Expression of Kielin/chordin-like protein is regulated by BMP-2 in osteoblasts
Kazuki Toba, Atsushi Yamada, Kiyohito Sasa, Tatsuo Shirota, Ryutaro Kamijo

TL;DR
This study shows that BMP-2 increases Kcp expression in osteoblasts, which helps promote bone formation through positive feedback.
Contribution
The study reveals that Kcp is regulated by BMP-2 and contributes to osteoblast differentiation via SMAD signaling.
Findings
Kcp expression in osteoblasts is increased by BMP-2 in a concentration- and time-dependent manner.
SMAD signaling is involved in the regulation of BMP-2-induced Kcp expression.
Kcp partially regulates BMP-2-induced osteoblast differentiation and mineralization.
Abstract
Bone morphogenetic protein (BMP), an osteoinductive factor, is a cytokine that induces osteoblast differentiation and mineralization, and expected to be applicable for hard tissue reconstruction. Kielin/chordin-like protein (Kcp), a member of the family of cysteine-rich proteins, enhances BMP signaling. The present study found that expression of Kcp in osteoblasts was induced by BMP-2 in a concentration- and time-dependent manner. Up-regulation of Kcp by BMP-2 was inhibited by Dorsomorphin, a SMAD signaling inhibitor. The involvement of up-regulation of Kcp by BMP-2 in induction of osteoblast differentiation by BMP-2 was also examined, which showed that suppression of Kcp