Transcription Factor CcFoxO Mediated the Transition from Summer Form to Winter Form in Cacopsylla chinensis
Chuchu Wei, Songdou Zhang

TL;DR
This study explores how the transcription factor CcFoxO helps the insect Cacopsylla chinensis transition from summer to winter form under cold stress.
Contribution
The study identifies CcFoxO as a key regulator of seasonal polyphenism in Cacopsylla chinensis, linking it to energy storage and morphological changes.
Findings
CcFoxO is responsive to low temperatures and regulated by CcTRPM.
CcFoxO influences energy storage by accumulating triglycerides and glycogen.
Knockdown of CcFoxO leads to high mortality and failed seasonal transition.
Abstract
Amid global climate change featuring erratic temperature fluctuations, insects adapt via seasonal polyphenism, essential for population sustainability and reproductive success. Cacopsylla chinensis, influenced by environment variations, displays a distinct summer form and winter form distinguished by significant morphological variations. Previous studies have highlighted the role of temperature receptor CcTPRM in orchestrating the transition in response to 10 °C temperature. Nevertheless, the contribution of the transcription factor FoxO in this process has remained ambiguous. Here, we aimed to explore the correlation between C. chinensis FoxO (CcFoxO) and cold stress responses, while identifying potential energetic substances for monitoring physiological shifts during this transition from summer to winter form under cold stress by using RNAi. Initially, CcFoxO emerges as responsive to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
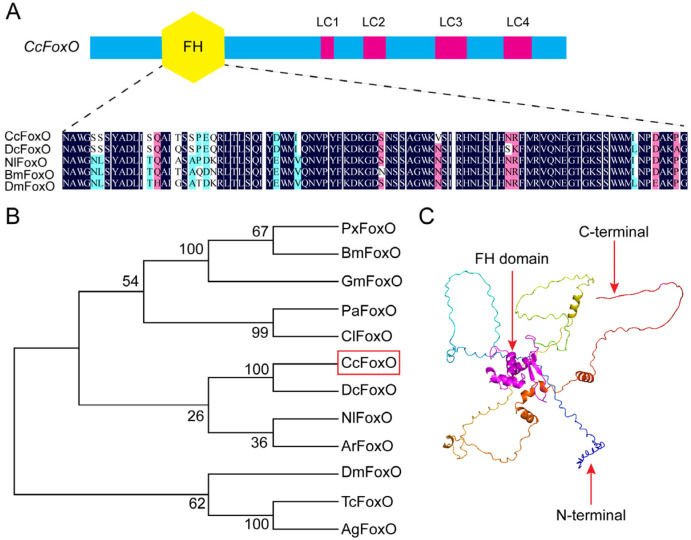 Figure 1
Figure 1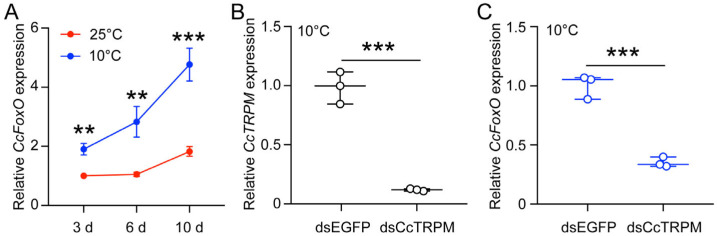 Figure 2
Figure 2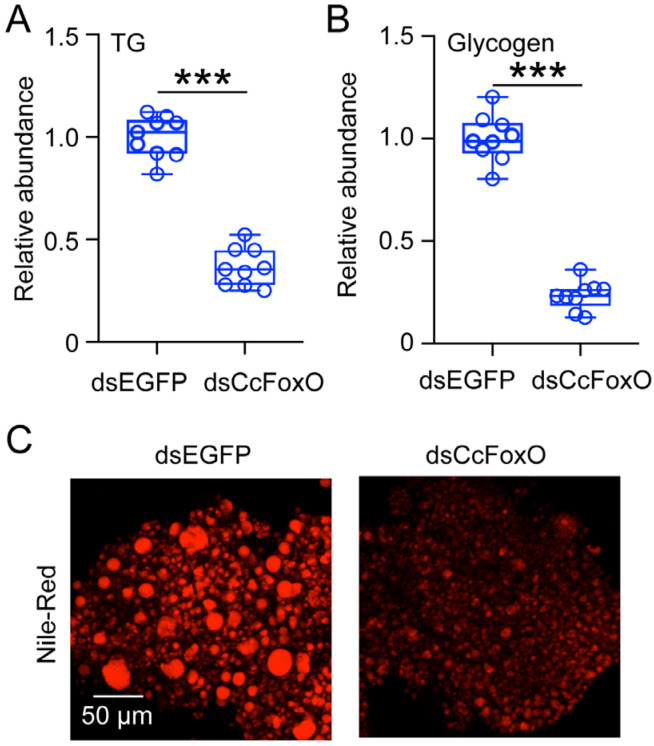 Figure 3
Figure 3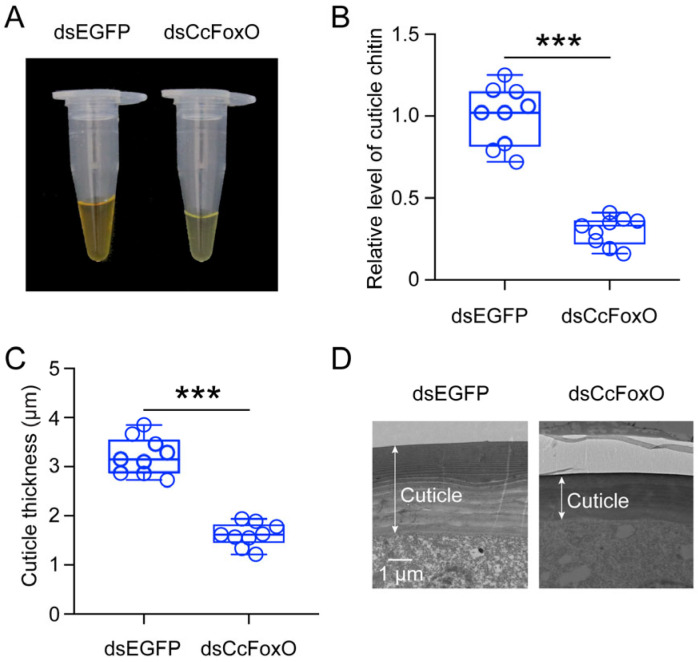 Figure 4
Figure 4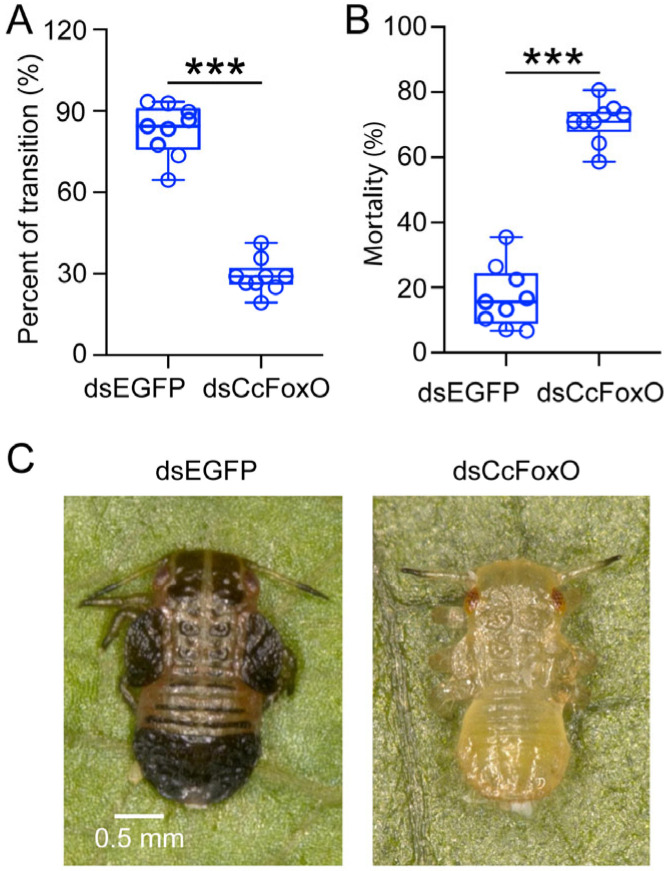 Figure 5
Figure 5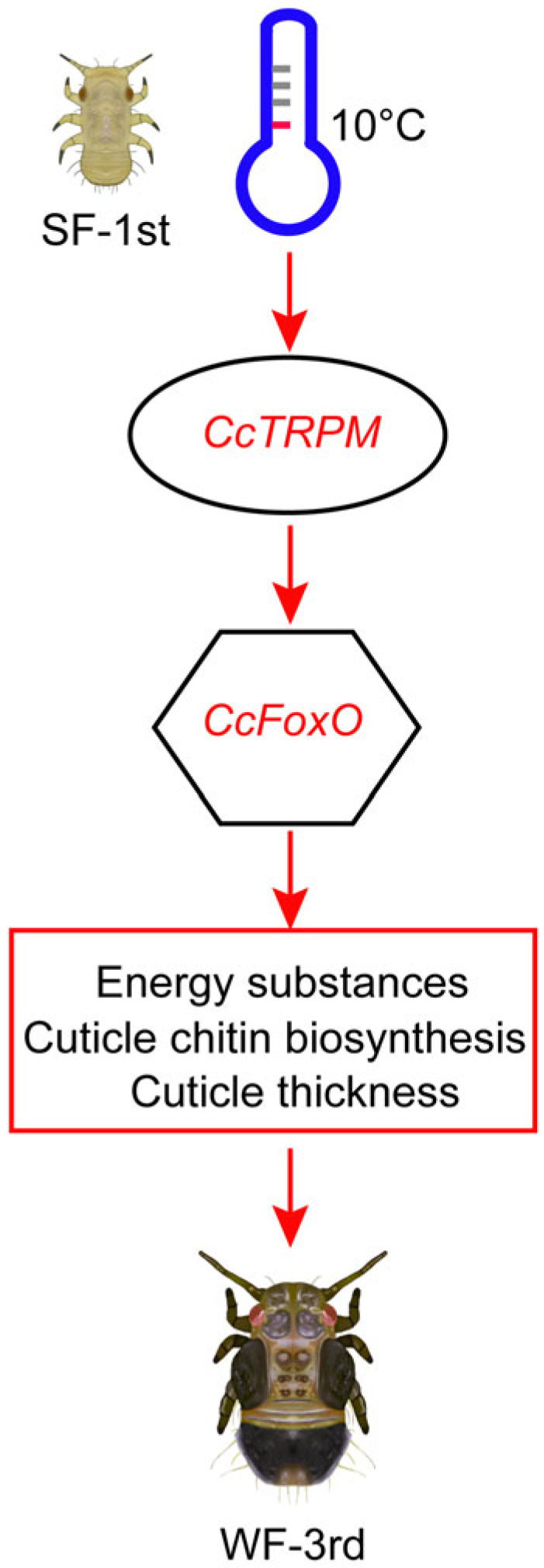 Figure 6
Figure 6- —National Natural Science Foundation of China
- —China Agriculture Research System
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurobiology and Insect Physiology Research · Physiological and biochemical adaptations · Insect and Arachnid Ecology and Behavior
1. Introduction
