Sirtulin–Ypk1 regulation axis governs the TOR signaling pathway and fungal pathogenicity in Cryptococcus neoformans
Zhenghua Chai, Yanjian Li, Jing Zhang, Chen Ding, Xiujuan Tong, Zhijie Zhang

TL;DR
This study reveals how acetylation of Ypk1, a key protein in the TOR pathway, affects the pathogenicity of the fungus Cryptococcus neoformans.
Contribution
The discovery of the sirtuin–Ypk1 regulation axis as a novel mechanism governing fungal pathogenicity and TOR signaling.
Findings
Ypk1 is acetylated at lysines 315 and 502, which are in its kinase functional domains.
Deacetylation of Ypk1 is essential for capsule formation, stress response, and pathogenicity.
Sirtuin proteins Dac1 and Dac7 directly interact with Ypk1 to facilitate deacetylation.
Abstract
Cryptococcus neoformans is a life-threatening fungal pathogen that is a causative agent for pulmonary infection and meningoencephalitis in both immunocompetent and immunodeficient individuals. Recent studies have elucidated the important function of the target of rapamycin (TOR) signaling pathway in the modulation of C. neoformans virulence factor production and pathogenicity in animal infection models. Herein, we discovered that Ypk1, a critical component of the TOR signaling pathway, acts as a critical modulator in fungal pathogenicity through post-translational modifications (PTMs). Mass spectrometry analysis revealed that Ypk1 is subject to protein acetylation at lysines 315 and 502, and both sites are located within kinase functional domains. Inhibition of the C. neoformans TOR pathway by rapamycin activates the deacetylation process for Ypk1. The YPK1Q strain, a hyper-acetylation…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
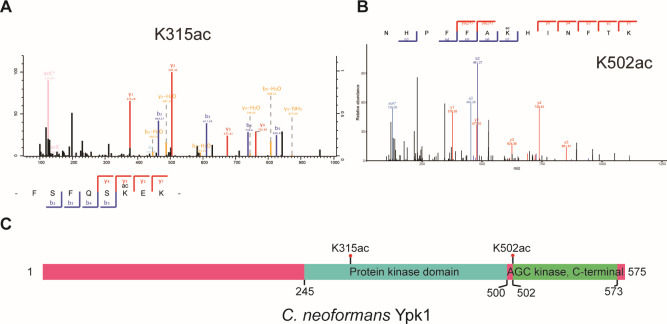 Fig 1
Fig 1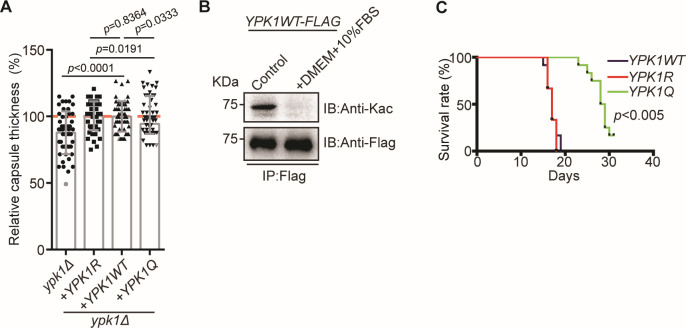 Fig 2
Fig 2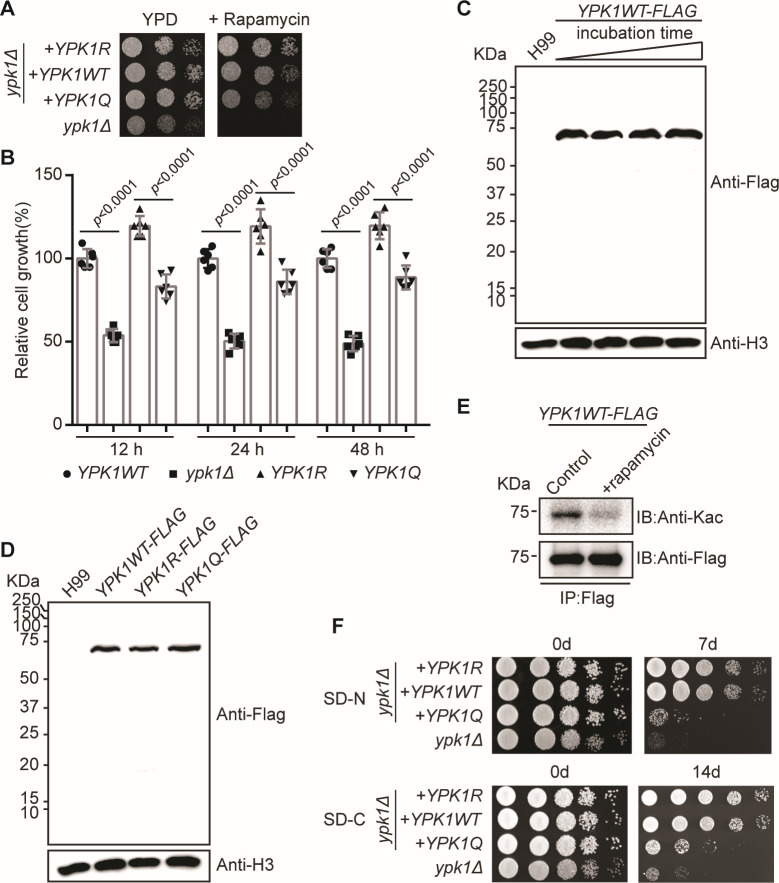 Fig 3
Fig 3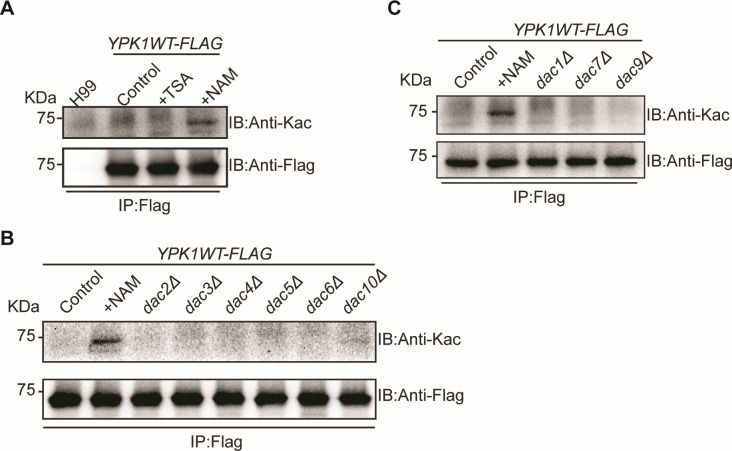 Fig 4
Fig 4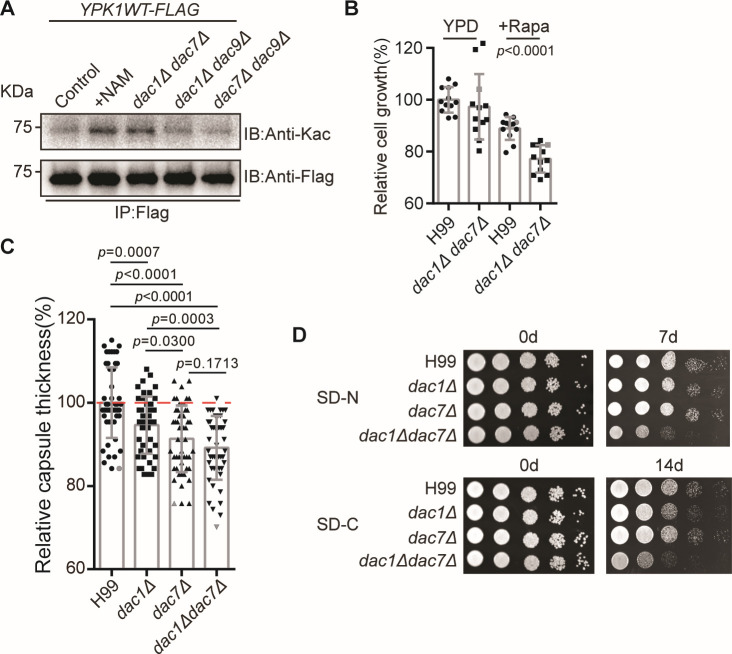 Fig 5
Fig 5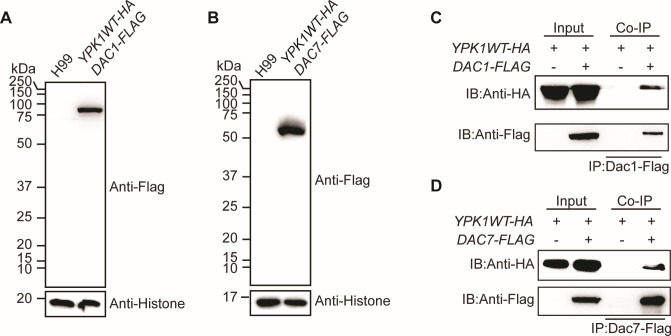 Fig 6
Fig 6- —MOST | National Natural Science Foundation of China (NSFC)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal Infections and Studies · Antifungal resistance and susceptibility · Peptidase Inhibition and Analysis