expression by si Kcp partially inhibited induction of osteoblast differentiation and mineralization by BMP-2. Together, these results suggest that Kcp induced by BMP-2 functions to provide positive feedback for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
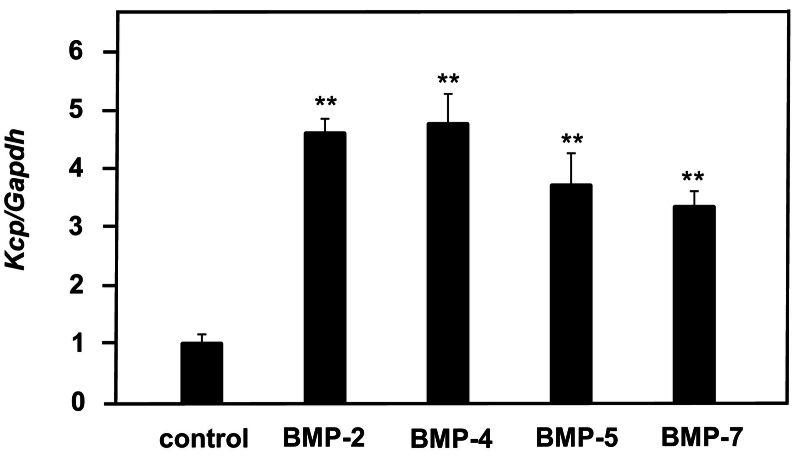 Figure 1
Figure 1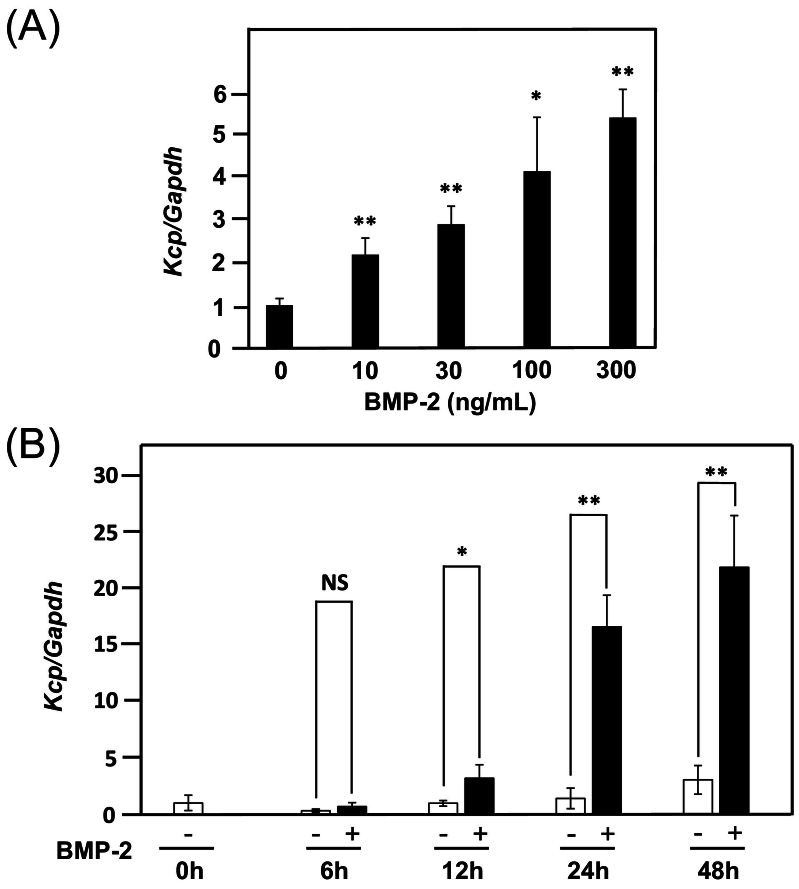 Figure 2
Figure 2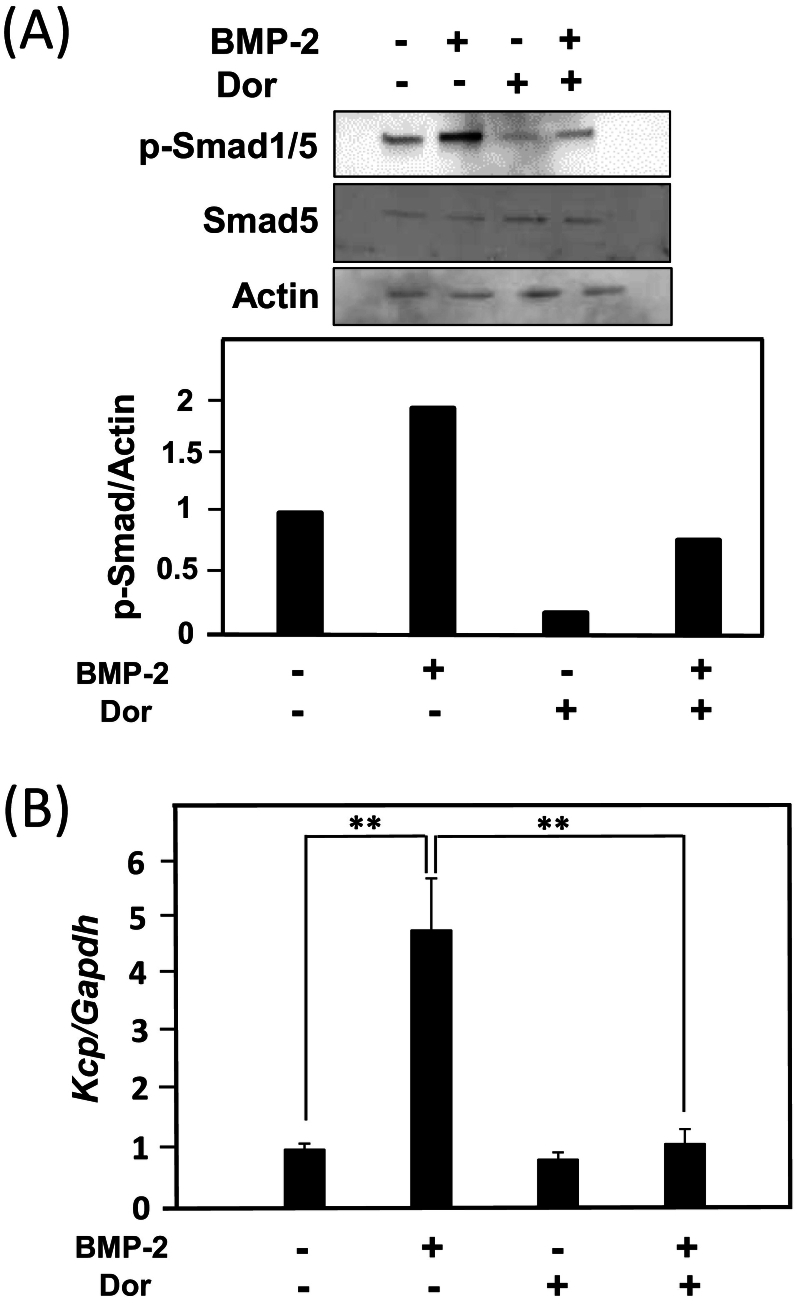 Figure 3
Figure 3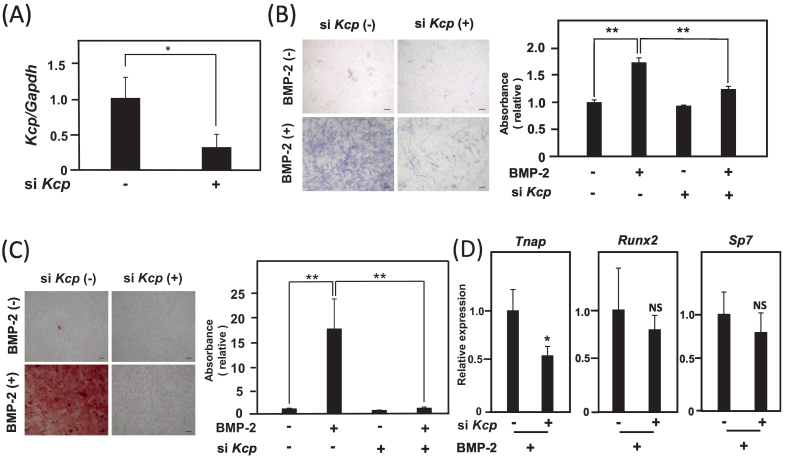 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
Topicsdental development and anomalies · Bone Metabolism and Diseases · Periodontal Regeneration and Treatments
Introduction
1
Bone morphogenetic proteins (BMPs) have important roles in embryogenesis, organogenesis, cell proliferation, and cell differentiation (Miyazawa et al., 2002). These proteins belong to the transforming growth factor-β (TGF-β) superfamily, with approximately 20 different BMP families identified and characterized thus far, and shown to regulate bone balance and osteogenic function by regulating osteoblast differentiation (Wan and Cao, 2005). BMPs are expected to be applied for hard tissue reconstruction, thus it is crucial to elucidate factors related to promotion of their activity for effective clinical applications (Zhu et al., 2022).
