Exosomal Preconditioning of Human iPSC-Derived Cardiomyocytes Beneficially Alters Cardiac Electrophysiology and Micro RNA Expression
Øystein Røsand, Jianxiang Wang, Nathan Scrimgeour, Gurdeep Marwarha, Morten Andre Høydal

TL;DR
This study shows that exosomes from human stem cell-derived heart cells can improve heart cell function under stress, potentially aiding in treating heart disease.
Contribution
The novel finding is that exosome preconditioning alters cardiac electrophysiology and miRNA expression in hypoxic conditions.
Findings
Exosome preconditioning increased beat period by 15.7% in hypoxic hIPSC-CMs.
Preconditioning improved excitation-contraction coupling by 25.3% under hypoxia.
Ten miRs and 44 gene targets were identified, linked to cell survival and ion channel modulation.
Abstract
Experimental evidence, both in vitro and in vivo, has indicated cardioprotective effects of extracellular vesicles (EVs) derived from various cell types, including induced pluripotent stem cell-derived cardiomyocytes. The biological effects of EV secretion, particularly in the context of ischemia and cardiac electrophysiology, remain to be fully explored. Therefore, the goal of this study was to unveil the effects of exosome (EXO)-mediated cell–cell signaling during hypoxia by employing a simulated preconditioning approach on human-induced pluripotent stem cell-derived cardiomyocytes (hIPSC-CMs). Electrophysiological activity of hIPSC-CMs was measured using a multielectrode array (MEA) system. A total of 16 h of hypoxic stress drastically increased the beat period. Moreover, hIPSC-CMs preconditioned with EXOs displayed significantly longer beat periods compared with non-treated cells…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