INTRODUCTION
Cryptococcus neoformans is an airborne opportunistic human fungal pathogen that is the causative agent of cryptococcosis in immunocompromised individuals (1, 2). It is responsible for approximately 135,900 deaths in Sub-Saharan Africa and 15% of global HIV/AIDS-related deaths (3). Studies have extensively shown a correlation between protein post-translational modifications (PTMs) and pathogenicity in human fungal pathogens (4, 5). Protein lysine acetylation is a reversible PTM that is essential for fungal biology. A number of studies have shown that acetylation is critical to the modulation of stress responses, fungal fitness, and invasive growth of Candida albicans and Aspergillus fumigatus in both in vivo and in vitro environments (6–10). However, how acetylation participates in regulating the fungal pathogenicity of C. neoformans remains largely unknown.
Others have shown that acetylation modulation proteins such as acetyltransferases and deacetylases are critical in regulating the transcriptional responses of fungal pathogenicity factors and drug resistance machineries (11–15). Transcriptome analysis following exposure of C. neoformans to deacetylase inhibitors shows significant alterations in the gene expression of virulence factors (16). Furthermore, deacetylases and acetyltransferases, such as Hos1, Hda1, and Gcn5, have been shown to play essential roles in fungal pathogenicity modulations (17–26). Cryptococcus neoformans Gcn5, an acetyltransferase, is involved in regulating gene expression as a response to environmental stress, osmotic stress, and capsule formation (21, 25, 26). Furthermore, the histone deacetylase (HDAC) null mutant strain demonstrated a fluconazole-sensitive cell growth phenotype (27, 28). Recent analysis of the acetylome in C. neoformans revealed that most HDACs play essential functions in modulating C. neoformans virulence factors, including melanin, capsule, and fungal burdens in animals (16). Analyses of two HDACs, Dac2 (Hos2) and Dac4 (Hda1), illustrated that both are responsible for maintaining C. neoformans fitness and animal survival by regulating the GTP-binding domain of Tef1, the elongation factor (16).