The advent of global climate change has introduced formidable obstacles, marked by frequent occurrences of extreme weather events and escalating climatic unpredictability causing habitat alterations. Insects, with evolutionary origins dating back 350 million years or more to the Devonian period of the Paleozoic Era, have persevered through ice ages, warm interglacial periods, and now navigate the current era of climate change [1]. Insects that thrive in different habitats are considered “winners in evolution”, showing great adaptability. Polyphenism, a production of gene expression regulation, offers greater flexibility in response to complex environmental changes, enabling a single genotype to manifest multiple phenotypes under varied environmental stresses [2]. Prior researchers have partially unveiled the factors that regulate polyphenism. These factors include biotic elements like pheromones, along with abiotic elements such as population density, nutrition, photoperiod, and temperature [3,4]. Polyphenism shows how insects are smart in adapting to nature, working well with changes in the environment to survive, thereby bolstering the stability of insect populations crucial for their reproductive success [1]. Leveraging this mechanism of adaptive response, humans can better understand how to manage agricultural population under certain environmental conditions. For example, people can use RNA interference (RNAi) to prepare RNA biopesticides based on the key target genes of the adaptive process to control specific pests.
Despite the importance of polyphenism, contemporary investigations center on model insects. For example, in response to nutritional deficiencies pea aphids undergo wing development, transitioning from wingless to winged forms [5]. Early sources of nutrients can affect the caste differentiation in ants [6]. The underlying molecular mechanisms of these phenomenon are particularly fascinating. Recent discoveries have unveiled the involvement of neuroendocrine hormone signaling and transcription factors in their regulation. The insulin signaling pathway plays an important role in wing differentiation [7]. Insulin receptors assume a pivotal role in determining the differentiation of long and short wings in the brown planthopper [8], while FoxO exerts negative control over the development of wing morphology in pea aphids amidst varying environmental circumstances [5]. Moreove, the pathway also plays a role in wing polyphenism in Pyrrhocoris apterus [9]. Responding to population density, the dopamine signaling pathway regulates the switch of locusts from solitary to social behavior, with dopamine phosphorylation aiding in this shift [10,11]. In Harpegnathos saltator, kr-h1 serves as a key transcription factor responsive to hormonal signals like juvenile hormone (JH) and ecdysteroid (20E), binding to and regulating different target genes in worker ants and queen ants to establish their social hierarchy [12]. However, research on multiple transcription factors that ultimately regulate polyphenism in non-model species responses remains scarce and unclear.
