells and aquaculture turtles, infection has been linked to upregulation of creatine kinase expression, potentially functioning to buffer ATP demands in tissues mounting immune-responses (Li et al. 2020) . Single cell expression data suggest that * argk-1 * is expressed in the hypodermis and intestine (Paker 2019). Worms with disrupted furrows synthesize high levels of the energetically expensive osmolyte glycerol in these same tissues (Lamitina et al. 2006, Possik et al. 2015, Dodd et al. 2018) ; activation of * argk-1 * could function to buffer ATP levels under these conditions. Basal expression of * argk-1 * was reduced by * skn-1 * RNAi, but not in * dpy-7 * worms (Figures 1B-C).
mfsd-8.4 * encodes a homolog of lysosomal chloride ion membrane transporter MFSD8 (Wang et al. 2021) and was induced 35.1-fold in * dpy-7 * worms. MFSD8 function and regulation are poorly understood; MFSD8 mutations are associated with neuronal ceroid lipofuscinoses disease in humans and with defects in protein secretion and lysosomal function in amoeba (Kirola et al. 2022, Yap et al. 2022) . Single cell expression data suggest that * mfsd-8.4 * is expressed in interneurons under basal conditions (Paker 2019). Lysosomes are remodeled during molting and impairing lysosome function causes molting defects (Miao et al. 2020) . If * mfsd-8.4 * is expressed in epidermal cells of * dpy-7 * worms, it could function to promote digestion of damaged cell components or secretion of proteins involved in regulation of cuticle remodeling. Expression of * mfsd-8.4 * was not reduced by * skn-1 * RNAi (Figures 1B-C).
Our results expand the diversity of genes activated by the cuticle damage response to include cysteine synthesis, energy metabolism, and lysosomal function. Unlike * gst-4 * and some other detoxification genes (Dodd et al. 2018) , none of these newly verified responses to * dpy-7 * mutation were dependent on * skn-1 * . There could be redundant or distinct mechanisms of activation; future studies could test the role of transcription factors ELT-3 and STA-2 that we and others previously showed to mediate parts of the response to * dpy-7 * mutation (Zugasti et al. 2014, Dodd et al. 2018). Creatine kinases and MFSD8 play important roles in human physiology and pathophysiology. Strong activation in * dpy-7 * worms provides a model for understanding regulation and function in the context of stress response.
Methods
Worms were maintained on OP50 E. coli on NGM agar at 20°C with standard conditions. For experiments, worm eggs were released with bleach and raised on dsRNA-expressing *E. coli * ( HT115 (DE3)); clone pPD129.36 (LH4440) encoding a 202-bp dsRNA not homologous to C. elegans genes was used as a control and the * skn-1 * dsRNA clone was derived from the ORFeome library (Open Biosystems, Huntsville, AL) as we have described previously (Choe et al. 2009, Tang and Choe 2015) .
Worms were collected and processed for RT-qPCR at the L4 stage (to avoid embryos) as we have described previously (Scolaro et al. 2019, Piloto et al. 2022) with slight modifications. After lysis, gDNA was degraded using DNase (Thermo Fisher EN007). Primers were designed using Primer-BLAST (U.S. National Library of Medicine) and span intron splice junctions. mRNA levels were normalized to * rpl-2 * and to controls using the delta-delta Ct method. Statistical significance was analyzed with Students t-tests and P-values and were corrected for multiple comparisons with Benjamini-Hochberg adjustments.
Reagents
Strains:
C. elegans strains used were wild-type N2 Bristol and CB88 * dpy-7 ( e88 ) * , which are both available at the *Caenorhabditis * Genetics Center.
Primers:
rpl-2 * – CTTTCCGCGACCCATACAA and CACGATGTTTCCGATTTGGAT
gst-4 * – TCCGTCAATTCACTTCTTCCG and AAGAAATCATCACGGGCTGG
gst-24 * – GGAGCGTTGAAGCCAAAAAC and TTGGGGGATTTCGAAGCCAT
gst-15 * – AGAAAATGAGAGACAAAACCCCA and AGATTGGGGGATGTCGAAGC
cyts-1 * – TTCGCCGTAGTTTCTGAGGA and CGGAGAGCAGTTGGTACCTTTAT
argk-1 * – CTGCGATAAGCTTGACCTCCA and TCCGAGACGAGCCCTGTTA
mfsd-8.4 * – CCAGACAAGACAGGAAGCAGT and AGAATCGTGGCAATGAATCCAG
RNAi:
HT115 *E. coli * with empty plasmid pPD129.36 (LH4440) or with the ORFeome * skn-1 * clone that covers * skn-1 c * exons 1-4 and overlaps with all predicted transcript variants (i.e., * skn-1 a-d) *
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aggad D Brouilly N Omi S Essmann CL Dehapiot B Savage-Dunn C Richard F Cazevieille C Politi KA Hall DH Pujol R Pujol N 2023313 Meisosomes, folded membrane microdomains between the apical extracellular matrix and epidermis.Elife 1210.7554/e Life.7590636913486 PMC 10010689 · doi ↗ · pubmed ↗
- 2Blackwell TK Steinbaugh MJ Hourihan JM Ewald CY Isik M 201585 SKN-1/Nrf, stress responses, and aging in Caenorhabditis elegans.Free Radic Biol Med 88Pt B 0891-584929030110.1016/j.freeradbiomed.2015.06.00826232625 PMC 4809198 · doi ↗ · pubmed ↗
- 3Chandler LM Choe KP 2022114 Extracellular matrix regulation of stress response genes during larval development in Caenorhabditis elegans.G 3 (Bethesda)121110.1093/g 3journal/jkac 22136000892 PMC 9635657 · doi ↗ · pubmed ↗
- 4Chisholm AD Hsiao TI 2012619 The Caenorhabditis elegans epidermis as a model skin. I: development, patterning, and growth.Wiley Interdiscip Rev Dev Biol 161759-768486187810.1002/wdev.7923539299 PMC 3607643 · doi ↗ · pubmed ↗
- 5Choe KP Przybysz AJ Strange K 200939 The WD 40 repeat protein WDR-23 functions with the CUL 4/DDB 1 ubiquitin ligase to regulate nuclear abundance and activity of SKN-1 in Caenorhabditis elegans.Mol Cell Biol 29100270-73062704271510.1128/MCB.01811-0819273594 PMC 2682033 · doi ↗ · pubmed ↗
- 6Clause KC Barker TH 2013528 Extracellular matrix signaling in morphogenesis and repair.Curr Opin Biotechnol 2450958-166983083310.1016/j.copbio.2013.04.01123726156 PMC 3773047 · doi ↗ · pubmed ↗
- 7Cox GN Laufer JS Kusch M Edgar RS 198061 Genetic and Phenotypic Characterization of Roller Mutants of CAENORHABDITIS ELEGANS.Genetics 9520016-673131733910.1093/genetics/95.2.31717249038 PMC 1214229 · doi ↗ · pubmed ↗
- 8Dietrich N Schneider DL Kornfeld K 20171116 A pathway for low zinc homeostasis that is conserved in animals and acts in parallel to the pathway for high zinc homeostasis.Nucleic Acids Res 45200305-1048116581167210.1093/nar/gkx 76228977437 PMC 5714235 · doi ↗ · pubmed ↗