Our previous studies have extensively mapped the acetylation sites in hundreds of C. neoformans orthologs of proteins involved in virulence (16). In these orthologs, Ypk1, a serine/threonine protein kinase, is reciprocally regulated by acetylation at K315 and K502. The hyperacetylation of Ypk1 defects modulates fungal fitness in lung and brain tissues. However, how these sites function in modulating the virulence factors remains uninvestigated. To further elucidate the function of Ypk1 acetylation in modulating C. neoformans pathogenicity, we systematically investigated the potential roles of both acetylation sites in Ypk1 protein function. We found that these sites are important modulators in the TOR process, nutrient utilization and fungal virulence. Screening C. neoformans deacetylase mutant strains shows that Ypk1 is a direct client protein for the sirtuins Dac1 (Sir2) and Dac7 (Hst2).
RESULTS
Acetylation modulates Ypk1 function in fungal pathogenicity
We have shown previously that Ypk1 acetylation is a critical part of modulating fungal fitness in lung and brain tissues (16). Mutating acetylation sites K^ac315^ and K^ac502^ to glutamine (Q), which mimics the hyperacetylation of Ypk1, dramatically decreased the colony-forming units in both tissues, indicating that acetylation is critical in regulating Ypk1 protein activity and fungal pathogenicity. While K^315^ was localized within the predicted protein kinase domain (Fig. 1A and C), K^502^ was found at the AGC kinase domain (Fig. 1B and C).
Cryptococcus neoformans Ypk1 is an acetylated protein. (A) Acetylation mass spectrograph of Ypk1 K315. (B) Acetylation mass spectrograph of Ypk1 K502. (C) Scheme of protein domains and acetylation sites of Ypk1. Protein domains were analyzed using InterPro (https://www.ebi.ac.uk/interpro/). Acetylation sites were mapped.
Cryptococcus neoformans capsule formation, used to validate the function of acetylation in Ypk1 mutant strains, showed that capsule thickness was reduced in the ypk1Δ strain compared to the wild-type strain (Fig. 2A). Additionally, a moderate reduction in the capsule structure was detected in the YPK1Q strain (expressing Ypk1^K315Q, K502Q^) compared to the YPK1R strain (expressing Ypk1^K315R, K502R^) (Fig. 2A). The evidence indicates that acetylation of Ypk1 inhibits the development of the capsular structure in C. neoformans, and it is highly likely that the deacetylation process is necessary for this process. Consequently, we cultured the YPK1WT cells in Dulbecco's modified Eagle medium (DMEM) that stimulates capsule formation and subsequently measured the level of acetylation of Ypk1 in this specific growth condition. Our analysis revealed a significant reduction in the overall acetylation levels of Ypk1 when cells were cultivated in DMEM supplemented with 10% fetal bovine serum (FBS). This finding suggests that the process of capsule formation necessitates the deacetylation of Ypk1 (Fig. 2B). Consistent with this point, animal infection experiments demonstrate a prolonged animal survival rate for the YPK1Q strain (Fig. 2C). These data suggest that the acetylation level of Ypk1 plays an important role in modulating functional pathogenicity.
Ypk1 acetylation regulates fungal pathogenicity. (A) Capsule analysis of YPK1 mutants. Capsule thickness, measured as a relative percentages of total capsule thickness. The capsule was induced in Dulbecco’s modified Eagle medium supplemented with 10% fetal calf serum for 2 days. At least 50 cells of each strain were measured. Two-tailed unpaired t-tests were used. (B) Immunoblots, using pan anti-Kac and anti-Flag antibodies, of proteins immunoprecipitated (using FLAG beads) from the YPK1–FLAG strain grown in yeast extract peptone dextrose (YPD) medium or DMEM supplemented with 10% fetal bovine serum (FBS) for 25 minutes. (C) The Kaplan–Meier survival chart of intranasally infected mice (n = 10 per strain).
Deacetylation is required for a Ypk1 response to rapamycin and starvation tolerance
The reciprocal BLAST analysis demonstrated significant homology between C. neoformans Ypk1 and S. cerevisiae Ypk1 (http://fungidb.org/). Studies of S. cerevisiae have shown that Ypk1 is a critical player in the TOR process (29–33). To examine the potential function of C. neoformans Ypk1 and its acetylation regulation in the TOR pathway, we challenged ypk1 mutant strains with the TOR inhibitor, rapamycin. Consistent with the published data, we found that a loss in function of the ypk1Δ strain resulted in a hypersensitive cell growth phenotype when supplementing with rapamycin (Fig. 3A and B). The rapamycin-sensitive phenotype of C. neoformans ypk1Δ strain fully recapitulated that from S. cerevisiae. These data demonstrate that the function of Ypk1 is conserved across fungal species (33). Complementing a copy of the wildtype YPK1 cDNA in the ypk1Δ strain (YPK1WT) rescued the cell growth impairment in rapamycin growth conditions. However, complementing YPK1R (YPK1^K315R; K502R^ mutation) fully rescued the growth defect of the ypk1Δ strain. Interestingly, the YPK1R strain demonstrated enhanced cell growth compared to the YPK1WT strain (Fig. 3A and B). Additionally, complementing the ypk1Δ strain with YPK1Q (YPK1^K315Q; K502Q^ mutation) showed reduced fungal cell growth in the presence of rapamycin, compared to the YPK1WT strain (Fig. 3A and B). Additional quantifiable tests were performed using liquid cell cultures, and the data fully recapitulated that obtained from YPD agar plates (Fig. 3B). These data suggest that C. neoformans Ypk1 functionally resembles that of S. cerevisiae, and acetylation negatively controls Ypk1 activity in C. neoformans.
