Complete genome sequence of Microbacterium foliorum phage Curie, a podovirus isolated from soil in Spokane, Washington
Emma N. Horton, Erika K. Beach, Kathryn T. Cook, Kyra G. Cronin, Avery T. Haag, Sierra M. Salter, Nicole A. Stojanovic, Zoe E. Fry, Brian M. Connolly, Rebekah F. Hare, Ann-Scott H. Ettinger, Marianne K. Poxleitner, Kirk R. Anders

TL;DR
This paper presents the full genome sequence of a soil-isolated bacteriophage called Curie, which infects Microbacterium foliorum.
Contribution
The discovery of a new actinobacteriophage Cluster GI formed by Curie and PineapplePizza.
Findings
The genome of phage Curie is 16,810 bp long with 90 bp terminal inverted repeats.
Curie's genome architecture is similar to PineapplePizza and other phi29-like phages.
Curie and PineapplePizza define a new actinobacteriophage Cluster GI.
Abstract
Bacteriophage Curie is a podovirus that infects Microbacterium foliorum. The Curie genome spans 16,810 bp, has 90 bp terminal inverted repeats, and includes 23 protein-coding genes. Its genome architecture resembles phage PineapplePizza and other phi29-like phages. Together, Curie and PineapplePizza form a new actinobacteriophage Cluster GI.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
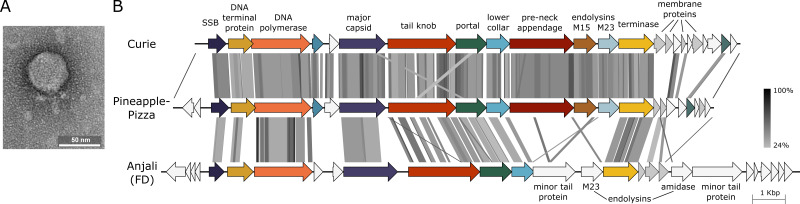 Fig 1
Fig 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Plant Virus Research Studies · Genomics and Phylogenetic Studies
ANNOUNCEMENT
Studying the genomic diversity of bacteriophages provides insight into their biology and evolution (1 – 3), and may also contribute to development of therapies against antibiotic-resistant bacterial infections (4). To explore the diversity of phages that can infect actinobacteria, we isolated phage Curie on Microbacterium foliorum. Here, we describe its genome.
Fifteen cubic centimeters of top layer soil was collected in Spokane, WA (47.66669 N, 117.40284 W; September 2021), incubated 7 days in 35 mL PYCa medium (5) seeded with 0.5 mL of a saturated, 2-day batch culture of Microbacterium foliorum NRRL B-24224, then filtered (0.22 µm). Plaques were found on lawns grown from 0.5 mL saturated M. foliorum culture and 10 µL phage solution in PYCa top agar on a PYCa base and incubated 2–7 days at 22°C (6). An isolated plaque was picked and replated three times, and a lysate was collected (6). Transmission electron microscopy revealed a podoviral morphology with average capsid width of 46 nm and tail length of 27 nm (Fig. 1A, n = 3).
Phage Curie virion and genome structure. (A) Transmission electron micrograph of Curie, negatively stained with 1% uranyl acetate. (B) Curie genome compared to PineapplePizza and Anjali, a representative of Cluster FD. Gray bands between genomes show tBLASTx (v.2.15) alignments (7). The shade of each band indicates percent identity according to the grayscale bar, 24%–100%. Genes (arrows and arrowheads) are filled with the same color if they were members of gene families formed by Phamerator (8), using sequences in the Actino_Draft database version 552 (9). Genes filled with gray encode proteins with at least one predicted transmembrane domain. Endolysins are labeled with the domain they contain: M15 peptidase, M23 peptidase, or N-acetylmuramoyl-L-alanine amidase. The genome alignment was generated with EasyFig v.2.2, filtering for tBLASTx e-values smaller than 10−50 (10).