Cacopsylla chinensis (Yang & Li), a member of Hemiptera: Psyllidae, exhibits a life cycle encompassing eggs, nymphs, and adults, posing a significant threat to pear orchards. In October 2011, 9.5 ha psyllid-infected orchards were defoliated in Japan [13]. With its piercing-sucking mouthpart, this insect induces leaf deformities and necrotic spots by extracting sap from pear leaves [14]. Additionally, it serves as a crucial vector for transmitting the economically detrimental Erwinia amylovora disease, which systematically attacks pear trees, leading to flower clusters dropping, fruits deforming, and branches withering, culminating in profound devastation [13]. Rather than entering diapause, C. chinensis survives the adverse environment through seasonal polyphenism, namely summer form and winter form [15]. Morphologically, the summer form displays a yellow–green hue and smaller stature, while the winter form presents as dark-brown and larger in size. As temperatures decrease, C. chinensis typically lays eggs along leaf veins before the leaf fall. The hatched nymphs exhibit traits suited for winter conditions (0–10 °C). Subsequently, with the temperature rise in early spring (15–25 °C), they lay eggs on rough bark, dormant buds, leaves, and flower buds, giving rise to summer-form nymphs [16]. Previous studies have identified the molecular mechanisms orchestrating phenotypic shifts in response to temperature variations. The third instar stage emerges as a critical juncture for the transition from summer form to winter form in C. chinensis. CcTRPC3 acts as a cold receptor, sensing the low temperature around 10 °C and triggering the expression of cuticle-binding proteins CcCPR4 and CcCPR9, eventually leading to alterations in total cuticle pigmentation, chitin content, and cuticle thickness [17]. Additionally, the low-temperature receptor CcTRPM influences the expression of rate-limiting enzymes in chitin biosynthesis, namely CcTre1 and CcCHS1. The maintenance of the summer to winter transition is mediated by the stable expression of CcTRPM regulated by miR-252 [18]. Nevertheless, the precise regulation mechanism by downstream signaling, especially transcription factors, in response to temperature receptors remains elusive.