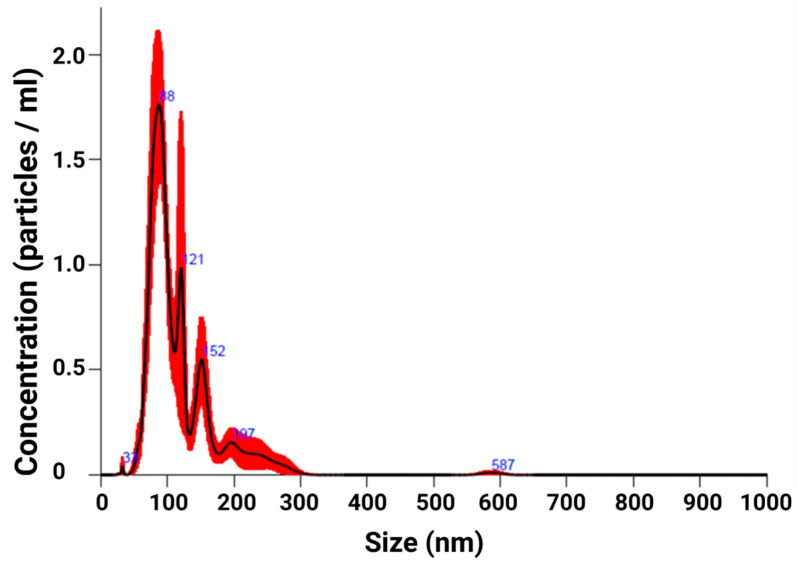 Figure 1
Figure 1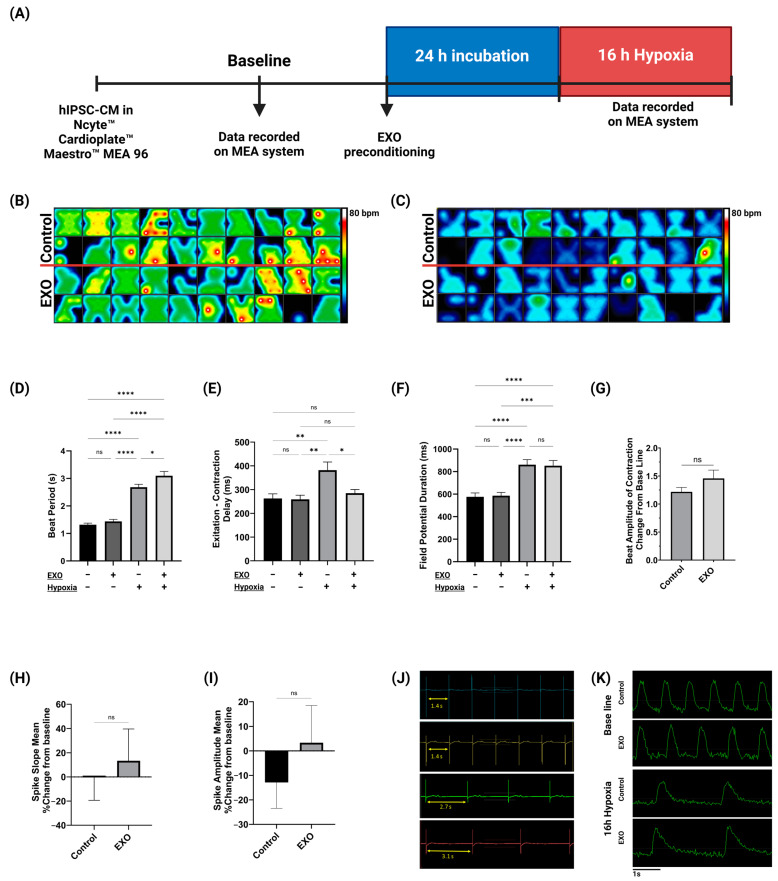 Figure 2
Figure 2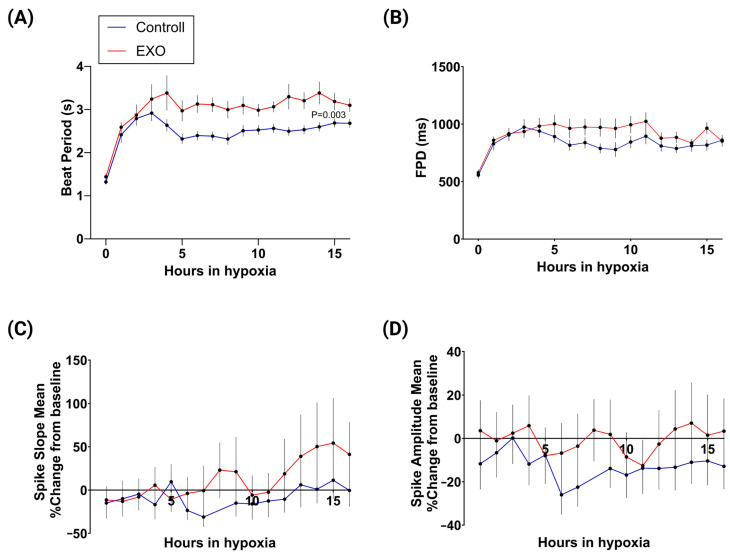 Figure 3
Figure 3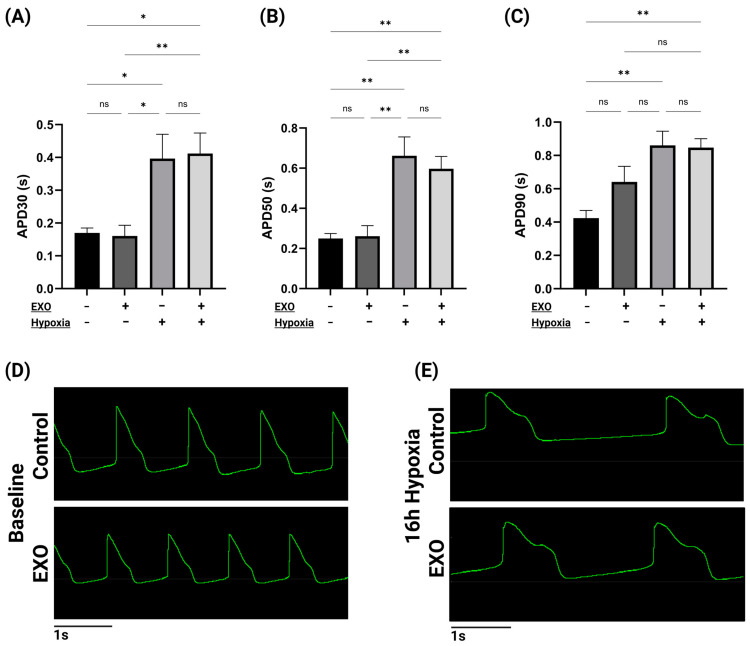 Figure 4
Figure 4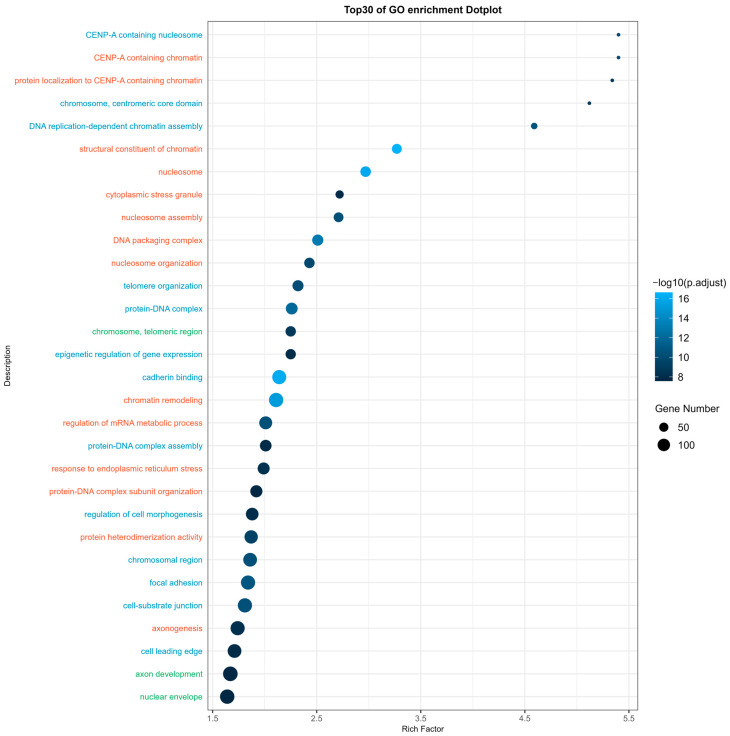 Figure 5
Figure 5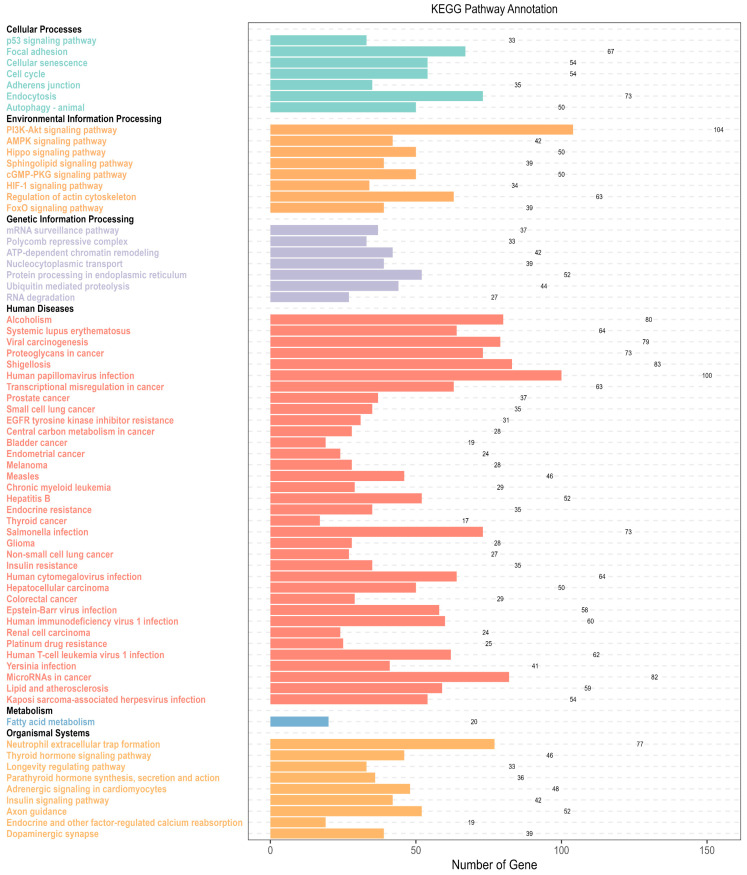 Figure 6
Figure 6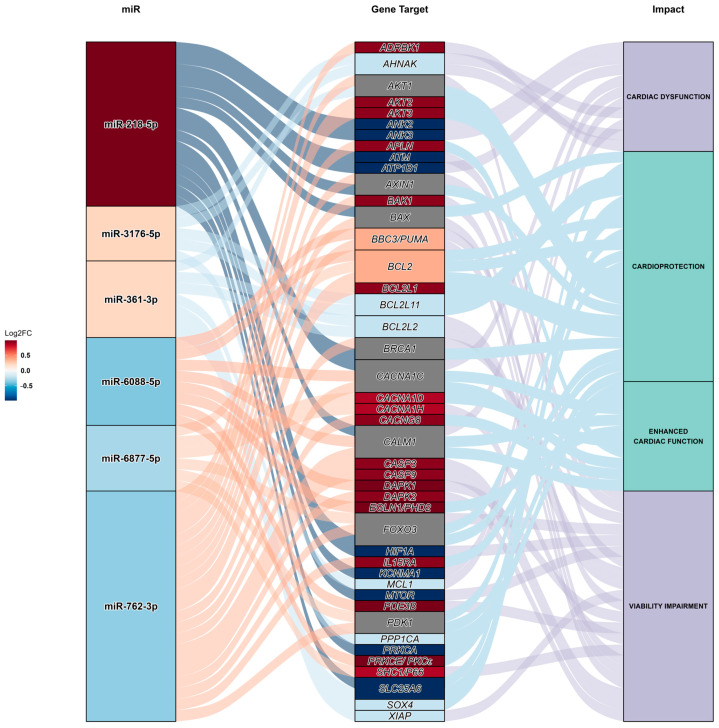 Figure 7
Figure 7- —The Liaison Committee between the Central Norway Regional Health Authority (RHA) and the Norwegian University of Science and Technology (NTNU) and NTNU health
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsExtracellular vesicles in disease · Planarian Biology and Electrostimulation · Pluripotent Stem Cells Research
1. Introduction
Despite significant research efforts, ischemic heart disease (IHD) is still the leading cause of mortality worldwide [1]. IHD is primarily caused by partial or total disruption of blood flow to the myocardium, resulting in reduced delivery of oxygen and nutrients to the cardiac cells. The hypoxic stress elicited by IHD induces metabolic stress, e.g., metabolic acidosis, accumulation of reactive oxygen species (ROS), and Ca^2+^ overload, which, if not resolved, results in cardiomyocyte apoptosis and/or necrosis, and ultimately, irreversible damage of the heart. If the cardiac damage exceeds a critical limit, patients will suffer from acute myocardial infarction (AMI) [2,3,4,5]. AMI is strongly associated with dysregulation of cardiac function and alterations in electrophysiological properties. Propper cardiac function relies on a tight regulation of cardiac depolarization, contraction and relaxation, and finally repolarization [6]. This is