Ypk1 acetylation is involved in regulating the TOR pathway. (A) Sample YPD agar plates without or with 2 ng/mL rapamycin supplementation after being separately spotted with ypk1Δ/YPK1R, ypk1Δ/YPK1WT, ypk1Δ/YPK1Q, and ypk1Δ strains followed by incubation at 30°C for 2 days. (B) Quantification of YPK1 mutant strains in YPD liquid cultures. Overnight fungal cultures were diluted in fresh YPD, supplemented with 2 ng/mL rapamycin. Cell densities were measured after cell growth at 12 hours, 24 hours, and 48 hours. Two-tailed unpaired t-tests were used. (C) Immunoblots, using anti-Flag and anti-H3 antibodies, of proteins isolated from YPK1 mutants grown in liquid YPD media for 6 hours. (D) Immunoblots quantifying Ypk1–Flag from YPK1WT–FLAG grown in liquid YPD media supplemented with 2 ng/mL rapamycin at 30°C for 0, 5, 15, or 25 minutes. Anti-H3 was used as a loading control. (E) Immunoblots, using pan anti-Kac and anti-Flag antibodies, of proteins immunoprecipitated (using FLAG beads) from the YPK1–FLAG strain grown in YPD liquid media supplemented with or without 2 ng/mL rapamycin at 30°C for 25 minutes. (F) Tolerance of strains to nitrogen or carbon starvation. ypk1Δ/YPK1R, ypk1Δ/YPK1WT, ypk1Δ/YPK1Q, and ypk1Δ strains were grown in liquid YPD for 15 hours and then grown on the SD-N medium and SD-C medium for 7 days and 14 days, respectively. Dilutions were grown on YPD plates for 2 days.
To elucidate the potential function of acetylation in regulating Ypk1 activity, we further investigated whether rapamycin could modulate acetylation levels and the protein levels of Ypk1. To this end, we integrated a YPK1–FLAG plasmid in the ypk1Δ strain. Immunoblotting assays showed that the expression levels of the YPK1WT–FLAG protein in response to elevated concentrations of rapamycin were unchanged (Fig. 3C). Comparable protein levels were detected in YPK1WT–FLAG, YPK1R–FLAG, and YPK1Q–FLAG (Fig. 3D). Upon treating cells with rapamycin, then immunoprecipitating Ypk1WT–FLAG, and monitoring the acetylation levels, we found that exogenous rapamycin decreased the total acetylation levels of Ypk1 (Fig. 3E). Taken together, these results show that rapamycin plays no function in modulating Ypk1 protein levels, but does activate the deacetylation process of Ypk1.
In Saccharomyces cerevisiae, the TORC2–Ypk1 signaling is required for amino acid starvation-induced autophagy (34). Therefore, we analyzed the starvation tolerance of these mutants under nitrogen starvation (SD-N) and carbon starvation (SD-C) conditions in C. neoformans. Our results showed that in the 7-day nitrogen starvation treatment, the ypk1Δ mutant exhibited profound growth defects, as expected, and the YPK1Q strain exhibited growth defects, almost identical to those in the ypk1Δ strain. The same trend of fungal growth was also observed after the 14-day carbon starvation treatment (Fig. 3F). This phenomenon indicates that the cryptococcal Ypk1 has functional similarities to that of S. cerevisiae, and the level of acetylation modification of Ypk1 plays a vital regulatory role in responding to conditions of starvation.
Deacetylation of Ypk1 is dependent on sirtuins
Li et al. have confirmed that there were two main deacetylase families present in Cryptococcus neoformans: HDAC family (histone deacetylase, including Dac2–6, 8, and 11) and sirtuin family (including Dac1, 7, and 9) (16). To elucidate the deacetylation process of Ypk1, we employed two deacetylase inhibitors: TSA, an HDAC inhibitor, and NAM, a sirtuin inhibitor. Cells treated with NAM followed by immunoprecipitation of Ypk1WT-FLAG and immunoblotting assays with Kac showed a dramatic increase in the acetylation level of C. neoformans Ypk1; however, cells treated with TSA maintained low acetylation levels (Fig. 4A). These results suggest that Ypk1 is a client protein for sirtuins rather than HDACs. To precisely identify the corresponding deacetylases, we disrupted nine deacetylase-encoding genes in the YPK1–FLAG strain, but failed to detect any change in acetylation levels of Ypk1–FLAG in these strains (Fig. 4B and C).