BMPs bind to the serine/threonine kinase receptors BMPR-I and II, four and three transmembrane receptors, respectively, for signal transduction. When BMPs bind to these receptors, BMPR-II phosphorylates BMPR-I (Ebendal et al., 1998), while activated BMPR-I phosphorylates the receptor-regulated Smads (R-Smads) Smad1, 5, and 8 (Gomez-Puerto et al., 2019). A phosphorylated R-Smad forms a heterotrimer with Smad4 and migrates into the nucleus to regulate expression of target genes. This SMAD-mediated BMP signaling is known to be inhibited by Dorsomorphin, which blocks the action of BMPR-I (Yu et al., 2008). It has also reported that BMP-2 activates members of the non-SMAD-mediated signaling pathway, including mitogen-activated protein kinase (MAPK), leading to induction of osteoblast differentiation (Chen et al., 2012).
Kielin/chordin-like protein (Kcp), which has 18 cysteine-rich domains and a high amino acid sequence homology to the BMP antagonist chordin, enhances BMP signaling by activating BMP receptors (BMPR-I) in the paracrine and promotes phosphorylation of intracellular Smads, while its physiological function is involved in attenuating renal fibrotic disease (Lin et al., 2005a). Recently, Kcp was shown to enhance BMP-2-induced osteoblast differentiation through SMAD signaling (Zhu et al., 2022; Nagasaki et al., 2023; Zhao et al., 2006). The present study was conducted to elucidate the regulatory mechanism of Kcp expression by BMP-2 in osteoblasts.
