The complete chloroplast genome of Taraxacum albidum (Asteraceae), a Japanese endemic dandelion
Haruka Mouri, Mizuki Tatsumi, Takako Nishino, Takeshi Suzuki, Tatsuyoshi Morita, Motomi Ito, Takaya Iwasaki

TL;DR
This paper reports the complete chloroplast genome of Taraxacum albidum, a Japanese dandelion, and compares it to related species.
Contribution
The study provides the first complete chloroplast genome sequence for T. albidum and identifies its closest relative.
Findings
The chloroplast genome of T. albidum is 151,451 bp long with a typical quadripartite structure.
It contains 113 unique genes and has a GC content of 37.7%.
Phylogenetic analysis shows T. albidum is closely related to T. mongolicum with nearly identical chloroplast genomes.
Abstract
Taraxacum albidum, a perennial herb of the Asteraceae family, exhibits both tetraploid and pentaploid in Japan. This study sequenced and characterized the complete chloroplast genome of T. albidum, revealing a 151,451 bp sequence with a typical quadripartite structure, comprising one large single-copy (LSC) region of 84,052 bp, one small single-copy (SSC) region of 18,541 bp, and two inverted repeat (IR) regions, IRa and IRb, each 24,429 bp in length. The chloroplast genome, excluding duplicates, contained 113 unique genes, including 79 protein-coding genes, 30 transfer RNA genes, and four ribosomal RNA genes. The GC content of this genome was 37.7%. Phylogenetic analysis revealed that T. albidum is most closely related to T. mongolicum, with the chloroplast genome sequences being nearly identical, differing by only one nucleotide. These findings suggest that the maternal lineage of T.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1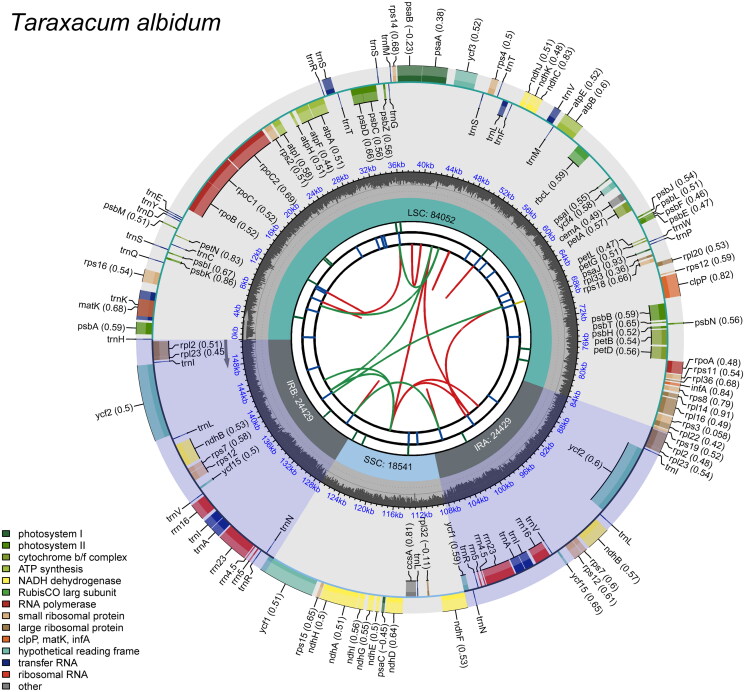 Figure 2
Figure 2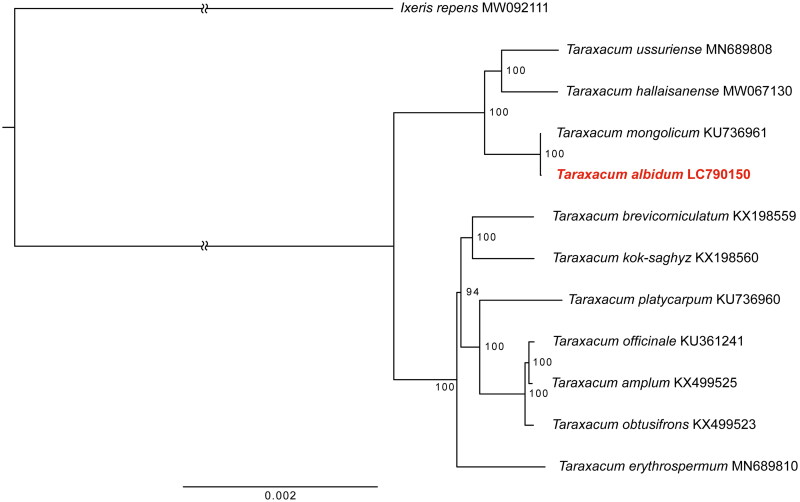 Figure 3
Figure 3- —Sasakawa Scientific Research Grant
- —The Japan Science Society
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Taxonomy and Phylogenetics · Sesquiterpenes and Asteraceae Studies · Plant Diversity and Evolution
Introduction
The genus Taraxacum Weber ex F.H.Wigg. 1780 (Asteraceae), known as dandelion, exhibits a high species diversity, comprising 2516 species (WFO 2023). In Japan, 14 species, one subspecies, and two varieties are distributed (Kadota et al. 2017). Although molecular phylogenetic analysis of this genus Taraxacum has been conducted using intergenic regions of chloroplast DNA and nuclear ITS regions (Wittzell 1999; Kirschner et al. 2015), the evolutionary relationships remain insufficiently resolved due to the complexity of genome composition resulting from multiple hybridizations and genome duplications. Additionally, since only a few Japanese species have been included in the previous studies, the phylogenetic relationships among Japanese dandelions are still unclear. We focused on T. albidum Dahlst. 1907 (Figure 1), an endemic and agamospermous species of the Japanese archipelago (Morita 1995). This species is known to exhibit both tetraploid (2n = 32) and pentaploid (2n = 40). A detailed cytogeographical study surveying 1308 individuals from over 200 localities in Japan found tetraploid individuals are rare and confined to Kyushu, while pentaploid individuals are widespread across the entire range (Sato et al. 2011). Based on the distribution, we presume the individual collected in Tokyo for this study to be pentaploid. The objective of this study is to construct the complete chloroplast genome of T. albidum and elucidate its maternal lineage, presumed to have originated from past hybridization events.