The sirtuin deacetylase family regulates Ypk1 acetylation. (A) Immunoblots, using anti-Kac and anti-Flag antibodies, showing YPK1 acetylation after growing the YPK1–FLAG strain in YPD media supplemented with 3 µM TSA or 20 mM NAM at 30°C for 6 hours, isolating the proteins, and then performing Flag immunoprecipitation. The control was the YPK1–FLAG strain grown in unaltered liquid YPD media. (B) Immunoblots, using anti-Kac and anti-Flag antibodies, showing YPK1 acetylation after separately growing the HDAC knockout strains dac2Δ/YPK1–FLAG, dac4Δ/YPK1–FLAG, dac5Δ/YPK1–FLAG, dac6Δ/YPK1–FLAG, and dac10Δ/YPK1–FLAG in YPD media supplemented with 20 mM NAM at 30°C for 6 hours, isolating the proteins, and then performing Flag immunoprecipitation. The controls were the YPK1–FLAG strains grown in unaltered liquid YPD media. (C) Immunoblots, using anti-Kac and anti-Flag antibodies, showing YPK1 acetylation after separately growing the sirtuin knockout strains dac1Δ/YPK1–FLAG, dac7Δ/YPK1–FLAG, and dac9Δ/YPK1–FLAG in YPD media supplemented with 20 mM NAM or 3 µM TSA at 30°C for 6 hours, isolating the proteins, and then performing Flag immunoprecipitation. The controls were the YPK1–FLAG strains grown in unaltered liquid YPD media.
This suggested that multiple deacetylases simultaneously regulate Ypk1 acetylation. To test this, we generated double-knockout mutants of sirtuin genes in the YPK1–FLAG strain and measured the acetylation level of Ypk1. Simultaneously disrupting DAC1 and DAC7 led to enhanced acetylation (Fig. 5A). Consistent with the rapamycin-sensitive growth phenotype of YPK1Q, dac1Δ dac7Δ impairs cell growth in the presence of rapamycin (Fig. 5B). Next, we examined the capsule structure of the knockout strains. These results showed a significant reduction in the thickness of the capsule structure in all three mutant strains, especially in the dac1Δdac7Δ double-knockout strain (Fig. 5C). It is consistent with the phenotype of the YPK1Q strain’s capsule structure (Fig. 2A). Furthermore, we tested the nutrient starvation tolerance of the sirtuin knockout strains. In the conditions of nitrogen and carbon starvation, the dac1Δdac7Δ double-knockout strain showed profound growth defects, and this was recapitulated by the growth phenotype of that from the YPK1Q strain (Fig. 5D). Together, these data demonstrate that both Dac1 and Dac7 have overlapping functions in regulating the deacetylation of Ypk1, which is crucial for adapting the cellular response to nutrition deficiency-induced stress.
Dac1 and Dac7 are critical for Ypk1 acetylation. (A) Immunoblots, using anti-Kac and anti-Flag antibodies, showing YPK1 acetylation after separately growing the sirtuin double-knockout strains dac1Δdac7Δ/YPK1–FLAG, dac7Δdac9Δ/YPK1–FLAG, and dac1Δdac9Δ/YPK1–FLAG in YPD media supplemented with 20 mM NAM at 30°C for 6 hours, isolating the proteins, and then performing immunoprecipitation. The controls were YPK1–FLAG strains grown in unaltered liquid YPD media. (B) Immunoblots showing that cell growth in the dac7Δ/dac9Δ strain is sensitive to rapamycin. Left, H99 and dac7Δ/dac9Δ strains grown in YPD media. Right, the same grown supplemented with 2 ng/mL rapamycin. (C) Capsule analysis of knockout strains. Capsule thickness, measured as a relative percentage of total capsule thickness. The capsule was induced in Dulbecco’s modified Eagle medium supplemented with 10% fetal calf serum for 2 days. At least 50 cells of each strain were measured. Two-tailed unpaired t-tests were used. (D) Tolerance of strains to nitrogen or carbon starvation conditions. H99, dac1Δ, dac7Δ, and dac1Δdac7Δ strains were grown in liquid YPD for 15 hours and then grown on SD-N medium and SD-C medium for 7 days and 14 days, respectively. Dilutions were grown on YPD plates for 2 days.
To determine whether Dac1 or Dac7 physically interact with Ypk1, two strains, YPK1-HA/DAC1–FLAG and YPK1-HA/DAC7–FLAG, were generated, and the expressions of Dac1–FLAG and Dac7–FLAG were confirmed using immunoblotting (Fig. 6A and B). We showed that both of these can be immunoprecipitated with Ypk1-HA (Fig. 6C and D). Collectively, these results show that Ypk1 activity is regulated by acetylation via two independent deacetylases, Dac1 and Dac7.
Dac1 and Dac7 can interact with Ypk1. (A) Immunoblots, using anti-Flag and anti-H3 antibodies, of protein samples isolated from H99 and YPK1–HA/DAC1–FLAG strains. (B) Immunoblots, using anti-Flag and anti-H3 antibodies, of protein samples isolated from H99 and YPK1–HA/DAC7–FLAG strains. (C) Analysis of Ypk1–HA and Dac1–Flag using a coimmunoprecipitation (Co-IP) assay performed on the YPK1–HA/DAC1–FLAG strain. Dac1–Flag was precipitated using Flag beads, and YPK1–HA was detected using anti-HA antibodies. (D) Analysis of Ypk1–HA and Dac7–Flag using a Co-IP assay performed on the YPK1–HA/DAC7–FLAG strain. Dac7–Flag was precipitated using Flag beads, and YPK1–HA was detected using anti-HA antibodies.