facilitated by various molecules in cardiomyocytes, e.g., ion channels, transporters, and intracellular Ca^2+^-handling proteins [7]. Irregular cardiac electrophysiology leads to cardiac arrhythmias that can negatively impact the myocardium’s ability to pump blood [7,8].
In cardiac physiology and pathology, there has been an increased interest in researching the intracellular communication facilitated by EVs. Exosome (EXO) is the most frequently used term to describe a subgroup of EVs characterized by their 30–150 nm diameter and biogenetic origin [9,10,11]. No longer regarded as just a means for the cells to discard waste, EXOs are now understood to facilitate intercellular communication by transporting signaling molecules between donor and target cells [11]. Internalized proteins, lipids, coding and non-coding RNA, DNA, and metabolites play essential roles in intercellular communication, changing the fate of recipient cells. Among these internalized elements, microRNA (miR) are emerging as important regulators of myocardial biology and disease, as evidenced by multiple studies [12,13]. miRs can regulate gene expression through transcriptional modulation, epigenetic targeting, or post-transcriptional regulation [14,15,16,17]. It is already well established that ischemia induces profound changes in miR expression, which is important for myocardial ischemia-related genes [18]. However, new evidence has demonstrated that miRs also can regulate the electrophysiology of the heart by either conventionally affecting the gene expression of ion channels, transporters, intracellular Ca^2+^-handling proteins, and other relevant factors [19,20,21,22,23], or by direct biophysical modulation of ion channels [24].