Image of Taraxacum albidum in bloom. This species is a perennial herb with white flowers and oblanceolate to linear-lanceolate leaves that are pinnatilobed or pinnatifid. The lateral lobes are triangular, either entire or irregularly dentate, often dentate between the lobes. The terminal lobe is also triangular and ranges from obtuse to acute at the apex. This photograph was taken by the first author Haruka Mouri on 14 February 2024, at Ochanomizu University, Tokyo, Japan. It is provided here free of any copyright restrictions.
Materials and methods
The plant material of T. albidum was collected at Ochanomizu University, Bunkyo-ku, Tokyo, Japan (latitude 35.7221° N, and longitude 139.7401° E) according to local regulations and with permission. The specimen was deposited in the Makino Herbarium of Tokyo Metropolitan University (Herbarium code: MAK) (https://www.biol.se.tmu.ac.jp/herbarium/) (contact person: Noriaki Murakami, [email protected]) under the specific identifying voucher number MAK-470661. Total genomic DNA was extracted from the dried leaf tissue of T. albidum using the Wizard Genomic DNA Purification Kit from Promega (Madison, WI). Prior to the standard DNA extraction procedure of the kit, we conducted two separate washes using PVP-HEPES buffer (pH 8.0 HEPES buffer, polyvinylpyrrolidone (PVP), l-ascorbic acid, 2-mercaptoethanol) (Setoguchi and Ohba 1995). PVP adsorbs polyphenols, thus inhibiting DNA-polyphenol binding (Echevarría-Machado et al. 2005). The extracted DNA was subcontracted to BGI (16 Dai Fu Street, Tai Po Industrial Estate, Tai Po, New Territories, Hong Kong) for shotgun sequencing with a data volume of 5 GB, NGS platform DNBSEQ, and read length PE 150. Adapter trimming and quality filtering were conducted using the software fastp v.0.20.0 (Chen et al. 2018) with default settings. De novo assembly of the chloroplast genome was conducted using NOVOPlasty v. 4.3.1 (Dierckxsens et al. 2017) (type: chloro, genome range: 120,000–180,000, K-mer: 39). The chloroplast genome sequence of T. mongolicum Hand.-Mazz. 1907 (GenBank accession number KU736961) (Kim, Park, Lee, Woo, et al. 2016) was used as a seed sequence for the NOVOPlasty analysis. To validate the accuracy of the assembly, we mapped trimmed reads to the assembled chloroplast genome and evaluated the depth of coverage using python scripts (Ni et al. 2023). Annotation of the obtained chloroplast genome was carried out using the software GeSeq (Tillich et al. 2017) and CPGAVAS2 (Shi et al. 2019) to identify protein-coding genes, tRNAs, and rRNAs. When discrepancies arose between the annotations provided by GeSeq and CPGAVAS2, manual corrections were made, guided by the annotation results from other species in the genus Taraxacum. The de novo assembly resulted in two patterns of chloroplast genome with full lengths; the difference between the two sequences was whether the SSC sequence is inverted. Both the two chloroplast genome sequences of T. albidum were annotated, and we selected one with homologous aligned sequences with that of T. mongolicum. The CPGView (Liu et al. 2023, http://www.1kmpg.cn/cpgview/) was used to draw the structural characteristics of the chloroplast genome and visualize the intron-containing genes.