Materials and methods
2
Reagents
2.1
BMP-2 was purchased from R&D Systems (355-BM), BMP-4 from FUJIFILM Wako Pure Chemical Corporation (023–18,461), BMP-5 from R&D Systems (6176-BM), and BMP-7 from R&D Systems (5666-BP), Dorsomorphin was purchased from FUJIFILM Wako Pure Chemical Corporation (044–33,751). U0126 (MEK 1/2 inhibitor; 9903) was purchased from Cell Signaling, and SP600125 (JNK inhibitor; S1460) and SB203580 (p38 MAPK inhibitor; S1076) were purchased from Selleck.
Cell culture
2.2
MC3T3-E1, a mouse cranial crown-derived osteoblast cell line, was maintained in culture medium produced using 500 ml of α-MEM along with l-glutamine and phenol red medium (FUJIFILM Wako Pure Chemical Industries; 135–15,175), supplemented with 55 ml of fetal bovine serum (10 % FBS) (SIGMA-ALDRICH Corporation; 173,012) and 5 ml of penicillin-streptomycin (1 % penicillin-streptomycin) (Life Technologies Corporation, 15,240–062), at 37 °C in a CO_2_ incubator (5 % CO_2_, 95 % air). For collecting primary cultured osteoblasts, the skull of a one-day-old mouse was excised, then attached soft tissue was removed and placed in a 0.1 % collagenase/0.2 % dispase solution produced using 40 ml of culture medium supplemented with 40 mg of collagenase (0.1 % collagenase) and 80 mg of dispase (0.2 % dispase), then shaken at 37 °C for 5 min. After discarding the supernatant, fresh 0.1 % collagenase/0.2 % dispase solution was added, then shaking was continued at 37 °C for 10 min. The supernatant was obtained, and centrifuged and processed through a 40-μm cell strainer. Primary cultured osteoblasts were then maintained in culture medium at 37 °C in a CO_2_ incubator.
Kcp knockout mice
2.3
Kcp knockout mice (B6.Cg-KCP < tm1Ysas>/Rbrc, Accession No. CDB401K; https://large.riken.jp/distribution/mutant-list.html) were purchased from RIKEN BioResource Center (Ikeya et al., 2006). All protocols used for the animal experiments in this study were approved by the Showa University Animal Experiment Committee.
Quantitative RT-PCR
2.4
Total RNA was extracted from cells using TRIzol® Reagent (Life Technologies; 15,596,018), then cDNA was synthesized using ReverTra Ace® qPCR RT Master Mix (TOYOBO CO., LTD.; FSQ-201). Quantitative real-time PCR (95 °C for 1 min, followed by 40–50 cycles at 95 °C for 15 s and 60 °C for 30 s) was performed using THUNDERBIRD® Probe qPCR Mix (TOYOBO CO., LTD.; QPS-101). TaqMan™ IDs (Applied Biosystems) for gene expression assays were as follows: mouse Gapdh (Applied Biosystems; Mm99999915_g1), mouse Kcp (Applied Biosystems; Mm01159615_m1, Mm01163386_m1), mouse Tnap (Applied Biosystems; Mm00475834_m1), mouse Runx2 (Applied Biosystems; Mm00501584_m1), and mouse Sp7 (Applied Biosystems; Mm04209856_m1) (Aizawa et al., 2019).