During the last decade, considerable resources have been used to develop cardioprotective strategies based on EXOs released from human-induced pluripotent stem cell-derived cardiomyocytes (hIPSC-CMs). Both in vitro and in vivo studies have shown that treatment with EVs secreted from IPSCs enhance angiogenesis, enhance cell proliferation, reduce cardiomyocyte apoptosis, and improve left ventricular function after myocardial infarction [25,26,27]. Of note, ischemic preconditioning is once such therapeutic strategy, where EXOs isolated from ischemic cardiomyocytes, once administrated, can elicit cardioprotective effects in recipient cardiomyocytes under ischemic conditions [28,29]. However, despite these promising data, the paracrine EXO signaling between cardiomyocytes of the same myocardium, as well as EXO signaling during ischemia, remains poorly understood. Improving our knowledge of cell–cell signaling within the failing heart may give rise to therapeutic solutions in the future. Therefore, this study aimed to investigate whether EXOs released by hIPSC-CMs during hypoxia can positively influence cardiac electrophysiology and miR expression during hypoxic stress. A multielectrode array (MEA) system was used as an in vitro method of recording real-time cardiac electrophysiological activity, highlighting the biological effects facilitated by hypoxic EXOs. Additionally, miR-sequencing was performed to compare miR expressions in EXO-preconditioned and non-preconditioned hIPSC-CMs, shedding light on important modulators of cardiac electrophysiology. Lastly, an miR gene target prediction analysis was performed, bridging the MEA and miR-sequencing analysis, emphasizing possible therapeutic gene targets for future studies. Thus, this study offers novel insights into the mechanisms of intercellular communication, facilitated by EXOs, during hypoxic stress across cardiomyocytes of the same group.
2. Results
2.1. EXO Isolation
Nanoparticle tracing analysis (NTA) of isolated EXOs (Figure 1) revealed a mean particle size of 167.4 nm (±25.6 nm). Furthermore, NTA measured EXO concentrations of 9.2 × 10^7^ EXOs/mL (±2.0 × 10^7^ EXOs/mL). Our NTA analysis detected particles predominately in the 30–200 nm size range, which is consistent with previous reports of EXO size range [30]. Few to no larger particles were detected.
2.2. Electrophysiological Activity of hIPSC-CMs
MEA data were analyzed at baseline and after 16 h of hypoxia (Figure 2A–C). Here, we report that 16 h of hypoxia drastically increased the beat period of both cell groups. Interestingly, after 16 h of hypoxia, we found that EXO-preconditioning of hIPSC-CMs increased the beat period significantly compared with the control group (+15.7%, p < 0.05) (Figure 2D). In addition, hypoxia significantly impacted hIPSC-CM excitation–contraction (EC)-coupling. We found that 16 h of hypoxia prolonged EC-coupling for the control group (+45.3%, p < 0.05), while hIPSC-CMs preconditioned with hypoxic EXOs displayed significantly faster EC-coupling compared to non-preconditioned CMs after 16 h of hypoxia (−25.3%, p < 0.05). The EC-coupling of EXO-preconditioned cells was comparable to the EC-coupling of cells at baseline normoxic recordings (Figure 2E). Our results also show that 16 h of hypoxia had a profound impact on the field potential duration (FPD) of hIPSC-CMs. We did, however, not find any difference in the FPD between the control and EXO-preconditioned hIPSC-CMs after hypoxia (Figure 2F). hIPSC-CM beat amplitude of contraction, measured by array-based impedance, was not significantly altered in EXO-preconditioned cells compared to non-treated cells (Figure 2G). Moreover, there were no significant changes in the spike slope (Figure 2H), nor the spike amplitude (Figure 2I) of the FPD, between the two groups after hypoxia. Data from the MEA recordings illustrate the beat periods (Figure 2J) and contractions (Figure 2K) for the hIPSC-CMs at baseline and after 16 h of hypoxia for both experimental groups.
Hourly MEA recordings over the 16 h hypoxia period revealed dynamic changes in the electrophysiological activity of hIPSC-CMs under hypoxic stress (Figure 3). Our results show that the beat period for preconditioned cells was significantly longer than the non-treated cells throughout the majority of the hypoxia period (Figure 3A, p = 0.003). Moreover, both the beat period and the FPD showed a swift increase during the initial hours of hypoxia (1–3 h), followed by a period of stabilization. Overall, there were no significant changes in FPD between preconditioned and non-preconditioned hIPSC-CMs (Figure 3B). However, the FPD of preconditioned cells was significantly increased at 8 h of hypoxia (+31%, p < 0.05) compared to non-preconditioned cells. We did not observe any difference in FPD spike slope (Figure 3C) or FPD spike amplitude (Figure 3D) throughout the 16 h of hypoxia.
Local extracellular action potential (LEAP) recordings (Figure 4) indicate that both APD30 and APD50 were significantly increased for both cell groups after 16 h of hypoxia (Figure 4A,B). Interestingly, APD90 was only significantly changed in non-treated cells after hypoxia (Figure 4C). Furthermore, EXO preconditioning did not significantly affect the APD30, APD50, or APD90 of hIPSC-CMs compared to the control group after 16 h of hypoxia. Data from the MEA recordings illustrate the LEAP for the hIPSC-CMs at baseline (Figure 4D) and after 16 h of hypoxia (Figure 4E) for both experimental groups.
2.3. miR Sequencing of Non-Treated and Pre-Conditioned hIPSC-CMs
In our study, small RNA-seq identified 1027 unique miRs in the hIPSC-CM (Table S2). Out of the detected miRs, 10 were differently expressed between the preconditioned and non-preconditioned cells. Compared with the control group, six miRs were downregulated and four miRs were upregulated in the EXO-preconditioned hIPSC-CMs (Table 1). A p-adjusted value of ≤0.2 was selected to ensure that only statistically significant miR regulations were included in the following target prediction analysis.
2.4. miR Target Prediction Analysis
Here, the Gene Ontology (GO) enrichment analysis (Figure 5) revealed that the top 30 enriched GO terms correlated with significantly expressed miRs identified in the study. These terms fell into three principal domains: biological process (orange), cellular component (blue), and molecular function (green). The significance of each term was determined by its p-adjusted value and Rich factor, underscoring the relevance and robustness of the enrichment within the dataset. The Rich factor reflects the ratio of input genes annotated in a term compared to all genes similarly annotated, whereby a higher Rich factor suggests a more substantial enrichment. In summary, the enriched terms predominantly pertain to the umbrella concept of genetic and epigenetic information processing, cell protein interaction, macromolecule aggregation, cell morphological changes, and cell stress response—notably, the response to endoplasmic reticulum stress.