FoxO encodes a FOXO transcription factor belonging to the FOX superfamily’s “O” branch, featuring a highly conserved DNA-binding domain named the forkhead domain. The forkhead (FH) domain comprises three α-helices, three β-folds, a loop structure, and the C-terminus, crucial for DNA binding and specificity recognizing DNA sequences. Initially identified in human tumors, FoxO was recognized as a chromosomal translocation partner [19]. Mammals harbor four FoxOs-FoxO1, FoxO3a, FoxO4, and FoxO6 located on distinct chromosomes, while invertebrates like Caenorhabditis elegans possess a single FoxO gene, daf-16, and Drosophila melanogaster houses dFoxO [20].
Researchers on FoxO in insects have primarily focused on regulating 20E and JH interactions during molting and metamorphosis, alongside attention to diapause, longevity, stress response, and other key physiological processes in insects [20,21]. In Helicoverpa armigera and Bombyx mori molting, 20E triggers increased FoxO expression and nuclear translocation, enhancing lipolysis in adipose cells [22,23]. In the developing ovary of Harpegnathos saltator queen ant, the insulin-resistant Imp-L2 reduces IIS-AKT/FOXO signaling pathway activity in the fat body, leading to dephosphorylation and nucleus translocation of FoxO, thereby activating longevity-associated genes [24]. In lagomorphs, FoxO aids in rapid responses to low temperatures, enabling survival under cold conditions by regulating gene expression associated with cold shock [25]. As a pivotal regulator downstream of the insulin signaling pathway, FoxO plays a crucial role in managing energy metabolism, influencing cellular energy balance, and closely relating to developmental processes and resistance to environmental stressors [26]. Consequently, CcFoxO might play a significant role in the shift from summer form to winter form in C. chinensis, which further elucidation is warranted.
To investigate the involvement of CcFoxO in the transition from summer form to winter form of C. chinensis, we utilized RNAi technology to disrupt the expression of CcFoxO and CcTRPM, studying the relationship between coldness and the function of CcFoxO. Phenotypic alterations were assessed by measuring total cuticle pigment content, cuticle chitin content, and cuticle thickness. Our study unveiled a significant role for CcFoxO in regulating the transition in response to low temperatures, impacting triglyceride and glycogen metabolism. This research aims to enhance understanding of the physiological function of the transcription factor FoxO, shed light on the ecological adaptation mechanism of C. chinensis, and potentially identify target genes for the effective control of psyllid pests.
2. Results
2.1. Cloing and Sequence Analysis of CcFoxO
The cDNA fragment, putatively encoding a forkhead box O protein, was identified as CcFoxO in C. chinensis. The full-length sequence of CcFoxO was confirmed through RT-PCR using specific primers (Table S1) and subsequent sequencing. Sequence analysis revealed that the open reading frame (ORF) of CcFoxO spanned 1296 bp, encoding a polypeptide of 431 amino acids with a predicted molecular weight of 47.67 kDa and a pI of 4.80. Alignment analysis demonstrated a substantial amino acid identity of the FH domains of CcFoxO with FoxO sequences from four other selected insect species (Figure 1A). In an evolutionary perspective, CcFoxO exhibited the closest relationship with the DcFoxO homologue (Diaphorina citri, XP_026688498.1), both being significant Hemiptera pests of fruit trees (Figure 1B). The potential tertiary protein structure of CcFoxO was predicted using the online server Alphafold3 and refined with PyMOL-v1.3r1 software, revealing the presence of the conserved FH domain (Figure 1C).
2.2. Temporal and Spatial Expression Patterns of CcFoxO
The newly hatched summer-form nymphs were cultured at 10 °C and 25 °C, and the relative expression levels of CcFoxO were detected at 3, 6, and 10 days. Results from qRT-PCR revealed significantly higher expression levels at 10 °C (1.90, 2.83, and 4.77) compared to 25 °C (1.00, 1.05, and 1.82) (p values = 0.00138, 0.00419, and 0.00092) (Figure 2A). The differential expression under the two temperature conditions indicated that low-temperature exposure significantly stimulated the expression of CcFoxO.
Under the 10 °C condition, knockdown of CcTRPM led to a notable decrease in the relative expression of CcTRPM, demonstrating the high efficiency of the RNAi experimental system (Figure 2B). Intriguingly, the downregulation of CcTRPM expression also considerably reduced the relative expression of CcFoxO (Figure 2C). The observed positive correlation between CcFoxO and CcTRPM expression manifests that these two genes are interconnected within the same signaling pathway.
2.3. Impact of CcFoxO on Triglyceride and Glycogen Levels in C. chinensis
The newly hatched summer-form nymphs were subjected to 10 °C conditions and fed with dsEGFP in the control group and dsCcFoxO in the experimental group. After a 15-day treatment period, triglyceride and glycogen levels were assessed. Each treatment consisted of nine biological replicates, with 30 nymphs per replicate. Analysis showed a significant reduction in triglyceride and glycogen levels following dsCcFoxO treatment compared to nymphs treated with dsEGFP (Figure 3A,B). Additionally, Nile red staining demonstrated a marked decrease in lipid droplet abundance post-interference with CcFoxO expression based on both the lightness and size (Figure 3C).
