Dual transcriptional inhibition of glutamate and alanine racemase is synergistic in Mycobacterium tuberculosis

TL;DR
This study shows that inhibiting two enzymes together in tuberculosis bacteria has a stronger effect than inhibiting them separately.
Contribution
The study confirms a synergistic interaction between inhibiting two specific enzymes in Mycobacterium tuberculosis using transcriptional repression.
Findings
Simultaneous repression of glutamate and alanine racemase shows synergy in Mycobacterium tuberculosis.
This synergy confirms earlier findings from chemical interaction studies.
Targeting multiple cell wall synthesis enzymes could be a promising strategy for tuberculosis treatment.
Abstract
Synergistic interactions between chemical inhibitors, whilst informative, can be difficult to interpret, as chemical inhibitors can often have multiple targets, many of which can be unknown. Here, using multiplexed transcriptional repression, we have validated that the simultaneous repression of glutamate racemase and alanine racemase has a synergistic interaction in Mycobacterium tuberculosis. This confirms prior observations from chemical interaction studies and highlights the potential of targeting multiple enzymes involved in mycobacterial cell wall synthesis.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
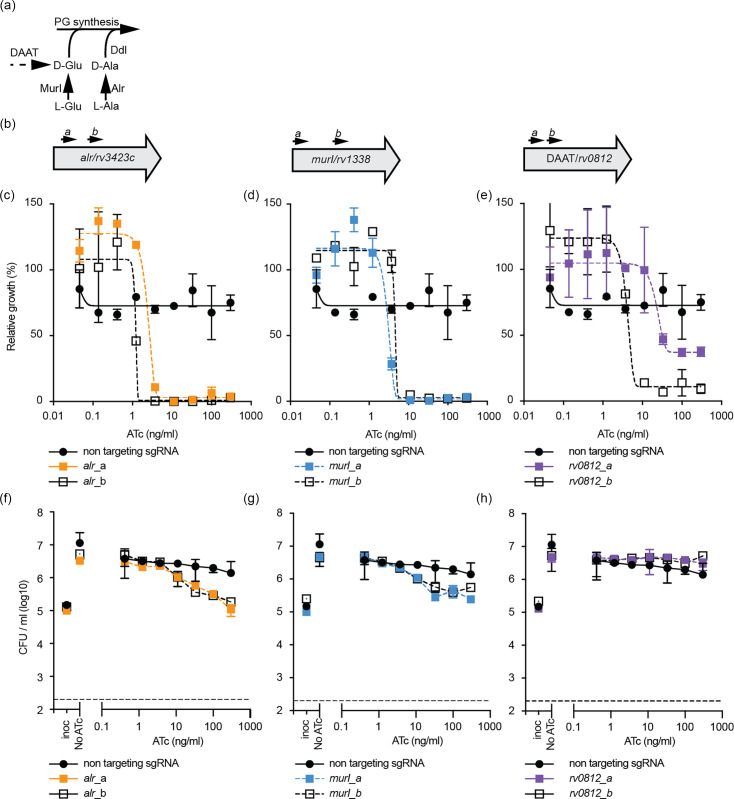 Fig. 1
Fig. 1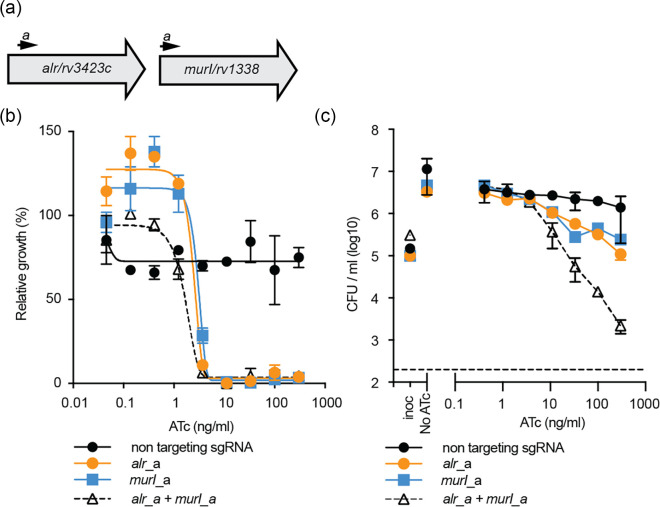 Fig. 2
Fig. 2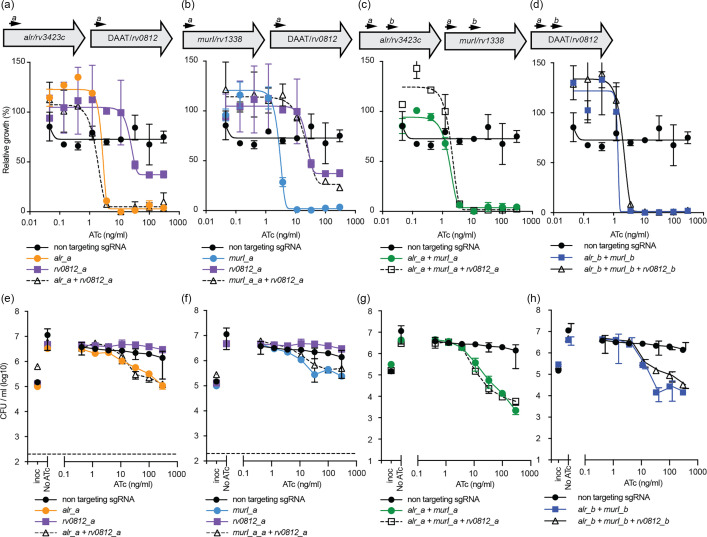 Fig. 3
Fig. 3| gRNA name | Target gene | Target sequence [coding (5′−3′)] | PAM (non-coding 5′−3′, NN…) | PAM score* | sgRNA length (bp) | Fwd oligo (5–3′) (GGGA_) | Oligo rev sequence (AAAC_) |
| alr_a | Rv3423c |
| CCAGAAC | 5 | 21 |
|
|
| alr_b | Rv3423c |
| CCAGAAA | 3 | 22 |
|
|
| murI_a | Rv1338 |
| CCGGAAT | 12 | 21 |
|
|
| murI_b | Rv1338 |
| CGAGCAT | 7 | 21 |
|
|
| rv0812_a | Rv0812 |