To confirm the phylogenetic position of T. albidum, 10 complete chloroplast genome sequences of other Taraxacum species were downloaded from GenBank, and Ixeris repens A.Gray 1858 (Asteraceae) was used as outgroup. Because SSC sequences were inverted in two species (T. obtusifrons Markl. 1940 and T. amplum Markl. 1940), they were inverted and oriented before the phylogenetic analysis. The chloroplast genome sequences were aligned using MAFFT v. 7.490 (Katoh and Standley 2013) with default settings in the software Geneious Prime 2022 (Biomatters Ltd., Auckland, New Zealand). A maximum-likelihood phylogenetic tree was constructed using RAxML v. 8 (Stamatakis 2014) in Geneious Prime. The GTRGAMMA model was used for the nucleotide substitution model, and 1000 bootstrap replicates were carried out.
Results
From the shotgun sequencing, we obtained 20,073,031 raw reads, with 96.15% of them having a Q20 quality score or higher. The assembled complete chloroplast genome of T. albidum, with a GenBank accession number of LC790150, was 151,451 bp in length, had an overall GC content of 37.7%, and an average coverage of ×908.33 (Figure 2 and Supplementary Figure 1). It exhibited a typical quadripartite structure comprising one large single-copy (LSC) region of 84,052 bp, one small single-copy (SSC) region of 18,541 bp, and two inverted repeat (IR) regions, IRa and IRb, each 24,429 bp in length (Figure 2). After excluding duplicates, the chloroplast genome included 113 unique genes: 79 protein-coding genes, 30 transfer RNA genes, and four ribosomal RNA genes. Among these, eight protein-coding genes, seven transfer RNA genes, and all four ribosomal RNA genes were duplicated, with each gene represented in both the IRa and IRb regions. Thirteen protein-coding genes were cis-spliced, with 11 of them (rps16, rpoC1, atpF, petB, petD, rpoA, rpl16, rpl2, ycf2, ndhB, and ndhA) containing a single intron, and two (clpP and ycf3) containing two introns each (Supplementary Figure 2A). Additionally, the placement of exon regions in the trans-splicing gene rps12 was identified (Supplementary Figure 2B). In the constructed phylogenetic tree, T. albidum was most closely related to T. mongolicum, a triploid species distributed from Mongolia to the Korean Peninsula and Japan (Figure 3). The chloroplast genome sequences of T. albidum and T. mongolicum were nearly identical, showing 99.999% similarity with only one nucleotide difference.
Complete chloroplast genome map of Taraxacum albidum. This circular map, generated by CPGView (http://www.1kmpg.cn/cpgview/), illustrates the chloroplast genome’s structure across six concentric tracks. Starting from the center, the first track shows the dispersed repeats, including direct (D) and palindromic (P) repeats, linked by red and green arcs, respectively. The second track depicts long tandem repeats as short blue bars, while the third track visualizes short tandem repeats (or microsatellites) with color-coded bars, indicating different repeat types. The fourth track delineates the genome’s regions: the small single-copy (SSC), large single-copy (LSC), and inverted repeats (IRa and IRb). The fifth track plots the GC content variation across the genome. The outermost track displays genes, color-coded according to function, with gene transcription directions indicated by the orientation: clockwise for inner genes and counterclockwise for outer genes. Codon usage bias, where applicable, is noted in parentheses following gene names. A legend in the bottom left clarifies the functional classification of genes by color.