The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway annotation (Figure 6) demonstrates the significant enrichment of various cell pathways based on the KEGG database. The enriched terms fall within the BRITE hierarchy parent categories: cellular processes, environmental information processing, genetic information processing, human diseases, metabolism, and organismal systems. The interrelations among the KEGG pathways mirror the effects of the enriched terms observed in GO analysis across various aspects.
These initial analyses yielded more than 900 predicted target genes for the miRs in question (Table S3). Following the comprehensive screening process, all predicted target genes from hsa-miR-1234-3p, hsa-miR-1293, and hsa-miR-9718 were excluded due to irrelevant target function and/or no recorded tissue expression in adult CMs. Finally, 44 predicted genes targeted were identified (Table 2).
A Sankey diagram (Figure 7) was generated as a means to illustrate the interactions of all miRs and the 44 predicted target genes. The Sankey diagram is organized into three columns representing up- and downregulated miRs, gene targets, and the impact of gene regulation. The diagram features interconnected flow paths that link different strata of the columns. The Sankey diagram shows that the upregulated mir-218-5p and downregulated mir-762-3p were predicted to target the highest number of genes. Based on the middle column of the diagram, 20 of the predicted targeted genes were predicted to be upregulated (red), while 16 were predicted to be downregulated (blue). The remaining eight predicted gene targets were predicted to be targeted by both up- and downregulated miRs. Their expression could therefore not be predicted. Furthermore, there was an even spread between possible beneficial and detrimental effects, based on the predicted targeted genes and their reported functions in CMs. Cardioprotection and viability impairment represent the largest parts of the impact column.
3. Discussion
Recent advances within the cardiovascular field have highlighted the potential of circulating EVs for cardioprotection. Studies have demonstrated that EVs can aid in cardiac remodeling, the enhancement of cardiac function, and the attenuation of cardiomyocyte cell death following myocardial infarction [25,26,27]. However, the biological implications of EV secretion and its effects on recipient cardiomyocytes, particularly in paracrine signaling, modulation of cardiac electrophysiological function, and miR expression during ischemic events, remain largely unexplored. The present study corroborates the profound impact of hypoxia on cardiomyocyte functionality and electrophysiological parameters. Moreover, our results show that EXOs secreted during 16 h of hypoxia beneficially alter the electrophysiological properties of hIPSC-CM, prolonging the cardiac beat period and improving EC-coupling during hypoxic stress. Next-generation sequencing identified 10 miRs differentially regulated in preconditioned cells compared to controls. Gene target prediction analysis revealed that these miRs are associated with stress response, genetic and epigenetic information processing, cell morphological changes, and cell survival. Furthermore, a total of 44 relevant gene targets were predicted to be impacted by the up- and downregulated miRs.
Our MEA data show that 16 h of hypoxia drastically increased the beat period compared to normoxic conditions. hIPSC-CM preconditioned with EXOs displayed a significantly longer beat period compared with non-treated cells after hypoxia, possibly due to the downregulation of the cyclic nucleotide gated 4 (HCN4) channel, known to be linked to slow heart rate [74]. A decrease in heart rate can be beneficial for CMs, as it decreases myocardial oxygen consumption and reduces the workload on the CMs, which may be advantageous in the context of hypoxia [75].
We also demonstrated that the hIPSC-CMs preconditioned with EXOs exhibited faster EC-coupling compared to non-treated cells, indicating more efficient translation of electrical signals (excitation) into mechanical contractions of the actin–myosin filaments (contraction) [76]. Cardiomyocyte contraction is regulated by complex pathways, including Ca^2+^ handling and troponin activation [77]. Improved EC-coupling without affecting the beat amplitude of contraction suggests that EXO-preconditioning influenced the rate, but not the amount, of Ca^2+^ release from the sarcoplasmic reticulum (SR). Multiple predicted gene targets of our differentially expressed miRs were predicted to impact intracellular Ca^2+^ handling. Dai et al. [37] showed that apelin (APLN) increased contractility in failing rat cardiac muscle by increasing Ca^2+^ availability. We found APLN to be a target of the downregulated mir-762-3p. Furthermore, ion channels, key modulators of Ca^2+^ regulation in cardiomyocytes, can significantly affect cardiac function. The L-type Ca^2+^ channel (LTCC) is crucial for the influx of Ca^2+^ across the sarcolemma. We predicted that several of the differentially expressed miRs target one or more subunits of the LTCC, such as the downregulated mir-762-3p, which was predicted to affect the calcium voltage-gated channel auxiliary subunit gamma 8 (CACNG8). CACNG8 has been shown to encode a subunit of LTCC that is involved in regulating LTCC gating properties [51].
Despite great changes in FPD after 16 h of hypoxia, no difference in FPD, spike slope, or spike amplitude was observed between the preconditioned cells and the control cells. We found that hypoxic stress increased APD30, APD50, and APD90 in both groups. However, EXO preconditioning did not have any effect on these parameters, suggesting that cardiomyocyte membrane re-polarization was not affected by the EXO treatment. Additionally, EXO preconditioning did not significantly alter hIPSC-CM contraction amplitude after hypoxia, suggest that EXO preconditioning affected the beat rate and EC-coupling of hIPSC-CMs without greatly altering the cardiomyocyte contraction, excitability, or action potential duration of the cells.