| GCAGGAT | 9 | 21 |
|
|
| rv0812_a | Rv0812 |
| CGGGAAG | 4 | 21 |
|
|
- —http://dx.doi.org/10.13039/501100018883 Maurice Wilkins Centre for Molecular Biodiscovery
- —Sir Charles Hercus Health Research Fellowship
- —http://dx.doi.org/10.13039/501100001505 Health Research Council of New Zealand
- —http://dx.doi.org/10.13039/501100018883 Maurice Wilkins Centre for Molecular Biodiscovery
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTuberculosis Research and Epidemiology · RNA and protein synthesis mechanisms · Mycobacterium research and diagnosis
Introduction
Mycobacterium tuberculosis remains a leading cause of death by infectious disease [1]. The mycobacterial cell wall is an interconnected structure of peptidoglycan (PG), arabinogalactan and mycolic acids that generates an impermeable barrier to resist host defences and limits the uptake of many antibiotics [2]. Many clinically utilized antibiotics, including the front-line antibiotic isoniazid, target the mycobacterial cell wall highlighting the essentiality and druggability of this process. Inhibiting cell wall synthesis can also increase cell permeability and synergize with functionally unrelated antibiotics by increasing their uptake [3]. d-Cycloserine (DCS) is a clinically utilized antibiotic that inhibits d-alanine:d-alanine ligase (Ddl) and alanine racemase (Alr), both of which contribute to mycobacterial PG synthesis (Fig. 1a) [48]. DCS synergizes with other drugs that target various components of PG synthesis including β-chloro-d-alanine (BCDA) that targets both glutamate racemase (MurI), an enzyme that contributes d-glutamate to PG synthesis (Fig. 1a) [59], and Alr, which contributes d-alanine [911]. Currently, DCS is used as a second-line agent primarily in the treatment of mycobacterial drug resistance, whilst BCDA, which is not approved for clinical use, has been well studied in mycobacteria [59].