DISCUSSION
The TOR pathway, first identified in S. cerevisiae, is a highly conserved process across taxa, playing essential functions in the regulation of a range of biological processes including autophagy, endocytosis, amino acid acquisition, and protein translation (34–43). While one copy of TOR was identified in the genomes of many organisms, two paralogs, Tor1 and Tor2, have been found in S. cerevisiae (44). They form two independent TOR complexes: TOR complex 1 (TORC1) and TOR complex 2 (TORC2) (44). The S. cerevisiae TORC2 is signally transduced via its downstream protein kinase Ypk1, which is functionally associated with sphingolipid homeostasis, mitochondrial respiration, plasma membrane integrity, and many other cellular processes (29, 30, 32, 33, 45). Two homologs of TOR were identified in the human fungal pathogen C. neoformans: Tor1 and Tlk1 (46). Molecular gene manipulation of TOR1 has shown that C. neoformans TOR1 is an essential gene, and the encoding protein product regulates numerous important biological responses (47). However, as a downstream target of Tor1, Ypk1 is indispensable for cell growth in C. neoformans and is required when growing in the presence of rapamycin (48). The C. neoformans Ypk1 has been shown to be involved in drug resistance, and its function is critical for fungal pathogenicity in a mouse infection model when an intravenous infection route is used (48).
Studies of S. cerevisiae have shown that Ypk1 functionally depends on its PTM, phosphorylation. Early studies showed that Pkh1 and Pkh2 phosphorylate Thr 504 on Ypk1, that TORC2 phosphorylates Ser 644 and Thr 662, and that Fpk1 and Fpk2 phosphorylate Ser 51 and Ser 71, respectively (33, 49). More recently, the C-terminus of Ypk1 has been shown to provide four additional TORC2-dependent phosphorylation sites: Ser 653, Ser 671, Ser 672, and Ser 678 (33). In C. neoformans, two phosphorylation sites that were presumptively identified are Thr402, essential for Ypk1 function in fluconazole resistance, and Ser 562, indispensable for Ypk1 function (48). Further evaluation of the protein modification of C. neoformans Ypk1 and its functions in regulating fungal pathogenicity and drug resistance would require in-depth mass spectrometry.
In our previous study, we utilized acetylation mass spectrometry to decipher the novel post-translational modification functions of Ypk1 (16). The results showed that the null ypk1 mutant produced an avirulence phenotype and the hyperacetylation of Ypk1, YPK1Q strain, showed a dramatic reduction in fungal burdens in lung and brain tissues (16).These results suggest that acetylation negatively regulates Ypk1 activity, thus hampering the protein function in modulating fungal pathogenicity in the host.
Interestingly, the YPK1R mutant consistently demonstrates superior growth compared to the YPK1WT strain in both their spotting and liquid culture assays. The primary factor responsible for this is the acetylation state of the wild-type Ypk1 protein. The Ypk1R and Ypk1Q mutagenesis approaches produce Ypk1 proteins with acetylation levels of 0% and 100%, respectively. The acetylation level of the wild-type Ypk1 protein varies between the deacetylation and complete acetylation status.
Our findings indicate that the YPK1R strain displays enhanced growth in different stressful situations, suggesting that deacetylation of Ypk1 is a beneficial mechanism for C. neoformans to cope with environmental stress. Nevertheless, the mechanism by which acetylation of Ypk1 regulates its functionality remains unknown. There are two potential scenarios. Considering that both acetylation sites are located inside the kinase domain, it is likely that acetylation has a significant role in the activity of the kinase. This, in turn, contributes to the regulation of phosphorylation for downstream targets of Ypk1. Furthermore, considering that Ypk1 undergoes both phosphorylation and acetylation, it is possible that these two changes play a role in a complex cross-talk networking mechanism, where one modification greatly influences the other. In order to gain a deeper understanding of the relationship between these two alterations, it is necessary to conduct a comprehensive examination of Ypk1 phosphorylation and its corresponding target proteins by the utilization of mass spectrometry.
The addition of rapamycin reduced the overall acetylation level of Ypk1. This indicates that Cryptococcus maintains a balance between inhibiting the TOR signaling transduction pathway and entering the deacetylation process of Ypk1, which is regulated by the sirtuin family. We have demonstrated that the deacetylation of Ypk1 is dependent on both Dac1 and Dac7, which are members of the sirtuin family of proteins, through a protein–protein interaction. The Ypk1 protein activity is associated with stress tolerance under nutritional deprivation conditions, as supported by the rapamycin treatment results and the published data on S. cerevisiae.
In conclusion, this study demonstrates a new PTM process of Ypk1, an essential player in the TOR process. Acetylation is critical for Ypk1 protein function in fungal virulence production and the pathogenicity machinery, and this study shows that not only is this a negative regulator for Ypk1, but also that deacetylation proteins are required for revealing the Ypk1 protein function. Given the essential function of Ypk1 in regulating the TOR process and fungal pathogenicity, this work enlightens the development of a novel drug target for fungal infection therapy.
