Caenorhabditis elegans brc-1 mutation increases the number of COSA-1 foci in him-8 and zim-2 mutants
Takamune T. Saito, Koki Yamamoto, Hirohito Minami, Taiki Tsujiue

TL;DR
This study shows that a mutation in the brc-1 gene increases COSA-1 foci in specific mutants of C. elegans, affecting chromosome pairing and crossover formation.
Contribution
The study reveals a novel role for brc-1 in regulating COSA-1 foci in mutants with pairing defects.
Findings
zim-1 mutants show increased COSA-1 foci compared to expected levels.
brc-1 mutation suppresses the excess COSA-1 foci in zim-1 mutants.
brc-1 mutation increases COSA-1 foci in him-8 and zim-2 mutants during oogenesis.
Abstract
Crossover designation factors such as COSA-1 are concentrated at the specific DNA double-strand break (DSB) sites to promote crossover formation. zim-1 mutants, which show defects in the homologous chromosome pairing of chromosomes II and III, increase the COSA-1 foci/normal bivalent state compared to the expected value. The excess designation was suppressed by an additional mutation in brc-1 in zim-1 mutants. We demonstrated that the number of COSA-1 foci in him-8 and zim-2 mutants, showing defects in the pairing of the X and V chromosomes, respectively, increased compared to the expected value, and brc-1 mutation accelerated the number of COSA-1 foci in oogenesis.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
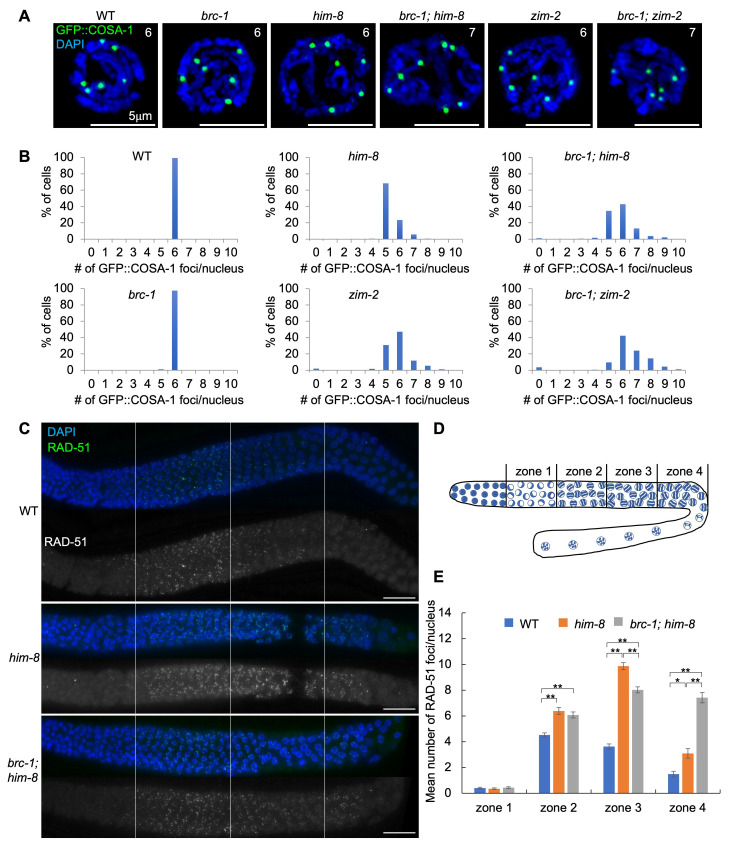 Figure 1
Figure 1|
Strain |
Genotype |
Available from |
|
|
|
CGC |
|
|
|
NBRP |
|
|
|
NBRP |
|
|
|
NBRP, CGC |
|
|
|
Saito lab |
|
|
|
Saito lab |
|
|
|
Villeneuve lab, CGC |
|
|
|
Saito lab |
|
|
|
Saito lab |
|
|
|
Saito lab |
|
|
|
Saito lab |
|
|
|
Saito lab |
- —Japan Society for the Promotion of Science (Japan)https://ror.org/00hhkn466
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsS100 Proteins and Annexins · Protease and Inhibitor Mechanisms · Clusterin in disease pathology
Description
Meiosis is a cell division process that generates haploid gametes from diploid parental cells. Crossover formation is essential for appropriate homologous chromosome segregation of meiosis I. A single crossover event per homologous chromosome pair is observed in wild-type Caenorhabditis elegans . The process of selecting a specific DNA double-strand break (DSB) as a future crossover site is called crossover designation (Kleckner
- et al.* 2003; Gray and Cohen 2016). Crossover designation factors such as COSA-1 , MSH-5 , and ZHP-3 concentrate at the sites of DSBs, which are further repaired by crossover (Kelly
- et al.* 2000; Jantsch
- et al.* 2004; Bhalla
- et al.* 2008; Yokoo
- et al.* 2012). Therefore, six COSA-1 foci are observed in the nucleus at the late pachytene stage in the wild-type (Yokoo
- et al.* 2012).