Transcriptional repression of alr, murI and rv0812 in M. tuberculosis. (a) Schematic of enzymatic reactions catalysed by MurI, Alr, Ddl and DAAT (if present). (b) Approximate binding locations of gRNA sequences targeting each gene. a and b correspond to gRNA_a and gRNA_b. Sequences and relevant gRNA information is contained within Table 1. (c–e) Growth of M. tuberculosis expressing either a non-targeting or (c) alr, (d) murI or (e) rv0812 targeting gRNA with differing levels of ATc (anhydrotetracycline). The non-targeting sequence (CGAGACGCATTAATCGTCTCC) is the nucleotide sequence between the bsmB1 cloning sites in pJR965. Cultures were grown in 96-well plates starting from an OD600 of 0.005. OD600 was measured on day 10. Results are the mean±range of two replicates from a representative experiment and expressed relative to a no ATc control. (f–h) Viability of M. tuberculosis expressing either a non-targeting or (f) alr, (g) murI or (h) rv0812 targeting gRNA with differing levels of ATc. Cultures were grown in 96-well plates starting from an OD600 of 0.005. c.f.u. ml−1 was determined after 5 days of growth and is presented as the mean±range of two replicates. Inoc, inoculating c.f.u. ml−1 on day 0.
Whilst these prior results suggest that simultaneous inhibition of Alr and MurI would produce a synergistic drug combination, there is no experimental validation of this interaction as both DCS and BCDA are able to inhibit or interact with multiple targets [4,5, 7, 1215]. Furthermore, the work in M. smegmatis has demonstrated that an aminodeoxychorismate lyase (ADCL) (MSMEG_5795) has significant d-amino acid transaminase moonlighting activity that can, in overexpressing strains, compensate for the loss of MurI by providing an alternative source of d-glutamate [16]. Here, using CRISPR interference (CRISPRi) transcriptional repression, the aim of this study was to (i) validate the synergistic interaction between the inhibition of alr and murI and (ii) determine if the ADCL homologue in M. tuberculosis (rv0812) has sufficient d-amino acid transaminase activity to compensate for the inhibition of murI expression [17]. Specifically, this was achieved by investigating the consequences of transcriptionally inhibiting alr, murI and rv0812 alone and in combination.
Methods
Mycobacterial CRISPRi uses 20–25 nucleotide guide RNA (gRNA) sequences to guide deactivated Cas9 (dCas) to target genes, where dCas9 binds to sterically inhibit transcription [18]. We individually cloned two unique gRNA sequences with complementarity to the non-template sequence of each gene into separate CRISPRi plasmids (i.e. pLJR965) using previously published protocols [1922] (Fig. 1b). Sequence-verified CRISPRi plasmids were electroporated into M. tuberculosis strain, mc^2^6230 (∆panCD, ∆RD1), as previously described. mc^2^6230 is a BSL2 avirulent auxotroph that has been approved for use under BSL2 containment at the University of Otago. The current literature is consistent with the mc^2^6230 strain, supplemented as required for in vitro use, not having effects on cell wall biosynthesis. Reduced phthiocerol dimycocerosate (PDIM) content has been noted in this strain, but this has also been found in other laboratory-propagated strains of M. tuberculosis [23]. Using 96-well plate growth assays, we initially investigated the consequences of repressing alr, murI and rv0812 alone. Assays were performed as previously described [192122]. Briefly, M. tuberculosis strains containing gRNA that targeted genes of interest were inoculated into 96-well plates containing 7H9 supplemented media [i.e. with OADC (0.005 % oleic acid, 0.5 % BSA, 0.2 % dextrose and 0.085 % catalase), 0.05 % tyloxapol (Sigma), 25 µg ml^−1^ pantothenic acid and 25 µg ml^−1^ kanamycin], with a threefold dilution of ATc at a starting OD_600_ of 0.005 in a total volume of 150 µl. Ninety-six-well plates were incubated without shaking at 37 °C for 10 days, and OD_600_ was measured in a Varioskan LUX microplate reader. To determine the effects on cell viability, plates were prepared as mentioned earlier, and the culture was extracted from wells containing relevant ATc concentrations on day 5, diluted and then spotted onto supplemented 7H11. Colonies were counted after 4 weeks at 37 °C.