MATERIALS AND METHODS
Strain and media
Cryptococcus neoformans wild-type strain H99 and mutant strains were routinely grown in liquid YPD media or YPD agar (1% yeast extract, 2% peptone, 2% dextrose, with or without 2% agar). To conduct the rapamycin test, rapamycin was added to liquid YPD media or YPD agar, reaching a final concentration of 2 ng/mL. To conduct an acetylation analysis, a deacetylase inhibitor, either trichostatin A (TSA) or nicotinamide (NAM) was added, reaching a final concentration of 3 µM or 20 mM, respectively. Using nitrogen-starvation medium [SD-N; 0.17% yeast nitrogen base (without amino acids and ammonium sulfate) and 2% glucose] and carbon-starvation medium [SD-C; 0.17% yeast nitrogen base (without amino acids and ammonium sulfate), 0.5% ammonium sulfate, 0.5% casamino acids, 0.002% tryptophan, 0.002% adenine, and 0.002% uracil) for nitrogen and carbon starvation treatments, respectively (50). Nourseothricin (100 µg/mL) and neomycin (G418) (200 µg/mL) were used to generate mutants. Dulbecco’s modified Eagle medium supplemented with 10% fetal calf serum (FCS) was used for capsule formation.
Strain generation
C. neoformans mutants were generated using the H99 strain and biolistic transformation (51). The R and Q mutants of YPK1WT were provided by Ding’s Lab (16). To generate the YPK1–FLAG dac1Δ dac7Δ strain, DAC1 and DAC7 were sequentially disrupted in the YPK1–FLAG strain. The upstream DNA sequences of DAC1 and DAC7 were amplified using the primer pairs 2,383/2,384 and 2,407/2,408, respectively. The downstream DNA sequences of DAC1 and DAC7 were amplified using the primer pairs 2,385/2,386 and 2,409/2,410, respectively. Overlapping PCRs were performed to join the upstream sequences, selected markers, and downstream sequences and to generate the integrated knockout cassettes.
The strains YPK1–HA DAC1–FLAG and YPK1HADAC7–FLAG were generated and used for co-immunoprecipitation assays. The YPK1–HA strain was first generated using the plasmid pHYG (containing a hygromycin resistance marker under the control of the Cryptococcus GPD promoter). The cDNA sequence of YPK1 was amplified using the primers 3,767 and 3,768, introducing the restriction sites NotI and StuI, respectively. The digested PCR fragment was then cloned into the pHYG plasmid, and the resulting plasmid was transformed into the H99 strain. Expression of Ypk1HA was confirmed by immunoblotting with the anti-HA antibody. The cDNA sequences of DAC1 and DAC7 were amplified using the primer pairs 3,667/3,510 and 3,670/3,671, respectively. Overlapping PCRs were performed to generate upstream-TEF1p–DAC1–4XFLAG–NAT-downstream and upstream-TEF1p–DAC7–4XFLAG–NAT-downstream, which were introduced into the YPK1HA strain. The protein expressions of Dac1–Flag and Dac7–Flag were confirmed using anti-Flag antibody.
Starvation survival assays
Overnight cultures of C. neoformans were harvested, washed three times with sterile PBS, and resuspended in 5 mL of SD-N or SD-C to a final concentration of 1 × 10^7^ cells/mL. The cultures were incubated with shaking at 30°C and 200 rpm for 7 days to induce nitrogen starvation and 14 days to induce carbon starvation. Subsequently, serially diluted cell suspensions at a starting concentration of 1 × 10^7^ cells/mL were spotted on YPD agar plates. Plates were then incubated at 30°C for 2 days to determine the survival rate under starvation conditions.
Animal infection
Six- to 8-week-old female mice (C57BL/6) were purchased from Changsheng Biotech (Liaoning, China) and used for infection experiments. Separately, each C. neoformans strain was suspended (10^5^ fungal cells in 50-µL PBS buffer) and intranasally infected in ten mice (52). Mouse care and infection took place at the College of Life and Health Sciences of Northeastern University under an alternating 12-hour light–dark cycle and ad libitum access to food and water. Infected mice were weighed 10 days after infection and then the morbidity was monitored twice daily. Mice were sacrificed at the end of the experiment.
Capsule formation
Capsule release experiments were performed as described elsewhere (53). Briefly, cells were grown overnight in YPD medium and then washed three times with PBS. The cells were resuspended in Dulbecco’s modified Eagle medium supplemented with 10% fetal calf serum and then incubated for 3 days at 37°C in 5% CO_2_. To observe capsule size, a Leica optical microscope (DM 2500) was used to capture images of cells (generally hundreds) stained with India ink. The size of the surrounding capsule was measured. The data were normalized using the thickness of the YPK1WT strain (in Fig. 2A) and H99 (in Fig. 5C) as the standard.
Cell growth curves
In this study, 96-well microtitre plates were used to assess the growth of cells in liquid culture medium. Overnight YPD cultures were washed three times with phosphate-buffered saline (PBS) and diluted to an optical density of 0.02 at 600 nm in fresh YPD, supplemented with 2 ng/mL rapamycin. Following this, 200 µL of the resulting cell suspension was carefully dispensed into individual wells of a 96-well plate. The well plate was subjected to incubation at a temperature of 30°C for either 12, 24, or 48 hours. Subsequently, optical density measurements at a wavelength of 600 nm were obtained using a Synergy HTX microplate reader manufactured by BioTek. The growth of the relevant strain was standardized by normalizing it to the well without rapamycin treatment. Six or twelve biological replicates were conducted for each strain. The data were graphed utilizing GraphPad Prism software. Two-tailed unpaired t-tests were used.
