Complete genome sequences of Arthrobacter globiformis phages Uzumaki and Argan of cluster AU6
Brandon Mathew, Andrew Sean Lee, Katie Chen, Michael Kaczmarski, Nigel Oommen, Asweel Mehaboob, Vrushali Patel, Yamini Patel, Hannah Saji, Muhammad Ayaan Shamsi, Bryan Gibb

TL;DR
This paper reports the full genome sequences of two phages, Uzumaki and Argan, that infect a soil bacterium from Long Island.
Contribution
The study provides new insights into the genomic and morphological characteristics of phages in cluster AU6.
Findings
Uzumaki and Argan have lambda-like morphology with prolate capsids.
The two phages share 97% gene content similarity.
They are classified in cluster AU6 with related Arthrobacter phages.
Abstract
Bacteriophages Uzumaki and Argan infect Arthrobacter globiformis B-2880 isolated from soil samples in Long Island, New York. These bacteriophages have lambda-like morphology with prolate capsid and share 97% gene content similarity. These traits place them in cluster AU6 with other related Arthrobacter phages.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
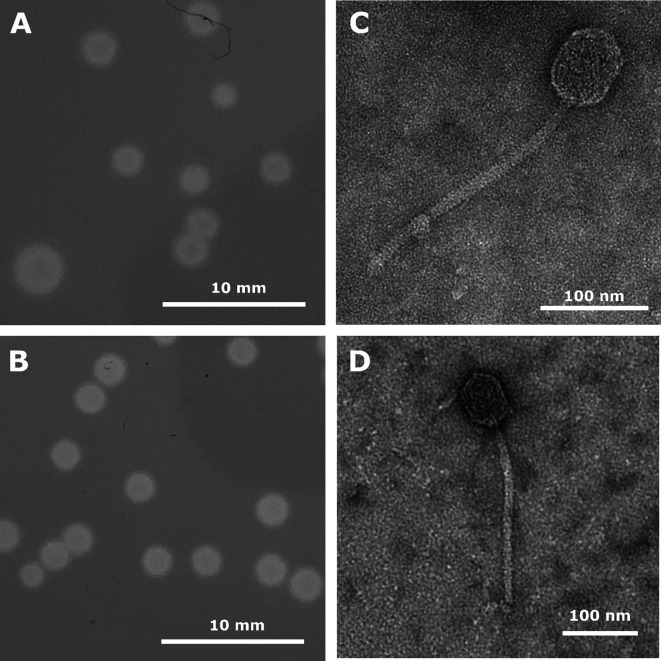 Fig 1
Fig 1| Phage | Genbank accession no. | SRA accession no. | Location (GPS coordinates) | Avg coverage | No. of reads (thousands) | Genome size (bp) | Gc % | Genome end (3' overhang) | No. of genes |
|---|---|---|---|---|---|---|---|---|---|
| Argan |
|
| 40.74181 N, 73.71456 W | 441 | 650.85 | 55,220 | 50.3 | CGCCGGCCT | 92 |
| Uzumaki |
|
| 40.70616 N, 73.70895 W | 1250 | 486.77 | 55,355 | 50.4 | CGCCGGCCT | 92 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Plant and Fungal Interactions Research · Genomics and Phylogenetic Studies
ANNOUNCEMENT
Arthrobacter phages Argan and Uzumaki were isolated from moist soil samples collected in Glen Oaks and Elmont, New York, respectively. Phage isolation, plaque purification, and genome extraction were performed according to the standard procedures described in the SEA-PHAGES Discovery Guide (https://seaphagesphagediscoveryguide.helpdocsonline.com/home), as part of the Science Education Alliance-Phage Hunters Advancing Genomics and Evolutionary Science (SEA-PHAGES) program (1). Both Argan and Uzumaki were isolated by enrichment on Arthrobacter globiformis B-2880 utilizing a peptone–yeast–calcium (PYCa) medium at 30°C for 24–48 h, undergoing three rounds of plaque purification and form clear round plaques 2–6 mm in diameter with defined edges (Fig. 1). High-titer phage lysate was produced by plate lysis using double-agar overlays in phage buffer and 0.22 μm filtered. Negative-stained transmission electron microscopy showed that both phages have prolate icosahedral heads (58–64 nm) and (236–238 nm) long non-contractile tails, consistent with other Caudoviricetes bacteriophage-like phage-lambda (Fig. 1) (2, 3).
Plaque morphology (A and B) and transmission electron microelectron micrographs (C and D) of Arthrobacter phages Uzumaki (A and C) and Argan (B and D). Phage plaques were imaged following incubation at 30°C for 48 h. Phage lysate was negatively stained with 1% uranyl acetate and imaged with a JEOL JEM-1400 transmission electron microscope at 120 KeV.