Homologous chromosomes begin to pair at the pairing center via pairing center-binding proteins ( ZIM-1 , ZIM-2 , ZIM-3 , and HIM-8 ) during the leptotene-zygotene transition (Phillips
- et al.* 2005; Phillips and Dernburg 2006). The homologous pairing of chromosomes II and III is impaired in
zim-1 * mutants (Phillips and Dernburg 2006) . Although * zim-1 * mutants are expected to have four COSA-1 foci per nucleus, an average of 6.12 COSA-1 foci are observed in these mutants (Li
- et al.* 2018). The mutation of
brc-1 * , which is a homolog of human breast cancer gene 1 (BRCA1) (Futreal
- et al.* 1994; Miki
- et al.* 1994; Boulton
- et al.* 2004), suppresses the increase in GFP::COSA-1 foci in
brc-1 ; zim-1 * double mutants (4.3–4.8 COSA-1 foci/nucleus) (Li
- et al.* 2018). In this study, we investigated whether
him-8 * and * zim-2 * mutants, in which the pairing of the X and V chromosomes, respectively, is defective (Phillips
- et al.* 2005; Phillips and Dernburg 2006), show an increase in COSA-1 foci compared to the expected value (5 foci/nucleus), and whether the phenotypes were suppressed by the
brc-1 * mutation.
We quantified the GFP::COSA-1 foci in the late pachytene stage in the wild-type, * him-8 * , * brc-1 , zim-2 * single mutants, and * brc-1 ; him-8 * , * brc-1 ; zim-2 * double mutants by 3D-fluorescent microscopy. As previously shown by many groups, six GFP::COSA-1 foci were observed in the wild-type ( Figure 1A, 1B). If a single crossover event was hypothesized to occur per homologous chromosome pair, then we would expect to observe five GFP::COSA-1 foci/nucleus in * him-8 * mutants with unpaired X chromosomes and in * zim-2 * mutants with unpaired chromosome V. However, in addition to five foci of GFP::COSA-1/nucleus, we observed that 30.5% and 65.4% of cells had more than six foci of GFP::COSA-1 in * him-8 * and * zim-2 * mutants, respectively ( Figure 1A, 1B). This phenotype was similar to that observed in * zim-1 * mutants (Li
- et al.* 2018). These data suggest that the interchromosomal effect causes excess crossover in a normal pair of homologous chromosomes if crossover formation does not occur in one or two sets of unpaired chromosomes during C. elegans oocyte meiosis (Herman and Kari 1989; Carlton
- et al.* 2006; Li
- et al.* 2018). A similar increase in crossover formation between normal chromosome pairs has been observed during Drosophila female meiosis when chromosomal rearrangements (heterozygous inversions or translocations) are present in the cell (Sturtevant 1919; Sturtevant 1921) .
Furthermore, we examined whether the * brc-1 * mutation suppressed the number of COSA-1 foci in a * him-8 * and * zim-2 * backgrounds. Similar to the wild-type, 97.4% of cells showed six GFP::COSA-1 foci in * brc-1 * mutants ( Figure 1A, 1B). GFP::COSA-1 foci were further increased in * brc-1 ; him-8 * and * brc-1 ; zim-2 * double mutants compared to * him-8 * and * zim-2 * single mutants. A total of 62.1% and 86.5% of cells had more than six foci of GFP::COSA-1 in * brc-1 ; him-8 * and * brc-1 ; zim-2 * double mutants, respectively (p<0.00001; Mann-Whitney U test, comparison with each single mutant). These results suggest that BRC-1 suppresses the excess of COSA-1 foci in the * him-8 * and * zim-2 * mutant backgrounds.