Protein immunoprecipitation and immunoblotting analysis
Protein immunoprecipitation and immunoblotting analysis were performed as described elsewhere (16). Briefly, protein samples were isolated using a Mini-Beadbeater-16 (BioSpec) and lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 0.1% NP-40, pH 7.5) supplemented with 1X protease inhibitor cocktail (CWBio), 40 mM PMSF, 3 µM TSA, and 20 mM NAM. Anti-FLAG M2 magnetic beads (Sigma) were used in the protein IP. For detection of protein acetylation, after incubating the protein samples with Anti-FLAG M2 magnetic beads for 4 hours, we washed them four times with TBS (50 mM Tris-HCl, 150 mM NaCl, 1% Triton X-100, pH 7.4) supplemented with 3 µM TSA and 20 mM NAM. Then, the bound proteins were extracted into the protein loading buffer at 95°C for 5 min, followed by Western blotting. Monoclonal and polyclonal Kac (1:2500, PTM Bio) was used to detect the level of protein acetylation. Protein–protein interactions were detected after washing with TBS buffer without Triton X-100. The bound proteins were extracted into the protein loading buffer at 95°C for 5 min, followed by Western blotting. Anti-M2 Flag (1:5000, Abcam), anti-HA (1:5000, Abcam), and anti-Histone H3 (1:5000, Cell Signaling Technology) were used for detection of the target protein. The signal was captured using a ChemiDoc XRS+ (Bio-Rad).
Mass spectrometry
Mass spectrometry was performed as described elsewhere (16). Overnight cultures of H99 were diluted in fresh YPD media and incubated at 30°C or 37°C for 6 hours. Cndac2Δ and Cndac4Δ cells were cultured at 30°C, and cells in the exponential phase were used. The protein was extracted using acetylome lysis buffer (8 M urea, 1% Triton X-100, 10 mM dithiothreitol, 1% protease inhibitor cocktail, 3 µM TSA, 50 mM NAM, and 2 mM EDTA), then it was treated with dithiothreitol (5 mM) and iodoacetamide (11 mM), diluted, and digested with trypsin at 37°C overnight. The resulting digested protein samples were desalted and vacuum-dried, then the peptides were resuspended in 0.5 M TEAB, and 6-plex TMT labeling was performed. High-performance liquid chromatography fractionation was performed, and the enriched acetylated peptides were isolated by immunoaffinity precipitation. The label-free peptides and TMT-labeled peptides were analyzed using LC–MS/MS, and the collected MS/MS data were processed using the MaxQuant search engine (version 1.5.2.8).
Statistics and reproducibility
The log-rank (Mantel–Cox) statistical test was used to analyze these animal survival tests. Paired-sample t tests were used in two-sample comparisons. Actual *P-*values were used consistently for representation.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Giles SS, Dagenais TRT, Botts MR, Keller NP, Hull CM. 2009. Elucidating the pathogenesis of spores from the human fungal pathogen Cryptococcus neoformans. Infect Immun 77:3491–3500. doi:10.1128/IAI.00334-0919451235 PMC 2715683 · doi ↗ · pubmed ↗
- 2Velagapudi R, Hsueh Y-P, Geunes-Boyer S, Wright JR, Heitman J. 2009. Spores as infectious propagules of Cryptococcus neoformans. Infect Immun 77:4345–4355. doi:10.1128/IAI.00542-0919620339 PMC 2747963 · doi ↗ · pubmed ↗
- 3Rajasingham R, Smith RM, Park BJ, Jarvis JN, Govender NP, Chiller TM, Denning DW, Loyse A, Boulware DR. 2017. Global burden of disease of HIV-associated cryptococcal meningitis: an updated analysis. Lancet Infect Dis 17:873–881. doi:10.1016/S 1473-3099(17)30243-828483415 PMC 5818156 · doi ↗ · pubmed ↗
- 4Leach MD, Brown AJP. 2012. Posttranslational modifications of proteins in the pathobiology of medically relevant fungi. Eukaryot Cell 11:98–108. doi:10.1128/EC.05238-1122158711 PMC 3272899 · doi ↗ · pubmed ↗
- 5Retanal C, Ball B, Geddes-Mc Alister J. 2021. Post-translational modifications drive success and failure of fungal-host interactions. J Fungi (Basel) 7:124. doi:10.3390/jof 702012433572187 PMC 7914884 · doi ↗ · pubmed ↗
- 6Wurtele H, Tsao S, Lépine G, Mullick A, Tremblay J, Drogaris P, Lee E-H, Thibault P, Verreault A, Raymond M. 2010. Modulation of histone H 3 lysine 56 acetylation as an antifungal therapeutic strategy. Nat Med 16:774–780. doi:10.1038/nm.217520601951 PMC 4108442 · doi ↗ · pubmed ↗
- 7Lu Y, Su C, Liu HP. 2012. A GATA transcription factor recruits Hda 1 in response to reduced Tor 1 signaling to establish a hyphal chromatin state in Candida albicans. P Lo S Pathog 8:e 1002663. doi:10.1371/journal.ppat.100266322536157 PMC 3334898 · doi ↗ · pubmed ↗
- 8Lee I, Oh JH, Shwab EK, Dagenais TRT, Andes D, Keller NP. 2009. Hda A, a class 2 histone deacetylase of Aspergillus fumigatus, affects germination and secondary metabolite production. Fungal Genet Biol 46:782–790. doi:10.1016/j.fgb.2009.06.00719563902 PMC 2755195 · doi ↗ · pubmed ↗