Genome extraction was performed from high-titer lysates using the Wizard DNA cleanup kit (Promega), and sequencing was performed at the University of Pittsburgh. Libraries were constructed using the NEB Ultra II FS DNA library prep kit and sequenced using an Illumina MiSeq v3 platform with single-end sequencing, resulting in 150 bp reads. Raw reads were assembled using Newbler v2.9 (4) with default settings, generating single contigs with an average read depth of approximately 1,250 for Uzumaki and 441 for Argan. The phage contigs were verified using Consed v29 to evaluate completeness and determine genomic termini (5). The genome parameters (length, GC content, and termini) and accession numbers (GenBank and SRA) are shown in Table 1. The genomes of both phages have defined ends with nine-base complementary 3′ single-stranded extensions.
Both phages were assigned to cluster AU6 containing other related Arthrobacter phages based on the gene content similarity (GCS) greater than 35% using the tool at PhagesDB (6, 7). Coding regions were predicted using GeneMark v3.25 (8) and Glimmer v3.02 (9). The coding regions were manually checked using Starterator v1.2 (http://phages.wustl.edu/starterator/), PECAAN v20211202 (https://blog.kbrinsgd.org/), and DNA Master v5.23.6 (10). Functions for the genes were found using NCBI BLASTp v2.9 (11), HHPred v2.0.13 (12), and Phamerator (13). No tRNA genes were identified by Aragorn v1.2.41 (14) and tRNAscanSe v2.0 (15). The membrane proteins were predicted using DeepTmHmm v2.0 (16) and TOPCONS 2.0 (17). All software was used with default settings.
Uzumaki and Argan are both predicted to encode 92 protein-coding genes, but only 25 and 23 are assigned putative functions. Argan and Uzumaki have 97% nucleotide identity and 85.87% GCS (6, 7). All genes in both genomes are transcribed on the same strand. The genome organization for both phages is similar, with the genes coding for phage structure and assembly located on the left half of the genome, whereas the genes involved in DNA replication and metabolism are found on the right half. No integrase or immuno-repressor genes were identified, suggesting that the phage exhibits an entirely lytic cycle, which is consistent with the clear plaque morphologies observed in each phage.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jordan TC , Burnett SH , Carson S , Caruso SM , Clase K , De Jong RJ , Dennehy JJ , Denver DR , Dunbar D , Elgin SCR , et al. . 2014. A broadly implementable research course in phage discovery and genomics for first-year undergraduate students. m Bio 5:e 01051-13. doi:10.1128/m Bio.01051-13 24496795 PMC 3950523 · doi ↗ · pubmed ↗
- 2Turner D , Shkoporov AN , Lood C , Millard AD , Dutilh BE , Alfenas-Zerbini P , van Zyl LJ , Aziz RK , Oksanen HM , Poranen MM , et al. . 2023. Abolishment of morphology-based taxa and change to binomial species names: 2022 taxonomy update of the ICTV bacterial viruses subcommittee. Arch Virol 168:74. doi:10.1007/s 00705-022-05694-2 36683075 PMC 9868039 · doi ↗ · pubmed ↗
- 3Ackermann HW . 1998. Tailed bacteriophages: the order caudovirales. Adv Virus Res 51:135–201.9891587 10.1016/S 0065-3527(08)60785-XPMC 7173057 · doi ↗ · pubmed ↗
- 4Miller JR , Koren S , Sutton G . 2010. Assembly algorithms for next-generation sequencing data. Genomics 95:315–327. doi:10.1016/j.ygeno.2010.03.001 20211242 PMC 2874646 · doi ↗ · pubmed ↗
- 5Gordon D , Green P . 2013. Consed: a graphical editor for next-generation sequencing. Bioinformatics 29:2936–2937. doi:10.1093/bioinformatics/btt 515 23995391 PMC 3810858 · doi ↗ · pubmed ↗
- 6Russell DA , Hatfull GF . 2017. Phages DB: the actinobacteriophage database. Bioinformatics 33:784–786. doi:10.1093/bioinformatics/btw 711 28365761 PMC 5860397 · doi ↗ · pubmed ↗
- 7Pope WH , Mavrich TN , Garlena RA , Guerrero-Bustamante CA , Jacobs-Sera D , Montgomery MT , Russell DA , Warner MH , Hatfull GF , Science Education Alliance-Phage Hunters Advancing Genomics and Evolutionary Science (SEA-PHAGES) . 2017. Bacteriophages of Gordonia spp. display a spectrum of diversity and genetic relationships. m Bio 8:e 01069-17. doi:10.1128/m Bio.01069-17 28811342 PMC 5559632 · doi ↗ · pubmed ↗
- 8Besemer J , Lomsadze A , Borodovsky M . 2001. Gene Mark S: a self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 29:2607–2618. doi:10.1093/nar/29.12.2607 11410670 PMC 55746 · doi ↗ · pubmed ↗