One of the major roles of BRC-1 is to stabilize RAD-51 filaments when crossover formation is impaired (Janisiw
- et al.* 2018; Li
- et al.* 2018). RAD-51 levels in pachytene are elevated in
him-8 * and * zim-1 * mutants compared to those in the wild-type (Carlton
- et al.* 2006; Li
- et al.* 2018). Removal of BRC-1 in
zim-1 * mutants results in a “dark zone” of RAD-51 in mid to late pachytene in which the accumulation of RAD-51 foci are suppressed (Li
- et al.* 2018). To examine whether BRC-1 promotes RAD-51 filament stability in
him-8 * mutants, we compared RAD-51 levels in four individual zones from the first cell of the transition zone to the end of late pachytene in the wild-type, * him-8 * , and * brc-1 ; him-8 * double mutants ( Figure 1C, 1D, 1E). Similar to a previous study, the levels of RAD-51 foci were elevated in all pachytene stages (zones 2, 3, and 4) in * him-8 * mutants compared with the wild-type (Carlton
- et al.*
- ( Figure 1C, 1E). The RAD-51 levels were decreased to 81% in mid-pachytene (zone 3) in
brc-1 ; him-8 * double mutants compared to * him-8 * single mutants (p<0.0001); however, obvious RAD-51 dark zone was not observed in * brc-1 ; him-8 * double mutants. Most RAD-51 foci remain in the late pachytene stage (zone 4) in * brc-1 ; him-8 * double mutants compared with * him-8 * single mutants (p<0.0001). These results suggest that the contribution of BRC-1 to RAD-51 filament stability at mid-pachytene in * him-8 * mutants was not as high as that in the * zim-1 * mutant.
We observed an excess number of GFP::COSA-1 foci in the * him-8 * and * zim-2 * single mutants compared with the expected value of the five autosome pairs. These findings are consistent with those of previous studies, which demonstrated that crossover formation increases in autosomes during oogenesis in * him-8 * mutants (Herman and Kari 1989; Carlton
- et al.* 2006). These data suggest that interchromosomal effects occur in
him-8 * and * zim-2 * mutants, similar to * zim-1 * mutants (Li
- et al.* 2018).
The phenotypes of * brc-1 ; him-8 * ( * zim-2 * ) and * brc-1 ; zim-1 * differed during oogenesis. The * brc-1 * mutation suppresses the formation of COSA-1 foci in * zim-1 * mutants (Li
- et al.* 2018), whereas the
brc-1 * mutation enhanced the formation of COSA-1 foci in * him-8 * and * zim-2 * mutants in the present study. A RAD-51 dark zone was observed in * brc-1 ; zim-1 * double mutants (Li
- et al.* 2018), whereas in
brc-1 ; him-8 * double mutants, a slight decrease in RAD-51 levels, but no clear RAD-51 dark zone, was observed. These observations suggest that the function of BRC-1 in the formation of COSA-1 foci and the processing (removal/stabilization) of RAD-51 may be regulated differently according to the number of chromosome pairs involved in oogenesis. Furthermore, COSA-1 foci formation is differentially regulated during oogenesis and male spermatogenesis (Li
- et al.* 2020). The single X chromosome condition in wild-type,
him-8 * mutant , and * brc-1 ; him-8 * double mutant males did not increase the number of COSA-1 foci compared with the hypothesized number of five COSA-1 foci (Li
- et al.* 2020). In contrast to oogenesis,
brc-1 ; zim-1 * males demonstrated enhanced formation of COSA-1 foci compared to * zim-1 * single mutants (Li
- et al.* 2020). Further research is required to explore the sex-specific regulation of crossover designs under pairing defect conditions.
Methods
Fixation and immunostaining:
Worms expressing GFP::COSA-1 (post L4 22–24 h) were dissected using a scalpel. For dissection, 30 µL of 15 mM sodium azide solution was placed on a cover glass, followed by approximately 20 worms. After removing 15 µL of the sodium azide solution, 15 µL of 2% Paraformaldehyde (PFA) was added, mixed to a final concentration of 1% PFA and left for 5 min to fix. After removing 15 µL of the mixture, the sample was sandwiched in a slide glass. The slide was placed at –80°C for 5 min. After removing the cover glass by cracking it with a razor blade, it was fixed again in ice-cold methanol (–20°C) for 1 min. Immunostaining was performed as previously described (Saito
- et al.* 2009). The primary and secondary antibodies used in this study were rabbit anti-RAD-51 antibody ( (Das
- et al.* 2022), 1:3,000) and goat FITC-conjugated anti-rabbit antibody (Jackson ImmunoResearch, 1:200), respectively. The sample was then washed with phosphate-buffered saline with Tween 20 for 5 min for three times, mounted with 8 µL VECTASHIELD with 4',6-diamidino-2-phenylindole (Vector laboratories, Burlingame, California, USA), and the cover glass was shielded by nail polish.