These discoveries suggest that CcFoxO plays a crucial role in maintaining the abundance of triglycerides and glycogen, essential substances representing the primary forms of lipids and carbohydrates in insects. These compounds serve as essential nutrient reserves, indicating that CcFoxO is involved in nutrient metabolism under cold temperature.
2.4. Involvement of CcFoxO in Regulating Total Cuticle Pigment, Cuticle Chitin Content, and Cuticle Thickness of C. chinensis
Following a 15-day dsCcFoxO treatment of first-instar nymphs, the total cuticle pigment content was significantly lower compared to the dsEGFP treatment group. The visible color difference in the pigment extract was noticeable to the naked eye (Figure 4A). Under disrupted CcFoxO expression conditions, first-instar summer-form nymphs were maintained at 10 °C. After 15 days, the cuticle chitin content (1.00) and thickness (3.22 μm) were significantly reduced than those treated with dsEGFP (0.30, 1.62 μm) (Figure 4B,C). Significant differences in cuticle thickness were also observed in the representative TEM image (Figure 4D).
These results indicated that disruption of CcFoxO affects C. chinensis’s ability to adapt to low temperatures in terms of cuticle composition and thickness, highlighting the role of CcFoxO in epidermal alterations.
2.5. Role of CcFoxO in Modulating the Transition from Summer Form to Winter Form
At 10 °C, the results of nymphs treated with dsEGFP or dsCcFoxO are divided into two categories: successful transformation or death. The transition percent of summer-form first instar nymphs to winter-form nymphs significantly decreased after 15 days of dsCcFoxO feeding (from approximate 82.85% to 29.09%) (p value = 1.89282 × 10^−10^) (Figure 5A). Moreover, the mortality under this treatment was significantly higher than that of the dsEGFP treatment group (from approximate 17.15% to 70.91%) (Figure 5B). Notably, nymphs in the dsFoxO-treated group exhibited light-colored, yellow–green, and small traits after the 15-day period, while those in the dsEGFP-treated group displayed dark, black–brown, and slightly larger characteristics (Figure 5C). These observations suggest that CcFoxO is involved in the transition from the summer form to winter form of C. chinensis, actively facilitating this transition under low-temperature conditions.
3. Discussion
The phenomenon of polyphenism is widespread in the insect realm, with current research predominantly focusing on model insects like butterflies, moths, aphids, locusts, ants, and bees [27]. Among these, the mechanisms of wing dimorphism in aphids stands out as a well-studied example following locusts’ morphological transformations [28,29]. These reported phenomena are primarily influenced by population density, alongside factors such as nutrition, temperature, and photoperiod [30]. The shift from a solitary phase to gregarious phase in locusts is governed by a biogenic amine signaling pathway [31]. Polyphenism, which entails the expression of different phenotypes from the same genetic background under different biotic or abiotic stresses, plays a pivotal role in the survival and reproduction of insects [2]. Some insects exhibit seasonal polyphenism, exemplified by cases such as Nemoria arizonaria, where caterpillar closely mimic the seasonal morphology of the host plant [32], and Bicyclus anynana, where the size of hindwing eyepatches changes with the seasons [33]. The seasonal polyphenism of these two insects may help them adapt to temperature changes along with season on the one hand, and also help them disguise themselves to avoid being hunted by natural enemies on the other hand. Similarly, the subject of this study, C. chinensis, demonstrates seasonal polyphenism by responding to 10 °C and transitioning from summer-form to winter-form phenotypes [18]. Cuticle thickness, cuticle chitin content, and cuticle pigment content, regulated by bursicon, serve as discerning indicators for phenotypic variations between summer and winter variants [34].
In this study, expression analysis of CcFoxO revealed an upregulation in its expression in response to low temperature. Additionally, the expression patterns of CcFoxO and CcTRPM were consistent under low-temperature conditions based on our previous publication [18]. CcTRPM, identified as a temperature receptor in C. chinensis, responds to low temperatures, regulates chitin synthesis, influences cuticle characteristics, and aids in reducing heat loss in cold temperatures [18]. Thicker skin and darker body color help psyllids better reduce water loss and protect body temperature in the cold winter to adapt to the environment. Treatment with dsCcFoxO hindered the changes in cuticle properties and total pigment content, resulting in a significant decrease in the transition percent from summer form to winter form and an increase in mortality. Based on these findings, it is reasonable to infer that CcFoxO acts downstream of CcTRPM, linking these two genes and mediating insect physiological processes in C. chinensis triggered by low temperatures. Insects such as ectotherms experience fluctuations in body temperature according to the surrounding environment. To combat challenges like cold stress and food scarcity during winter, many species accumulate fat reserves beforehand. Fat serves as an energy reservoir during colder months and significantly affects reproductive capacity [35]. Notably, disruption of CcFoxO expression led to reduced triglycerides and glycogen content, accompanied by a partial absence of lipid droplets. This highlights the involvement of CcFoxO in the transition from the summer form to winter form of C. chinensis, affecting energy reserves, cuticle chitin biosynthesis, and cuticle properties, aligning with previous findings. For instance, Aedes aegypti increases triacylglycerol accumulation in dormant eggs to cope with cold environments as it migrates from tropical to temperate regions. The species adjusts energy reserves by increasing proteins and carbohydrates when triacylglycerol levels are low in a given population [36]. Similarly, insects enhance cold tolerance by increasing glycogen reserves during cold stasis. Glycogen broken down into sugar alcohols and protective compounds aids in surviving low temperatures [37]. Therefore, the elevation in triglycerides and glycogen, regulated by CcFoxO, represents an adaptation to cold conditions during the winter-form stage. Under dsCcFoxO treatment, the regulating of glycogen and triglycerides may be the first and then decrease in the transition percent.