Western blotting
2.5
MC3T3-E1 cells (5 × 10^5^ cells) were treated with a cell lysate sample buffer solution (1 ml) along with 3-Mercapto-1,2-propanediol (x4) (FUJIFILM Wako Pure Chemical Corporation; 196–16,142). Samples (25 μl) and marker proteins (25 μl) (Precision Plus Protein Western C Standards, BIORAD; 1,610,376) were separated into polyacrylamide gels using SDS-PAGE. After transfer, blocking was performed with 4 % skin milk (Difco Skim Milk, BD; 232,100) (Kinoshita et al., 2021). Smad5 (Smad5 D4G2 rabbit mAb, Cell Signaling TECHNOLOGY; 12,534), p-Smad1/5 (phospho-Smad1/5 Ser463/465; 41D10, rabbit mAb, Cell Signaling TECHNOLOGY; 9516), actin (SIGMA-ALDRICH; A5060), and anti-rabbit IgG horseradish peroxidase-linked whole donkey (GE Healthcare; NA934V) antibodies were used. Thereafter, reaction solution (ECL Prime Western Blotting Detection Reagent, Global Life Sciences Solutions Operations; RPN2232) treatment was performed and images were obtained using a VersaDoc IMAGING SYSTEM 5000 (BIORAD).
Knockdown by si Kcp
2.6
MC3T3E1 cells were transfected with RNAi max (Life Technologies Corporation; 13,778–150) for si Kcp in accordance with the protocol of the manufacturer. The respective oligos for Kcp were as follows: 5’-CCACAAUGGGCAGUCUUAUGGUCAU-3′ and 5’-AUGACCAUAAGACUGCCCAUUGUGG-3′.
ALP staining
2.7
MC3T3-E1 cells were introduced to si Kcp and seeded into 96-well plates (2 × 10^4^ cells/well), then cultured for 72 h in the presence or absence of BMP-2 (300 ng/ml). Cells were then fixed with 4 % paraformaldehyde (FUJIFILM Wako Pure Chemical Corporation; 163–20,145) for 30 min and washed with 1 X PBS. Next, they were stained with 10 ml of Tris-HCl buffer (200 mM, pH 8.5) containing 1 mg of naphthol AS-MX phosphate (Sigma-Aldrich) and 2.5 mg of Fast blue BB (Sigma-Aldrich). After washing with distilled water, observations were performed with a microscope.
ALP activity
2.8
MC3T3-E1 cells were cultured in 96-well plates and washed with 1 X PBS, then 1 % Nonidet P40 was added. After treatment at room temperature for one hour, the cells were disrupted with ultrasonic waves (Handy Sonic UR-21P). Next, 5 ml of ALP activity buffer produced using ALP solution buffer (1 mM MgCl_2_, 200 mM Tris-HCl) containing 24 mg of disodium p-nitrophenyl phosphate hexahydrate) was added to the lysate and absorbance (405 nm) was determined (Sasa et al., 2018).
Alizarin red staining
2.9
MC3T3-E1 cells were introduced to si Kcp and seeded into 96-well plates (2 × 10^4^ cells/well), then cultured for seven days in calcification culture medium, produced using 50 ml of culture medium with β-glycerophosphate (10 mM) and dexamethasone (10 nM), with the medium replaced every four days. The supernatant was collected and discarded, then cells were washed with 1 X PBS, fixed with 95 % methanol for 20 min at room temperature, and stained with an Alizarin red staining solution made with 1 g of Alizarin Red S (FUJIFILM Wako Pure Chemical Corporation; 011–01192) in 100 ml of distilled water (pH 6.3–6.5) for five minutes. After washing with distilled water, microscopic observations were performed. For quantitative analysis, the cell mixture was dissolved in a cetylpyridinium chloride solution, produced using 10 g of cetylpyridinium chloride (NAKARAI Chemical Corporation; 07905–05) in 100 ml distilled water, and absorbance at 570 nm was determined.
Histomorphometric analyses
2.10
Femora were obtained from Kcp knockout mice, then fixed in 70 % ethanol and subjected to Villanueva bone staining without decalcification (Ito Bone Histomorphometry Institute). Parameters related to bone structure were examined, including total bone volume per tissue volume (BV/TV, %), trabecular thickness (Tb.Th, μm), osteoid thickness (O·Th, μm), number of osteoblasts per bone surface (N.Ob/BS, N/mm), and number of osteoclasts per bone surface (N.Ob/BS, N/mm).
Statistical analysis
2.11
Data are expressed as the mean ± SD. Student's t-test was used for statistical analyses, with p < 0.05 considered to indicate significance. Experiments were performed at least twice.