The mitochondria are vital for intracellular Ca^2+^ dynamics via Ca^2+^ exchange, ATP generation, and ROS production. Cardiomyocytes utilize ATP to activate the contractile machinery, such as the calcium pumps (SERCA2a and the plasma membrane Ca^2+^ ATPase pump (PMCA)). ATP availability is therefore crucial for Ca^2+^ handling and cardiac function [78,79]. Furthermore, during myocardial ischemia, reducing levels of oxidative stress and inflammation helps to alleviate cellular damage. Many of our predicted gene targets were implicated in impacting ROS regulation and mitochondrial function, such as the downregulated miR-6088-5p. miR-6088-5p, which was predicted to target protein kinase C epsilon (PRKCE), has been shown to modulate mitochondrial function. Furthermore, upregulation of PRKCE has been shown to improve glucose utilization, maintain ATP production, and reduce cardiac pathophysiologic responses during chronic hypoxia [69]. Downregulation of mir-6088-5p might therefore aid the mitochondria in providing sufficient ATP to facilitate improved intracellular Ca^2+^ handling in preconditioned hIPSC-CMs [80].
Many of the miRs differentially regulated in the EXO-preconditioned hIPSC-CMs have directly been associated with cardiac disease, i.e., miR-218-5p, miR-361-3p, miR-6088-5p, and miR-762-3p. An in vitro study conducted by Sun et al. [81] found that miR-218-5p expression decreased after myocardial infarction (MI). Furthermore, they showed that miR-218-5p mediated myocardial fibrosis by inhibition of transforming growth factor beta 1 (TGF-β1). Furthermore, a loss-of-function study performed by Zhang et al. [82] showed that downregulation of miR-361-3p further progressed hypoxia-induced cell injury in H9c2 cells. Downregulated miR miR-6088-5p has previously been shown to be a possible biomarker for hypertrophic cardiomyopathy (HCM), as it was found to be downregulated in HCM patients [83]. In 2019, Yan et al. [84] published a study showing that miR-762-3p translocated in to the mitochondria and was significantly upregulated following anoxia/reoxygenation treatment. Furthermore, they also showed that myocardial ischemia/reperfusion injury was reduced by knockdown of miR-762-3p.
Accurate characterization of EXO is an important aspect of EV based research. A limitation of this study is that the characterization of the hIPSC-CM derived EXOs primarily relied on NTA. While NTA provides valuable information on particle size and concentration, it cannot reliably distinguish EXOs from other similarly sized particles, such as high-density lipoproteins (HDL). Implementing the detection of known EXO markers, such as heat shock proteins (HSP70 and HSP90), tetraspanins (CD9, CD81), or endosomal markers (TSG101), via western blotting, would enhance the robustness of the EXO characterization process [85]. However, the EXO yield from the hIPSC-CM was insufficient to meet the high sample input requirements for such analysis. Our findings should therefore be interpreted in light of this limitation.
4. Materials and Methods
4.1. hIPSC-CMs for EXO Production
For EXO production, an Ncyte™ Cardioplate™ Maestro™ MEA 96 (Ncardia, # Nc-K-NCP-M96, Leiden, The Netherlands) with pre-seeded Ncyte^TM^ hIPSC-CMs (10^4^ cells/well) was acquired and maintained following the manufacturer’s instructions. Once received, the hIPSC-CMs were maintained in Cardiomyocyte Culture Media (# Nc-M-CMCM-250, Ncardia, Leiden, The Netherlands) at 21% O_2_, 5% CO_2_ in a humidified atmosphere at 37 °C. Once settled following shipping, the cells were thoroughly washed with sterile phosphate-buffered saline (PBS) (1X) to remove any trace of the culture media before the cells were exposed to hypoxic stress: 16 h incubation with hypoxia medium (Table S1) at 1% O_2_, 5% CO_2_, 94% N_2_ in a humidified atmosphere at 37 °C [86,87]. Following 16 h of hypoxia, EXOs were isolated from the conditioned medium of the hypoxia-stressed hIPSC-CMs. hIPSC-CMs were used within 14 days of arrival, as recommended by the manufacturer.
4.2. EXO Isolation and Resuspension
EXO isolation was performed using Total Exosome Isolation Reagent (from the cell culture medium) (Invitrogen^TM^, #4478359, Waltham, MA, USA), following the manufacturer’s instructions. Initially, hypoxia-conditioned medium was aspirated from a 96-well MEA plate and pooled before being centrifuged at 2000× g for 30 min, removing cells and debris. The supernatant was then transferred to a new collection tube before a 0.5 volume of the Total Exosome Isolation Reagent (from the cell culture medium) was added. The supernatant and isolation reagent were mixed well before incubation at 2 °C to 8 °C overnight. The following day, the samples were centrifuged at 10,000× g for 1 h at 2 °C to 8 °C. Then, the supernatant was aspirated and discarded. Lastly, the EXO pellet was resuspended in Cardiomyocyte Culture Media. EXOs were administered as a preconditional treatment directly after isolation/resuspension.
4.3. EXO Quantification and Characterization
Nanoparticle Tracing Analysis (NTA) was used to ensure that hIPSC-CM-derived EXOs were sufficiently isolated. NTA was performed using a NanoSight NS300 (Malvern Instruments Ltd., Worcestershire, UK) instrument. For this purpose, EXOs secreted during hypoxic conditions were analyzed. All samples were diluted to 1:100 and the following settings were selected, following the manufacturer’s manual guide (NanoSight NS300 User Manual, MAN0541-01-EN-00, 2017): The camera level was adjusted so the particles were distinctly visible (camera level 12) and the autofocus was adjusted to avoid detecting indistinct particles. For each sample, five 60 s measurements were captured under these conditions: temperature (21 °C), syringe pump speed (70). All measurements were analyzed using the in-built NanoSight Software NTA 3.2 Dev Build 3.2.16, with the detection threshold set to 8. Hardware: camera; type: sCMOS; laser type: Blue405. To minimize data skewing from single large particles, the number of measured particles always exceeded that of the proposed minimum.