Previous researchers have recognized the FoxO family gene as a forkhead transcription factor present in both mammals and insects. Following dsCcFoxO treatment, C. chinensis failed to exhibit corresponding phenotypic changes in response to low temperatures, showing no alterations in cuticle properties or pigment content. FoxO is known to be involved in the PI3K/Akt signaling pathway and the MAPK signaling pathway, impacting FoxO activity through phosphorylation and nuclear localization. The PI3K/Akt pathway is vital for cell growth, metabolism, and survival, responding to insulin and growth factors [38]. The MAPK pathway triggers FOXO phosphorylation in response to stressors, affecting cellular responses to oxidative stress, nutrient deprivation, and other external stimuli [39]. Both pathways influence cellular metabolism, including the metabolism of energy substrates like glycogen and triglycerides. The involvement of CcFoxO in these signal pathways also may be the reason for high mortality in dsCcFoxO treatment. While our study confirms the role of CcFoxO in regulating glycogen and triglycerides in response to low temperatures, the upstream regulatory components remain ambiguous and warrant further exploration. Further studies could explore the involvement of specific signaling pathways by monitoring changes in associated indicators.
Unlike prior research focusing on FoxO’s role in modulating ecdysteroids and juvenile hormones during insect molting, this study sheds light on FoxO’s reaction to seasonal polyphenism for the first time. Short-daylight and low-temperature conditions elevate FoxO activity in the moth H. armigera, reducing TGFβ signaling and inducing pupal diapause [40]. Both diapause and seasonal polyphenism exhibit protective mechanisms against low temperatures, underscoring FoxO’s involvement in responding to cold conditions in both phenomena. Studies on FoxO can enhance our understanding of its functional roles and elucidate the correlation between diapause and seasonal polyphenism by investigating the governing signaling pathways. As shown in Figure 6, at 10 °C, the low-temperature receptor CcTRPM was activated in first instar summer-form nymphs leading to increased expression of CcFoxO, facilitating energy accumulation, promoting cuticle chitin synthesis, and increasing cuticle thickness. Subsequently, the nymphs transitioned into the third instar winter form, enhancing adaptability to low temperatures and reducing mortality rates. These findings not only deepen our understanding of FoxO’s functionality but also provide new insights for environmentally friendly prevention and the control of C. chinensis.
4. Materials and Methods
4.1. Host Plant and Insect Rearing
The summer form and winter form of C. chinensis were collected from the “Lixiangzhuangyuan” pear orchards in Daxing District, Beijing City, China, during the summer and later fall seasons, respectively. The wild-type populations were purified through more than 20 generations of inbreeding in the integrated pest management (IPM) laboratory at China Agricultural University. Nymphs and adults from both forms were reared on pear seedlings enclosed with dense mesh screens. The summer form was reared at 25 ± 1 °C and 65 ± 5% relative humidity, while the winter-form was reared at 10 ± 1 °C and 25 ± 5% relative humidity, both under a 12 L: 12 D photoperiod. The host plants, 2–3-year old Korla fragrant pear seedlings ranging from 60–100 cm in height, were propagated from cuttings and maintained with regular watering and fertilization.
4.2. Molecular Characterization of CcFoxO
The FoxO homolog was identified from the transcriptome database of C. chinensis, and the accurately sequenced fragment was designated as CcFoxO (GenBank accession number: PP931994). The SMART online tool (http://smart.embl-heidelberg.de/, accessed on 25 June 2024) was used for predicting conserved domains. Subsequently, the three-dimensional structure of the protein was predicted using Alphafold 3 (https://alphafoldserver.com/, accessed on 25 June 2024), with further modifications performed using PyMOL-v1.3r1 software. Homologous protein sequences of CcFoxO were retrieved from NCBI BLASTP (https://blast.ncbi.nlm.nih.gov/, accessed on 25 June 2024) for 3 Lepidoptera, 3 Hemiptera, 2 Hymenoptera, and 2 Coleoptera insect species. The selected protein sequences exhibit lower E values and higher homology across various species in the search results, encompassing both closely and distantly related organisms. This approach enhances the clarity and accuracy of phylogenetic tree presentation. Amino acid sequence alignment analysis of the 12 protein sequences mentioned above was performed using DNAman 9.0 software. Phylogenetic analysis and the construction of a phylogenetic tree were carried out using the neighbor-joining (NJ) method in MEGA10.1.8 software.
4.3. qRT-PCR Analysis for mRNAs
To determine the mRNA expression levels of CcFoxO under different temperature conditions, summer-form 1st instar nymphs were gathered at 3, 6, and 10 days post-exposure to temperature of 25 and 10 °C in the incubators. Subsequently, these nymphs were promptly preserved in an ultra-low temperature freezer (−80 °C) for total RNA extraction.