Results
3
BMP-2 induces Kcp expression in concentration- and time-dependent manner
3.1
Kcp functions as an agonist to enhance BMP signaling (Lin et al., 2005a). Therefore, to examine the expression of Kcp when BMPs are applied, MC3T3-E1 cells were treated with 300 ng/ml of BMP-2, −4, −5, and − 7 for 24 h, then increases in expression were observed (Fig. 1). Similar experiments were conducted using mouse skull-derived primary cultured osteoblasts, with increased Kcp expression observed (Suppl. Fig. 1). These results indicated that BMPs induce Kcp expression in osteoblasts. Furthermore, when MC3T3-E1 cells were exposed to various concentrations of BMP-2 for 24 h, a concentration-dependent increase in Kcp expression was noted. As for the effects of BMP-2 treatment on Kcp for times ranging from 6 to 48 h, no increase in expression was observed after 6 h of treatment, while induction of expression was observed after 12 h (Fig. 2). It is thus suggested that BMP-2 induces Kcp expression in a concentration- and time-dependent manner.Fig. 1MC3T3-E1 cells were treated with 300 ng/ml of BMP-2, −4, −5 and − 7 for 24 h, then the expression level of Kcp was examined. Using a control Kcp/Gapdh value of 1, the relative ratio of Kcp/Gapdh after BMP treatment was determined. **p < 0.01 (Student's t-test).Fig. 1. Fig. 2(A) Various concentrations (10, 30, 100, 300 ng/ml) of BMP-2 were applied to MC3T3-E1 cells for 24 h, then the expression level of Kcp was examined. Using a control Kcp/Gapdh value of 1, the relative ratio of Kcp/Gapdh when treated with BMP-2 at each concentration was determined. (B) Time course analysis of the effects of BMP-2 (300 ng/ml) on Kcp gene expression from 6 to 48 h. *p < 0.05, **p < 0.01 (Student's t-test). NS: not significant.Fig. 2
BMP-2 induces Kcp expression through SMAD signaling
3.2
Next, to investigate whether induction of Kcp expression by BMP-2 occurs via activation of SMAD signaling, the mode of Kcp expression following treatment with Dorsomorphin, an inhibitor of SMAD signaling (Yu et al., 2008), was examined. BMP-2 treatment resulted in phosphorylation of Smad1/5, with that activation inhibited by Dorsomorphin treatment (Fig. 3A). Furthermore, induction of Kcp expression by BMP-2 was suppressed by Dorsomorphin (Fig. 3B). BMP-2 is known to activate the canonical BMP-SMAD signaling pathway as well as the BMP-MAPK signaling pathway (Chen et al., 2012). To confirm whether MAPKs such as ERK1/2, JNK, and p38 MAPK are involved in BMP-2-induced Kcp expression, the level of Kcp expression with BMP-2 treatment in the presence of MAPK inhibitors was examined. Following pretreatment of MC3T3-E1 cells with 10 μM of U0126 (MEK inhibitor), SP600125 (JNK inhibitor), and SB203580 (p38 MAPK inhibitor), treatment with 300 ng/ml of BMP-2 was performed. Surprisingly, the MAPK inhibitors did not inhibit up-regulation of Kcp expression induced by BMP-2, but rather an increase in Kcp expression was observed (Suppl. Fig. 2). These results suggest that induction of Kcp expression is mediated through SMAD signaling and not MAPK signaling.Fig. 3MC3T3-E1 cells were treated with 300 ng/ml of BMP-2 and/or 500 nM of Dorsomorphin (Dor) for 24 h. (A) Dorsomorphin blocked BMP-2-induced phosphorylation of Smad1/5. Using a control phosphorylated Smad 1/5/Actin value of 1, the expression level of phosphorylated Smad 1/5 was calculated as the relative ratio of Smad 1/5/Actin when treated with BMP-2 and Dorsomorphin. (B) The expression level of Kcp when treated with Dorsomorphin was examined. Using a control Kcp/Gapdh value of 1, the expression level of Kcp was calculated as the relative ratio of Kcp/Gapdh when treated with BMP-2 and Dorsomorphin. **p < 0.01 (Student's t-test).Fig. 3