DNA was extracted from the lysate with the Promega DNA Wizard Clean-Up System. A library was made with the NEB Ultra II FS kit (v.3 reagents) and sequenced with Illumina MiSeq, producing 290,051 single-end, 150-base reads that were assembled with Newbler v.2.9 (11). Consed v.29 (12) was used to assess and finish the assembly, which had an average coverage depth of 2,141×, resulting in a genome of 16,810 bp with terminal inverted repeats of 90 bp. The G + C content was 50.1%, distinct from M. foliorum DNA at 68.7% (13). As no sequencing reads extended beyond the ends, a terminal protein may be covalently attached, similar to phi29 (14). Curie was most closely related to PineapplePizza (GenBank ON724010), sharing an average nucleotide identity of 63% (DNA Master v.5.23, http://cobamide2.bio.pitt.edu) (Fig. 1B). Curie joined PineapplePizza to form actinobacteriophage Cluster GI, based on nucleotide sequence similarity and shared gene content (8, 9, 15, 16).
Twenty-three protein-coding genes were predicted using GeneMark v.2.5p (17), Glimmer v.3.02 (18), Starterator v.1.2 (https://github.com/cdshaffer/starterator), and Phamerator (8). No tRNA genes were detected with Aragorn v.1.2 (19) or tRNAscan-SE v.2.0 (20). Gene functions were predicted using HHPred alignments in PDB, Pfam, SCOP, and CDD databases (21 – 25), NCBI BLAST v.2.14 (7), the Actinobacteriophage Database (9), and DeepTMHMM v.1.0 (https://dtu.biolib.com/DeepTMHMM). (All software used default parameters, unless noted.) Curie, PineapplePizza, and phages in Cluster FD such as Anjali (MK016490) share a similar core genome architecture (Fig. 1B). The shared region extends from the ssDNA-binding protein gene on the left to terminase on the right, although Anjali and its relatives (isolated on Arthrobacter globiformis) carry different minor tail and endolysin genes (Fig. 1B). The shared genes are similar to those of the podovirus phi29 (26), whose virion structure is well-described (27). Curie and the other phi29-like genomes contain 10–13 small genes near the genome ends, several of which may encode transmembrane domains (Fig. 1B). Although we detected no sequence similarity with phi29 holin, we speculate that one or more of the membrane proteins functions as a holin.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dion MB , Oechslin F , Moineau S . 2020. Phage diversity, genomics and phylogeny. Nat Rev Microbiol 18:125–138. doi:10.1038/s 41579-019-0311-5 32015529 · doi ↗ · pubmed ↗
- 2Hatfull GF . 2020. Actinobacteriophages: genomics, dynamics, and applications. Annu Rev Virol 7:37–61. doi:10.1146/annurev-virology-122019-070009 32991269 PMC 8010332 · doi ↗ · pubmed ↗
- 3Jansson JK , Wu R . 2023. Soil viral diversity, ecology and climate change. Nat Rev Microbiol 21:296–311. doi:10.1038/s 41579-022-00811-z 36352025 · doi ↗ · pubmed ↗
- 4Hatfull GF , Dedrick RM , Schooley RT . 2022. Phage therapy for antibiotic-resistant bacterial infections. Annu Rev Med 73:197–211. doi:10.1146/annurev-med-080219-122208 34428079 · doi ↗ · pubmed ↗
- 5Petrovski S , Seviour RJ , Tillett D . 2011. Genome sequence and characterization of the Tsukamurella bacteriophage TPA 2. Appl Environ Microbiol 77:1389–1398. doi:10.1128/AEM.01938-10 21183635 PMC 3067230 · doi ↗ · pubmed ↗
- 6Poxleitner M , Pope W , Jacobs-Sera D , Sivanathan V , Hatfull G . 2018. Phage Discovery Guide. Howard Hughes Medical Institute, Chevy Chase, MD. Available from: https://seaphagesphagediscoveryguide.helpdocsonline.com/home
- 7Altschul SF , Gish W , Miller W , Myers EW , Lipman DJ . 1990. Basic local alignment search tool. J Mol Biol 215:403–410. doi:10.1016/S 0022-2836(05)80360-2 2231712 · doi ↗ · pubmed ↗
- 8Cresawn SG , Bogel M , Day N , Jacobs-Sera D , Hendrix RW , Hatfull GF . 2011. Phamerator: a bioinformatic tool for comparative bacteriophage genomics. BMC Bioinformatics 12:395. doi:10.1186/1471-2105-12-395 21991981 PMC 3233612 · doi ↗ · pubmed ↗