Total RNA extraction for mRNA from the above samples was performed, employing the TaKaRa RNA Extraction kit (Cat# 9767, TaKaRa, Shiga, Japan) based on the kit protocol. Subsequently, the synthesis of the first-strand cDNAs was carried out using the Takara PrimeScript RT reagent kit (cat. no. RR047A, Takara, Shiga, Japan) following the manufacturer’s guidelines. The quantification of mRNA was executed utilizing the Bio-Rad CFX Connect Real-Time PCR System (Bio-Rad, Hercules, CA, USA) with the Takara TB Green Premix Ex TaqII kit (catalog no. RR820A, Takara, Shiga, Japan). Ccβ-actin (GenBank accession number: OQ658571) was utilized as the reference gene in the qRT-PCR analysis. Specific primer sequences can be found in Table S1 and the melting curve of the qRT-PCR primer for CcFoxO was in Figure S2.
4.4. Synthesis of Double-Stranded RNA and RNAi Assays
The production of double-stranded RNAs (dsRNA) targeting EGFP, CcTRPM, and CcFoxO was accomplished using the MEGAscript T7 high yield transcription kit (Cat#AM1334, Thermo Fisher Scientific) with primers (Table S1) integrated with T7 RNA polymerase promoter sequences. The synthesized dsRNAs were purfied using MEGAclear columns (Ambion) and eluted in nuclease-free water treated with diethyl pyrocarbonate (DEPC). The concentration and purity of the dsRNA were assessed through ultraviolet spectrophotometry and gel electrophoresis, respectively.
Each biological replicate, including at least 30 nymphs, was administered dsRNAs (1000 ng/μL) targeting various genes through a stem-leaf device. Each treatment was repeated three times. For the stem-leaf device, a fresh pear leaf was inserted into a 250 μL PCR tube containing different concentrations of dsRNAs, and the entire assembly was placed in a 50 mL sterile plastic tube. For the RNAi assays, summer-form 1st instar nymphs were segregated into three groups: (1) Nymphs were sampled at 2 and 4 days under 25 °C, while 3 and 6 days under 10 °C for RNAi efficiency analysis via qRT-PCR. (2) Variations in morphological characteristics were monitored every two days under 10 °C by documenting the number of summer-form and winter-form individuals according to the alterations in body color and external characteristics. (3) Samples were collected at 12 days after dsRNA feeding for the assessment of cuticle ultrastructure, total cuticle pigment content analysis, triglyceride and glycogen content determination, and evaluation of cuticle chitin content under 10 °C condition.
4.5. Transmission Electron Microscopy (TEM) Analysis of Cuticle Ultrastructure
To compare the nymph cuticle ultrastructure, transmission electron microscopy was applied following established procedures [15,18]. Initially, headless body samples were fixed in 4% polyformaldehyde (PFA) for 48 h, followed by post-fixation in 1% osmium tetroxide for 1.5 h. Subsequently, the samples underwent a standard ethanol/acetone series, followed by infiltration and embedded in spurr resin. Superthin sections (approximately 70 nm) of the thorax were prepared, stained with 5% uranyl acetate and Reynolds’ lead citrate solution. Observation, imaging, and measurements of the sections were performed using a Hitachi HT7800 transmission electron microscope operating at 120 kv. Each sample comprised five nymphs.
4.6. Phenotypic Observation of Summer Form and Winter Form
To quantify phenotypic divergence among nymphs under different treatments, three indicators were utilized: total pigment content of the cuticle, cuticle chitin content, and cuticle thickness. The experimental subjects were used 15 days after dsCcFoxO or dsEGFP treatment of 1st-instar nymphs, with 30 nymphs per replicate and 9 replicates for every group.
For the total pigment content analysis, nymph cuticles were dissected and treated with acidified methanol (with 1% concentrated hydrochloric acid). The cuticle tissues were ground, subjected to 24 h of thermostatic oscillator at 200× g and 25 °C, and the resulting supernatants were filtered through a 0.45 μm filter membrane to obtain the total pigment extract post centrifugation.
To analyze cuticle chitin content, staining with Triticum vulgaris lectin–fluorescein (WGA-FITC) was performed. Nymph samples were immobilized in 4% PFA for 12 h at 4 °C, dehydrated with sucrose solutions (10%, 20%, and 30%), and rinsed with sterile 1 × PBS solution. Gradual embedding at −25 °C was achieved using SAKURA Tissue-Tek OCT compound. Ultrathin sections (approximately 70 nm) of insect body materials were obtained with a Leica CM1850 cryotome. Staining of cuticle chitin and cell nucleus in the sections were carried out with WGA-FITC (50 μg/mL) and DAPI (10 μg/mL) separately, followed by washing with sterile PBS buffer. Fluorescence images were captured using a Leica SP8 confocal microscope.
To assess the cuticle chitin content, the chitin ELISA kit (Cat# YS80663B, Yaji Biotechnology, China) was employed following the manufacturer’s instructions. Initially, a 1.5 mL sterile centrifuge tube was used to homogenize the sample, consisting of 50 nymphs per sample, using an electric homogenizer, followed by centrifugation at 3000× g for 20 min. Subsequently, the samples, along with standard samples, were incubated in ELISA plates for approximately 30 min under 37 °C conditions. The ELISA plates were rigorously washed with a washing buffer at least five times (each time for 30 s). The enzyme labeling reagent was then added to the ELISA plates for further incubation and washing. Finally, the A and B solutions of the chromogenic agent were thoroughly mixed for color development under 37 °C conditions and maintained in the dark.