Results and discussion
Consistent with prior reports of gene essentiality, gRNA targeting alr and murI inhibited growth but had a bacteriostatic phenotype and did not reduce viability compared to the starting inoculum [212425] (Fig. 1c, d, f, g). Both gRNAs targeting rv0812 impaired the growth of M. tuberculosis, with no effect on viability (Fig. 1e, h). This impaired growth is in agreement with prior Tnseq experiments showing that the disruption of rv0812 had a growth defect in M. tuberculosis H37Rv [26].
We next hypothesized that if the simultaneous repression of alr and murI had a synergistic interaction, then dual CRISPRi transcriptional repression would either reduce the amount ATc needed to inhibit bacterial growth or alternatively have a lethal interaction when inhibiting two bacteriostatic genes. To investigate this, we constructed CRISPRi plasmids that expressed two gRNA sequences that individually targeted alr or murI using golden gate cloning as previously described (Fig. 2a) [27]. In CRISPRi phenotypic assays, targeting both alr and murI did not change the concentration of ATc needed to inhibit bacterial growth (Fig. 2b), yet it did produce a bactericidal effect on viability (Fig. 2c). Specifically, the repression of both alr and murI reduces viability by approximately 2-log_10_ c.f.u. ml^−1^ at 300 ng ml^−1^ ATc compared to the individual bacteriostatic gRNAs that did not reduce viability (Fig. 2c). The bactericidal effect of dual alr+murI targeting was concentration dependent, with an approximately 0.5 and 1-log_10_ reduction in c.f.u. ml^−1^ at 33 and 100 ng ml^−1^ ATc (Fig. 2c). In conclusion, the dual repression of alr and murI has a synergistic interaction against bacterial viability in M. tuberculosis.
Transcriptional repression of alr and murI is synergistic in M. tuberculosis. (a) Approximate binding locations of gRNA sequences targeting each gene in dual targeting CRISPRi plasmids. (b) Growth of M. tuberculosis expressing either a non-targeting, alr, murI or alr and murI. Cultures were grown in 96-well plates starting from an OD600 of 0.005 with differing levels of ATc. OD600 was measured on day 10. Results are the mean±range of two replicates from a representative experiment and expressed relative to a no ATc control. (c) Viability of M. tuberculosis expressing either a non-targeting, alr, murI or alr and murI. Cultures were grown in 96-well plates starting from an OD600 of 0.005 with differing levels of ATc. c.f.u. ml−1 was determined after 5 days of growth and is presented as the mean±range of two replicates. Inoc, inoculating c.f.u. ml−1 on day 0.
We hypothesized that if rv0812 was functionally redundant for murI activity in M. tuberculosis, then repression rv0812 would have a synergistic interaction with either (i) repression of alr or (ii) repression of murI or would enhance the synergistic interaction between the dual repression of alr and murI. To investigate this, we constructed CRISPRi plasmids that expressed combinations of gRNA to repress either (i) alr+rv0812, (ii) murI+rv0812 or (iii) alr+murI+rv0812 using golden gate cloning as previously described (Fig. 3a–c) [27]. In CRISPRi phenotypic assays, the simultaneous targeting of rv0812 with alr or murI did not change the concentration of ATc needed to inhibit bacterial growth (Fig. 3a–c). The repression of rv0812 with either alr or murI also maintained the bacteriostatic effect on viability that was similar to the repression of alr or murI alone (Fig. 3d, e). We did note an apparent rescue, by CRISPR inhibition of rv0812, of some of the murI-induced growth defects. However, the repression of rv0812 did not alter the bactericidal effect of the dual repression of alr+murI with the triple- and double-repression strain having similar effects on viability when using either the a or b gRNA (Fig. 3c, f). Prior work in M. smegmatis observed that mutations in the MSMEG_5795 (i.e. rv0812 homologue) promoter were selected and upregulated MSMEG_5975 when a ∆murI mutant was grown for prolonged periods in the absence of d-Glu [16]. It is possible that in our experimental setup, the expression levels of rv0812 in M. tuberculosis are too low and do not allow for rescue of murI transcriptional inhibition. In conclusion, the repression of rv0812 does not augment the effect of inhibiting either alr or murI expression in M. tuberculosis.