Imaging:
Images were obtained using an all-in-one fluorescence microscope (BZ-X800; KEYENCE, Osaka, Japan) equipped with a Plan Apochromat 100x objective (NA1.45; BZ-PA100; KEYENCE). Approximately 30–40 Z-stack images of germline nuclei were captured at intervals of 0.2 µm. Green fluorescence was detected using a GFP filter (excitation, 470/40 nm; emission, 525/50 nm; dichroism, 495 nm; OP-87763; KEYENCE, Osaka, Japan). The BZ-X analysis software (BZ -H4A) and a 3D application (BZ-H4R) were used for the analysis.
Reagents
Strains:
**: **
After receiving the original strains, TTS65 , TTS185 , TTS186 , TTS272 , and AV630 were outcrossed six times with N2 at the Saito Laboratory.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bhalla N Wynne DJ Jantsch V Dernburg AF 20081024 ZHP-3 acts at crossovers to couple meiotic recombination with synaptonemal complex disassembly and bivalent formation in C. elegans.P Lo S Genet 4101553-7390 e 1000235 e 100023510.1371/journal.pgen.100023518949042 PMC 2567099 · doi ↗ · pubmed ↗
- 2Boulton SJ Martin JS Polanowska J Hill DE Gartner A Vidal M 200416 BRCA 1/BARD 1 orthologs required for DNA repair in Caenorhabditis elegans.Curr Biol 1410960-9822333910.1016/j.cub.2003.11.02914711411 · doi ↗ · pubmed ↗
- 3Carlton PM Farruggio AP Dernburg AF 200623 A link between meiotic prophase progression and crossover control.P Lo S Genet 221553-7390 e 12e 1210.1371/journal.pgen.002001216462941 PMC 1359072 · doi ↗ · pubmed ↗
- 4Futreal PA Liu Q Shattuck-Eidens D Cochran C Harshman K Tavtigian S Bennett LM Haugen-Strano A Swensen J Miki Y 1994107 BRCA 1 mutations in primary breast and ovarian carcinomas.Science 26651820036-807512012210.1126/science.79396307939630 · doi ↗ · pubmed ↗
- 5Jantsch V Pasierbek P Mueller MM Schweizer D Jantsch M Loidl J 200491 Targeted gene knockout reveals a role in meiotic recombination for ZHP-3, a Zip 3-related protein in Caenorhabditis elegans.Mol Cell Biol 24180270-73067998800610.1128/MCB.24.18.7998-8006.200415340062 PMC 515049 · doi ↗ · pubmed ↗
- 6Kelly KO Dernburg AF Stanfield GM Villeneuve AM 2000101 Caenorhabditis elegans msh-5 is required for both normal and radiation-induced meiotic crossing over but not for completion of meiosis.Genetics 15620016-673161763010.1093/genetics/156.2.61711014811 PMC 1461284 · doi ↗ · pubmed ↗
- 7Li Q Saito TT Martinez-Garcia M Deshong AJ Nadarajan S Lawrence KS Checchi PM Colaiacovo MP Engebrecht J 2018111 The tumor suppressor BRCA 1-BARD 1 complex localizes to the synaptonemal complex and regulates recombination under meiotic dysfunction in Caenorhabditis elegans.P Lo S Genet 14111553-7390 e 1007701 e 100770110.1371/journal.pgen.100770130383767 PMC 6211623 · doi ↗ · pubmed ↗
- 8Li Q Hariri S Engebrecht J 2020812 Meiotic Double-Strand Break Processing and Crossover Patterning Are Regulated in a Sex-Specific Manner by BRCA 1-BARD 1 in Caenorhabditis elegans.Genetics 21620016-673135937910.1534/genetics.120.30329232796008 PMC 7536853 · doi ↗ · pubmed ↗