Knockdown of Kcp expression involved in promotion of osteoblast differentiation and mineralization by BMP-2
3.3
Finally, to investigate how induction of Kcp expression by BMP-2 in osteoblasts affects osteoblast differentiation induced by BMP-2, the effects of BMP-2 on osteoblast differentiation when Kcp expression was suppressed were examined. si Kcp treatment suppressed Kcp expression by approximately 70 % (Fig. 4A). Analyses of ALP staining to determine activity showed that BMP-2-induced osteoblast differentiation was reduced by suppression of Kcp expression (Fig. 4B). In addition, mineralization in osteoblasts treated with BMP-2 was suppressed by si Kcp (Fig. 4C), while expression of Alp (Tnap) was also suppressed by si Kcp, whereas expressions of transcription factors for osteoblast differentiation were not affected (Fig. 4D). Based on these results, it is considered possible that induction of Kcp expression is involved, at least in part, in promotion of osteoblast differentiation and mineralization by BMP-2.Fig. 4(A) The expression level of Kcp was examined when si Kcp was applied to MC3T3-E1 cells. *p < 0.05 (Student's t-test) (B) Measurements of ALP staining (left panels) and ALP activity (right panels). MC3T3-E1 cells were treated with 300 ng/ml of BMP-2 and si Kcp for three days, then the effects on osteoblast differentiation were determined. Scale bar shown represents 100 μm. **p < 0.01 (Student's t-test) (C) MC3T3-E1 cells were stained with Alizarin red in the presence or absence of 300 ng/ml of BMP-2 and si Kcp for seven days. Alizarin red dye associated with the cells was dissolved and quantified by measurement of absorbance at 570 nm. Scale bar shown represents 100 μm. **p < 0.01 (Student's t-test) (D) MC3T3-E1 cells were treated with 300 ng/ml of BMP-2 for two days and si Kcp, then expressions levels of osteoblast differentiation marker genes (Tnap, Runx2, Sp7) were examined using quantitative RT-PCR. *p < 0.05, NS: not significant.Fig. 4
Discussion
4
The present results indicate that Kcp provides positive feedback for promotion of osteoblastic differentiation by BMP-2 in osteoblasts. BMP antagonists are induced by BMPs such as Noggin, which is known to block BMP signaling induced by BMP-2 (Gazzerro et al., 1998). Furthermore, BMP-2 has been found to up-regulate the expression of Gremlin 2, a negative regulator of BMP function during osteoblast differentiation (Suzuki et al., 2012). That study also noted regulation of BMP activity by Noggin and Gremlin 2 following a negative feedback system, which may have important roles for protecting bone metabolism from excessive activation by BMP. On the other hand, Nephronectin, thought to promote osteoblast differentiation, has been shown to be induced by BMP-2 in C2C12 cells (Kurosawa et al., 2016). Thus, BMP signaling is considered to be controlled by both positive and negative feedback regulation during the process of osteoblast differentiation.
In the present study, Kcp expression induced by BMP-2 was shown to be suppressed by the SMAD signaling inhibitor Dorsomorphin, thus it is speculated that Kcp induction by BMP-2 is mediated through SMAD signaling. BMP signaling has been shown to activate MAPK in osteoblasts in addition to SMAD-dependent signaling (Weiskirchen et al., 2009), such as previous findings indicating that Osterix, a key transcription factor for osteoblast differentiation, is induced by BMP-2 through p38-mediated MAPK signaling (Celil et al., 2005). However, the present results showed that MAPK activation did not inhibit Kcp expression induced by BMP-2, rather an increase in Kcp expression was observed. A future study that examines regulation of Kcp expression through MAPK signaling is needed.