Transcriptional repression of rv0812 does not interact with alr or murI repression that is synergistic in M. tuberculosis. (a–d) Approximate binding locations of gRNA sequences targeting each gene in multiplexed CRISPRi plasmids. (a–h) Growth of M. tuberculosis expressing either a non-targeting and (a) alr±rv0812, (b) murI±rv0812 or (c) alr+murI±rv0812. Cultures were grown in 96-well plates starting from an OD600 of 0.005 with differing levels of ATc. OD600 was measured on day 10. Results are the mean±range of two replicates from a representative experiment and expressed relative to a no ATc control. (d–f) Viability of M. tuberculosis expressing either a non-targeting and (d) alr±rv0812, (e) murI±rv0812 or (f) alr+murI±rv0812. Cultures were grown in 96-well plates starting from an OD600 of 0.005 with differing levels of ATc. c.f.u. ml−1 was determined after 5 days of growth and is presented as the mean±range of two replicates. Inoc, inoculating c.f.u.
Here, using CRISPRi transcriptional repression, we have investigated the interactions between murI and alr enzymes that contribute d-glutamate and d-alanine, respectively, to PG biosynthesis in M. tuberculosis. The simultaneous inhibition of both murI and alr had a synergistic interaction with regard to bacterial killing. Whilst we did not observe any reduction in the ATc concentration needed to inhibit growth, our observation of a synergistic killing interaction between murI and alr supports prior results showing an interaction between BCDA and DCS, chemical inhibitors of these respective enzymes [9]. Notably, rv0812 did not appear to be functionally redundant for murI, within the parameters of our experimental paradigm. Given that DCS also targets Ddl, which codes for the essential enzyme Ddl, future work using CRISPR technology to explore synergy with this target would be of interest.
Although our results support the synergistic interaction between murI and alr inhibition, there are distinct differences between transcriptional and chemical inhibition. For example, CRISPRi transcriptional inhibition leads to the removal of the resulting protein and any other co-transcribed genes, whilst chemical inhibition only affects enzyme function leaving the protein largely intact. These differences may confound a direct correlation between our genetic study and future chemical interaction studies on target murI and alr inhibitors. In conclusion, these results highlight the potential of drug combinations that target murI and alr in addition to those that target multiple enzymes within cell wall biosynthesis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Global tuberculosis report 2022 Geneva World Health Organization 2022
- 2Jankute M Cox JAG Harrison J Besra GS Assembly of the mycobacterial cell wall Annu Rev Microbiol 20156940542310.1146/annurev-micro-091014-10412126488279 · doi ↗ · pubmed ↗
- 3Mc Neil MB Chettiar S Awasthi D Parish T Cell wall inhibitors increase the accumulation of rifampicin in Mycobacterium tuberculosis Access Microbiol 20191 e 00000610.1099/acmi.0.00000632974492 PMC 7470358 · doi ↗ · pubmed ↗
- 4Chen J Zhang S Cui P Shi W Zhang W et al Identification of novel mutations associated with cycloserine resistance in Mycobacterium tuberculosis J Antimicrob Chemother 2017723272327610.1093/jac/dkx 31628961957 PMC 5890659 · doi ↗ · pubmed ↗
- 5Prosser GA Rodenburg A Khoury H de Chiara C Howell S et al Glutamate racemase Is the primary target of β-Chloro-d-Alanine in Mycobacterium tuberculosis Antimicrob Agents Chemother 2016606091609910.1128/AAC.01249-1627480853 PMC 5038272 · doi ↗ · pubmed ↗
- 6Prosser GA De Carvalho LPS Metabolomics reveal D-alanine:D-alanine ligase as the target of d-cycloserine in Mycobacterium tuberculosis ACS Med Chem Lett 201341233123710.1021/ml 400349 n 24478820 PMC 3903091 · doi ↗ · pubmed ↗
- 7Halouska S Fenton RJ Zinniel DK Marshall DD Barletta RG et al Metabolomics analysis identifies d-alanine-d-alanine ligase as the primary lethal target of d-cycloserine in mycobacteria J Proteome Res 2014131065107610.1021/pr 401057924303782 PMC 3975674 · doi ↗ · pubmed ↗
- 8Batson S De Chiara C Majce V Lloyd AJ Gobec S et al Inhibition of D-Ala:D-Ala ligase through a phosphorylated form of the antibiotic d-cycloserine Nat Commun 20178193910.1038/s 41467-017-02118-729208891 PMC 5717164 · doi ↗ · pubmed ↗