Maximum-likelihood (ML) phylogenetic tree constructed from complete chloroplast genome sequences of 11 Taraxacum species and Ixeris repens as an outgroup. Bootstrap values, based on 1000 iterations, are shown at each node to indicate confidence levels. The species in this study is highlighted in red. The chloroplast genome sequences analyzed, with their respective GenBank accession numbers, include: Ixeris repens MW092111 (Lee, Yang, Kim, et al. 2021), Taraxacum ussuriense Komarov 1932 MN689808 (Lee, Kim, Woo, et al. 2021), Taraxacum hallaisanense Nakai 1915 MW067130 (Lee, Kim, Kim, et al. 2021), Taraxacum mongolicum KU736961 (Kim, Park, Lee, Woo, et al. 2016), Taraxacum brevicorniculatum Korol. 1940 KX198559 (Zhang et al. 2017), Taraxacum kok-saghyz Rodin 1933 KX198560 (Zhang et al. 2017), Taraxacum platycarpum Dahlst. 1907 KU736960 (Kim, Park, Lee, Woo, et al. 2016), Taraxacum officinale F.H.Wigg. 1780 KU361241 (Kim, Park, Lee, Lee, et al. 2016), Taraxacum amplum KX499525 (Salih et al. 2017), Taraxacum obtusifrons KX499523 (Salih et al. 2017), and Taraxacum erythrospermum Andrz. ex Besser 1822 MN689810 (Lee, Kim, Woo, et al. 2021).
Discussion and conclusions
Since chloroplasts are predominantly maternally inherited in most angiosperms (Hagemann 2004), the phylogenetic tree based on chloroplast genomes provide insights into the maternal lineage. These results suggest that the maternal lineage of T. albidum is likely T. mongolicum. Based on allozyme analysis results, Morita et al. (2021) proposed a hybrid origin of the pentaploid T. albidum from an unreduced gamete of the tetraploid T. albidum and a reduced gamete of any diploid Taraxacum species. This hypothesis aligns with our findings. Combining this hypothesis and our results, we infer that the direct maternal species of the pentaploid T. albidum is the tetraploid T. albidum, and that the maternal species of the tetraploid T. albidum is T. mongolicum. However, our study did not encompass all Taraxacum species within this geographical range, so there might be species more closely related to T. albidum. The continued collection and analysis of chloroplast genome data from various species within this genus will further elucidate the detailed evolutionary history.
Supplementary Material
SupplymentaryFigure_2 600dpi6inch.tif
SupplymentaryFigure_1 600dpi6inch.tif
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chen S, Zhou Y, Chen Y, Gu J. 2018. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 34(17):i 884–i 890. doi:10.1093/bioinformatics/bty 560.30423086 PMC 6129281 · doi ↗ · pubmed ↗
- 2Dierckxsens N, Mardulyn P, Smits G. 2017. NOVO Plasty: de novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 45(4):e 18. doi:10.1093/nar/gkw 955.28204566 PMC 5389512 · doi ↗ · pubmed ↗
- 3Echevarría-Machado I, Sánchez-Cach LA, Hernández-Zepeda C, Rivera-Madrid R, Moreno-Valenzuela OA. 2005. A simple and efficient method for isolation of DNA in high mucilaginous plant tissues. Mol Biotechnol. 31(2):129–135. doi:10.1385/MB:31:2:129.16170213 · doi ↗ · pubmed ↗
- 4Hagemann R. 2004. The sexual inheritance of plant organelles. In: Daniell H, Chase C, editors. Molecular biology and biotechnology of plant organelles: chloroplasts and mitochondria. Dordrecht, Netherlands: Springer; p. 93–113.
- 5Kadota Y, Setoguchi H, Soejima A, Toma T, Nakata M, Morita T, Yonekura K. 2017. Asteraceae. In: Ohashi H, Kadota Y, Murata J, Yonekura K, editors. Wild flowers of Japan. Vol. 5. Tokyo: Heibonsha; p. 198–369.
- 6Katoh K, Standley DM. 2013. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 30(4):772–780. doi:10.1093/molbev/mst 010.23329690 PMC 3603318 · doi ↗ · pubmed ↗
- 7Kim J-K, Park JY, Lee YS, Lee HO, Park H-S, Lee S-C, Kang JH, Lee TJ, Sung SH, Yang T-J. 2016. The complete chloroplast genome sequence of the Taraxacum officinale F.H.Wigg (Asteraceae). Mitochondrial DNA B Resour. 1(1):228–229. doi:10.1080/23802359.2016.1176881.33473461 PMC 7800007 · doi ↗ · pubmed ↗
- 8Kim J-K, Park JY, Lee YS, Woo SM, Park H, Lee S, Kang JH, Lee TJ, Sung SH, Yang T. 2016. The complete chloroplast genomes of two Taraxacum species, T. platycarpum Dahlst. and T. mongolicum Hand.-Mazz. (Asteraceae). Mitochondrial DNA B Resour. 1(1):412–413. doi:10.1080/23802359.2016.1176881.33490398 PMC 7800970 · doi ↗ · pubmed ↗