4.4. Culturing and Maintenance of Human IPSC-Derived Cardiomyocytes
Firstly, the Ncyte™ Cardioplate™ Maestro™ MEA 96 with pre-seeded hIPSC-CMs (10^4^ cells/well) was maintained in maintenance medium at 21% O_2_, 5% CO_2_ in a humidified atmosphere at 37 °C until stable electrophysiological activity was observed using the MEA system. Each Ncyte™ Cardioplate™ Maestro™ MEA 96 well contained 8 poly(3,4-ethylenedioxythiophere) (PEDOT) microelectrodes. A total of 20 wells were selected as a control group, and 20 wells were selected for EXO precondition. Well placement was evenly distributed for both groups to avoid skewing of the data based on the well location on the plate.
4.5. EXO Preconditioning
As a preconditional treatment, EXOs resuspended in Cardiomyocyte Culture Media were administered to hIPSC-CMs pre-seeded in Ncyte™ Cardioplate™ Maestro™ MEA 96, as described above. Cells were administered EXOs (200 μL/well, 9.2 × 10^4^ EXO/μL) or regular maintenance medium (200 μL/well) and the plate was incubated for 24 h at 21% O_2_, 5% CO_2_ and at 37 °C.
4.6. Hypoxia
Directly following EXO preconditioning, all cells were thoroughly washed with sterile PBS (1X) to remove any trace of the culture media before the cells were exposed to 16 h of hypoxic stress: incubation with hypoxia medium (200 μL/well) at 1% O_2_, 5% CO_2_, 94% N_2_ at 37 °C [87,88].
4.7. Analysis of Cardiac Electrophysiology on MEA System
Cells were stabilized for 10 min in the MEA system (Maestro Edge, Axion Biosystem, Atlanta, GA, USA) at 21% O_2_, 5% CO_2_ and at 37 °C. Cardiac electrophysiological activity was recorded using AxIS Navigator^TM^ version 3.9.1. Using the MEA system, field potential duration (FPD), beat period, FPD spike slope, FPD spike amplitude, excitation–contraction (EC) coupling, cardiac action potential duration (APD), beat amplitude of contraction, and local extracellular action potential (LEAP) were recorded and analyzed. LEAP induces stronger cell–electrode coupling by applying electrical signals to the electrodes. This alters the signal amplitude and shape, changing the signal from a field potential morphology to more closely resemble a cardiac action potential morphology [88]. APD30, APD50, and APD90 represent time to 30%, 50%, and 90% membrane repolarization, respectively. Data were recorded at baseline (normoxia), directly after EXO preconditioning (24 h), and directly after hypoxia (16 h). Analysis was performed using Axion software: the AxIS Metric Plotting Tool^TM^ (version 2.4) and Cardiac Analysis Tool^TM^ (version 3.3) (Axion Biosystems, Atlanta, GA, USA).
4.8. RNA Isolation
hIPSC-CMs exposed to hypoxic stress within the MEA system were gently washed with PBS (1X, 200 μL). Then, QIAzol lysis reagent (100 μL) (Qiagen, #79306, Hilden, Germany) was added to the cells and the lysate was collected and pooled into three samples for the control group and three samples for the EXO-preconditioned group. Subsequently, RNA was isolated using the miRNeasy Micro Kit (Qiagen, #217084, Hilden, Germany) following the manufacturer’s instructions.
4.9. miR Sequencing
miR sequencing was performed to evaluate the impact of the EXO preconditioning on the miR transcriptome of the hIPSC-CMs. The library preparation and sequencing service was provided by the Genomics Core Facility (GCF), Norwegian University of Science and Technology (NTNU). The GCF is funded by the Faculty of Medicine and Health Sciences at NTNU and the Central Norway Regional Health Authority. First, RNA sample quality control was performed using the RNA 6000 Pico Kit (Agilent, #5067-1513, Santa Clara, CA, USA) with a 2100 Bioanalyzer System (Agilent, #G2939BA, Santa Clara, CA, USA) following the manufacturer’s instructions. Then, a NEXTFLEX^®^ Small RNA-Seq Kit v4 with UDIs (PerkinElmer, #NOVA-5132-44 and #NOVA-5132-32, Waltham, MA, USA) was used to generate the sample library, and a 4200 TapeStation system (Agilent, Santa Clara, CA, USA) was used for quality control of the sample library. Based on the TapeStation data, sample concentrations were adjusted and then quantified using a Qubit Q32853 with the Qubit™ dsDNA BR Assay Kit (Invitrogen, #Q32850, Waltham, MA, USA). Finally, sequencing was performed on a NovaSeq 6000 (Illumina, San Diego, CA, USA) performing single reads, with 101 cycles for read 1, 10 cycles for index 1, and 10 cycles for index 2.
4.10. miR Target Prediction Analysis
After miR sequencing, raw data were additionally processed by the GCF. Utilizing the DESeq2 package in R [89], statistical analyses were conducted to determine the significance of miR expression differences across experimental conditions. The process involved normalizing raw count data for sequencing depth, estimating and reducing gene-wise dispersions for increased stability, and fitting a negative binomial model suitable for count data with overdispersion. The Wald test yielded a p-value, indicating differential expression. The Benjamini–Hochberg procedure was subsequently applied to account for the false discovery rate arising from repeated testing, yielding an adjusted p-value. The result of this process was the production of a datasheet consisting of 5 columns and 1028 rows. The column names were as follows: miR immature names, baseMean, log2FoldChange, lfcSE, stat, pvalue, padj (Table S2). All miRs with an adjusted p-value of <0.2 were selected and subjected to DIANA tool miRPath 4.0 for mature miRNA name retrieval [90]. This yielded 10 miRs. The resulting list of miRs with mature miRNA names was analyzed in R.
The list of 10 miR names was queried in three databases (miRecords, miRTarBase, and TarBase) for validated miR–target interactions using the multimiR package in R [91,92,93,94]. All unique gene targets were converted into ENTREZID using the clusterProfiler package, with 0.12% of 3450 gene names failing to map [95]. Overrepresentation analysis was performed on the resulting miR gene targets in the clusterProfiler package for Gene Ontology (GO) enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis [96,97].