4.7. Analysis of Triglyceride and Glycogen Content
To investigate the energy metabolism of C. chinensis, metabolites associated with cold tolerance and energy were measured using commercial kits. The samples were tested using whole insects following the manufacturer’s instructions, encompassing the triglycerides (TG) assay kit (Cat# A110-1-1, Nanjing, Jiancheng, China) and glycogen assay kit (Cat# A043-1-1, Nanjing, Jiancheng, China) [41]. Each treatment consisted of nine biological replicates, with 50 nymphs per replicate.
4.8. Nile Red Staining
Summer-form nymphs underwent treatment and were subsequently placed on a glass slide. An adequate volume of PBS buffer was added dropwise to separate the fat bodies at room temperature. The isolated tissue was then transferred to a staining dish filled with PBS buffer. After blotting the PBS buffer, the tissue was fixed with 4% paraformaldehyde (PFA) for 30 min at room temperature. Subsequently, the PFA solution was blotted, and the samples were washed three times with PBS for 10 min each. A solution of 0.5 mg/mL Nile red dye was diluted to a ratio of 1:2500 and applied for overnight staining at 4 °C. After blotting the dye, samples were washed three times with PBS, each wash lasting 10 min. The DAPI mother solution at 20 ng/μL was diluted 1:10 and stained for 5 min. DAPI was then blotted, and a PBS wash was performed three times for 10 min each. Finally, after mounting, the samples were photographed, saved, and scanned using a laser confocal microscope.
4.9. Statistical Analysis
IBM SPSS Statistics 26.0 software and GraphPad Prism 8.0 software were employed for data analysis and figure generation, respectively. Results were presented as means ± standard deviation (SD) based on three or nine replicates. Significance among various groups was assessed using Student’s t-test (** p < 0.01, *** p < 0.001, ns: not significant) or one-way analysis of variance (ANOVA) followed by Tukey’s honest significant difference (HSD) test for multiple comparisons (distinguishable letters indicated by p < 0.05).
5. Conclusions
In summary, we used RNA interference to reveal the regulatory role of a specific gene in the seasonal changes of an insect species. We also looked into how the insect’s response to energy-related substances changed. Our results indicated that under certain temperature conditions, a receptor gene was activated in the insect nymphs, leading to the expression of the gene we studied. This gene then played a role in energy storage, chitin synthesis, and adaptation to cold temperatures, ultimately affecting the insect’s survival rates. These findings not only enhance our understanding of gene function but also offer potential insights for environmentally friendly pest management strategies. Further research is required to fully grasp the mechanisms behind these seasonal changes, including exploring the signaling pathways that control the gene of interest and identifying other genes influenced by its expression.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1González-Tokman D. Dáttilo W. Effects of Climate Change on Insects: Physiological, Evolutionary, and Ecological Responses Oxford University Press Oxford, UK 2024
- 2Yang C. Andrew Pospisilik J. Polyphenism—A Window into gene-environment interactions and phenotypic plasticity Front. Genet.20191013210.3389/fgene.2019.0013230863426 PMC 6399471 · doi ↗ · pubmed ↗
- 3Tanaka S. Harano K.-I. Nishide Y. Sugahara R. The mechanism controlling phenotypic plasticity of body color in the desert locust: Some recent progress Curr. Opin. Insect Sci.201617101510.1016/j.cois.2016.05.01127720068 · doi ↗ · pubmed ↗
- 4Richard G. Trionnaire G.L. Danchin E. Sentis A. Epigenetics and insect polyphenism: Mechanisms and climate change impacts Curr. Opin. Insect Sci.20193513814510.1016/j.cois.2019.06.01331557627 · doi ↗ · pubmed ↗
- 5Yuan Y. Wang Y. Ye W. Yuan E. Di J. Chen X. Xing Y. Sun Y. Ge F. Functional evaluation of the insulin/insulin-like growth factor signaling pathway in determination of wing polyphenism in pea aphid Insect Sci.20233081682810.1111/1744-7917.1312136178731 · doi ↗ · pubmed ↗
- 6Fischer R.C. Ölzant S.M. Wanek W. Mayer V. The fate of Corydalis cava elaiosomes within an ant colony of Myrmica rubra: Elaiosomes are preferentially fed to larvae Insectes Sociaux 200552556210.1007/s 00040-004-0773-x · doi ↗
- 7Smýkal V. Pivarči M. Provazník J. BazalováO. Jedlička P. Lukšan O. Horák A. VaněčkováH. BenešV. Fiala I. Complex evolution of insect insulin receptors and homologous decoy receptors, and functional significance of their multiplicity Mol. Biol. Evol.2020371775178910.1093/molbev/msaa 04832101294 PMC 7253209 · doi ↗ · pubmed ↗
- 8Xu H.-J. Xue J. Lu B. Zhang X.-C. Zhuo J.-C. He S.-F. Ma X.-F. Jiang Y.-Q. Fan H.-W. Xu J.-Y. Two insulin receptors determine alternative wing morphs in planthoppers Nature 201551946446710.1038/nature 1428625799997 · doi ↗ · pubmed ↗