Physiological functions of Kcp have been shown to inhibit renal fibrotic disease (Lin et al., 2005b), decrease acute and chronic renal injury (Soofi et al., 2013), and suppress aging- and high-fat diet-induced nonalcoholic fatty liver disease (Soofi et al., 2017). Recently, Ye et al. reported that Kcp knockout mice demonstrated cardiac aging (Ye et al., 2023). In the present study, bone morphometric analysis showed no obvious differences in trabecular bone obtained from wild-type and Kcp knockout mice (Suppl. Fig. 3). Additional physicological study findings are needed to clarify whether up-regulation of Kcp expression by BMP-2 is involved in bone metabolism.
Conclusion
5
In the present study, induction of Kcp by BMP-2 was found to provide positive feedback for promotion of osteoblastic differentiation in osteoblasts. Additional research aimed at hard tissue regeneration as well as development of methods for analysis, such as fracture and bone defect repair models, will be conducted in future studies in order to elucidate the physiological mechanism of BMP signal activation by Kcp.
Financial support statement
This work was supported in part by a grant from 10.13039/501100001691JSPS Kakenhi (21K09833).
CRediT authorship contribution statement
Kazuki Toba: Writing – original draft, Formal analysis, Data curation, Conceptualization. Atsushi Yamada: Writing – review & editing, Writing – original draft, Funding acquisition, Formal analysis, Data curation, Conceptualization. Kiyohito Sasa: Formal analysis, Data curation. Tatsuo Shirota: Writing – review & editing, Supervision. Ryutaro Kamijo: Supervision, Conceptualization.
Declaration of competing interest
None of the authors have conflicts of interest to declare regarding the contents of this article.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aizawa R.Yamada A.Seki T.Tanaka J.Nagahama R.Ikehata M.Kato T.Sakashita A.Ogata H.Chikazu D.Maki K.Mishima K.Yamamoto M.Kamijo R.Cdc 42 regulates cranial suture morphogenesis and ossification Biochem. Biophys. Res. Commun.512220191451493085318610.1016/j.bbrc.2019.02.106 · doi ↗ · pubmed ↗
- 2Celil A.B.Hollinger J.O.Campbell P.G.Osx transcriptional regulation is mediated by additional pathways to BMP 2/Smad signaling J. Cell. Biochem.95320055185281578651110.1002/jcb.20429 · doi ↗ · pubmed ↗
- 3Chen G.Deng C.Li Y.P.TGF-beta and BMP signaling in osteoblast differentiation and bone formation Int. J. Biol. Sci.8220122722882229895510.7150/ijbs.2929 PMC 3269610 · doi ↗ · pubmed ↗
- 4Ebendal T.Bengtsson H.Soderstrom S.Bone morphogenetic proteins and their receptors: potential functions in the brain J. Neurosci. Res.5121998139146946956710.1002/(SICI)1097-4547(19980115)51:2<139::AID-JNR 2>3.0.CO;2-E · doi ↗ · pubmed ↗
- 5Gazzerro E.Gangji V.Canalis E.Bone morphogenetic proteins induce the expression of noggin, which limits their activity in cultured rat osteoblasts J. Clin. Invest.10212199821062114985404610.1172/JCI 3459 PMC 509165 · doi ↗ · pubmed ↗
- 6Gomez-Puerto M.C.Iyengar P.V.Garcia de Vinuesa A.Ten Dijke P.Sanchez-Duffhues G.Bone morphogenetic protein receptor signal transduction in human disease J. Pathol.247120199203024625110.1002/path.5170 PMC 6587955 · doi ↗ · pubmed ↗
- 7Ikeya M.Kawada M.Kiyonari H.Sasai N.Nakao K.Furuta Y.Sasai Y.Essential pro-bmp roles of crossveinless 2 in mouse organogenesis Development 133222006446344731703528910.1242/dev.02647 · doi ↗ · pubmed ↗
- 8Kinoshita M.Yamada A.Sasa K.Ikezaki K.Shirota T.Kamijo R.Phorbol-12-myristate 13-acetate inhibits Nephronectin gene expression via protein kinase C alpha and c-Jun/c-Fos transcription factors Sci. Rep.1112021203603464582410.1038/s 41598-021-00034-x PMC 8514542 · doi ↗ · pubmed ↗