For the GO enrichment analysis, molecular function, biological process, and cellular component, all three of which are subcategories of GO terms in humans, were enriched from the target genes with the parameters of the p-value and q-value (Benjamini–Hochberg-adjusted p-value) set to a cut-off of 0.05. The p-value adjustment method was set to Benjamini–Hochberg. The results were extracted and ranked by adjusted p-value in a descending manner. The top 30 enriched GO terms were then plotted onto a dot plot.
KEGG Enrichment Analysis, all target genes were enriched via the clusterProfiler package with the organism set to human and the p-value and q-value cut-offs set to 0.05. The p-value adjustment method was set to Benjamini–Hochberg. Then, the BRITE hierarchy parent terms for each KEGG term were manually annotated based on the KEGG database description. All statistically significant KEGG terms were plotted onto a KEGG enrichment graph.
Based on the top terms of both GO and KEGG enrichment analyses, detailed miR-target interactions were manually examined for their relevance in the experimental context through a comprehensive screening based on target gene tissue expression (The Human Protein Atlas and GeneCards) and published studies on Web of Science and PubMed.
Lastly, a Sankey diagram was generated based on miRs, log2Fold Change, and impact columns (positive or negative) to demonstrate miR target gene effects. miRNAs with a <0.2 adjusted p-value were selected for the first column. Gene targets were selected for the second column, with function impact categorized into four areas: viability impairment, cardiac dysfunction, cardioprotection, and contractility/enhanced cardiac function. miRNAs were ranked based on the log2 fold changes detected, with the flow demonstrating either inhibition or promotion of the gene target and function impact.
4.11. Statistical Analysis
Statistical analysis of the NTA data was performed with the NanoSight Software NTA 3.2 Dev Build 3.2.16. GraphPad Prism 9 (GraphPad Software, San Diego, CA, USA) was used for statistical analyses of the MEA data. Significant differences among the samples were determined using one-way ANOVA or two-tailed independent sample t-test, or mixed-effects analysis: Šídák’s multiple comparisons test, depending on analysis. Quantitative data are expressed as the mean of raw values or %-change from baseline ± SEM. A minimum p-value of ≤0.05 was considered significant. miR target prediction analysis was conducted in R 4.3.1, utilizing several packages for over-representation analysis [98]. R packages: tidyverse [99,100], clusterProfiler [95,101], org.Hs.eg.db [100], DOSE [101], multiMiR [94,95,96,97,98,99,100,101,102]. The resulting gene targets were evaluated and ranked based on their adjusted p-values, with the most significant findings being selected for further interpretation in the context of the experiment.
5. Conclusions
This study provides new insights into vesicle-based cardioprotective strategies, specifically EXO-mediated cell–cell signaling between hypoxic hIPSC-CMs. We show that hIPSC-CMs preconditioned with hypoxic EXOs demonstrated an increased beat period and enhanced EC-coupling efficiency after hypoxic stress, suggesting potential cardioprotective benefits by reducing myocardial oxygen consumption and workload, and improving contraction cycles and overall cardiac function.
This study also highlighted the intricate involvement of miRs in these responses. Next-generation sequencing identified 10 differentially regulated miRs linked to key cardiac functions, including cell survival, contraction, apoptosis, ROS regulation, and ion channel modulation. These findings represent the potential for miR-targeted therapies in the future.
Overall, our results reveal novel aspects of EXO-mediated cardioprotection, paving the way for targeted interventions to improve outcomes in ischemic heart disease. Further studies on the identified miRs could enhance our understanding and treatment of ischemic heart disease.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Nowbar A.N. Gitto M. Howard J.P. Francis D.P. Al-Lamee R. Mortality from ischemic heart disease: Analysis of data from the World Health Organization and coronary artery disease risk factors From NCD Risk Factor Collaboration Circ. Cardiovasc. Qual. Outcomes 201912 e 00537510.1161/CIRCOUTCOMES.118.00537531163980 PMC 6613716 · doi ↗ · pubmed ↗
- 2Mosterd A. Hoes A.W. Clinical epidemiology of heart failure Heart 2007931137114610.1136/hrt.2003.02527017699180 PMC 1955040 · doi ↗ · pubmed ↗
- 3Greco S. Gaetano C. Martelli F. Hypoxami R regulation and function in ischemic cardiovascular diseases Antioxid. Redox Signal.2014211202121910.1089/ars.2013.540324053126 PMC 4142792 · doi ↗ · pubmed ↗
- 4Ertracht O. Malka A. Atar S. Binah O. The mitochondria as a target for cardioprotection in acute myocardial ischemia Pharmacol. Ther.2014142334010.1016/j.pharmthera.2013.11.00324275322 · doi ↗ · pubmed ↗
- 5Kalogeris T. Baines C.P. Krenz M. Korthuis R.J. Cell biology of ischemia/reperfusion injury Int. Rev. Cell Mol. Biol.20122982293172287810810.1016/B 978-0-12-394309-5.00006-7PMC 3904795 · doi ↗ · pubmed ↗
- 6Huang C.L. Lei M. Cardiomyocyte electrophysiology and its modulation: Current views and future prospects Philos. Trans. R. Soc. B 20233782022016010.1098/rstb.2022.0160 PMC 1015021937122224 · doi ↗ · pubmed ↗
- 7Yang D. Deschênes I. Fu J.D. Multilayer control of cardiac electrophysiology by micro RN As J. Mol. Cell. Cardiol.202216610711510.1016/j.yjmcc.2022.02.00735247375 PMC 9035102 · doi ↗ · pubmed ↗
- 8Benjamin E.J. Muntner P. Alonso A. Bittencourt M.S. Callaway C.W. Carson A.P. Chamberlain A.M. Chang A.R. Cheng S. Das S.R. Heart disease and stroke statistics—2019 update: A report from the American Heart Association Circulation 2019139 e 56e 52810.1161/CIR.000000000000065930700139 · doi ↗ · pubmed ↗