Dose-Dependent Prophylactic Efficacy of Filarial Antigens Glutathione-S-Transferase and Abundant Larval Transcript-2 against Brugia malayi Challenge in Mastomys
Mohini Rambhau Nakhale, Priyanka Bhoj, Namdev Togre, Vishal Khatri, Lalit Batra, Udaikumar Padigel, Kalyan Goswami

TL;DR
This study tests how well different doses of two filarial antigens protect Mastomys against Brugia malayi infection, finding that a specific dose of one antigen with an adjuvant provides strong protection.
Contribution
The study identifies the optimal dose of rBmALT-2 combined with alum for maximal prophylactic efficacy against B. malayi infection in Mastomys.
Findings
A 50 μg dose of rBmALT-2 with alum induced 79–81% killing of L3 larvae in vivo and in vitro.
The combination of rBmALT-2 and rWbGST with alum induced 63–68% larval killing.
Vaccinated Mastomys showed elevated IL-10, IL-4, and IFN-γ levels, indicating Th1 and Th2 immune responses.
Abstract
To identify the most effective dose of filarial rBmALT-2 and rWbGST alone or in combination against B. malayi infection in vitro and in vivo. Mastomys (n = 5–7/group) received intramuscular (i.m.) injection with three different doses (25, 50, and 100 μg) of rBmALT-2 or rWbGST, either alone or in combination with alum as the adjuvant. Protective immunity was studied by in vivo and in vitro cytotoxicity assay. To evaluate the cellular immune response, splenocyte proliferation and cytokine profile were assessed. Serological results revealed a substantial (p < 0.005) induction of IgG1, IgG2a, and IgG3 responses in vaccinated Mastomys. Mastomys immunized with 50 μg rBmALT-2 + alum induced 79–81% killing against the L3 larvae challenge in vivo and in vitro ADCC assay (p < 0.005); whereas rWbGST + alum alone or in combination with rBmALT-2 + alum induced 63–68% killing (p < 0.005) in vivo…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
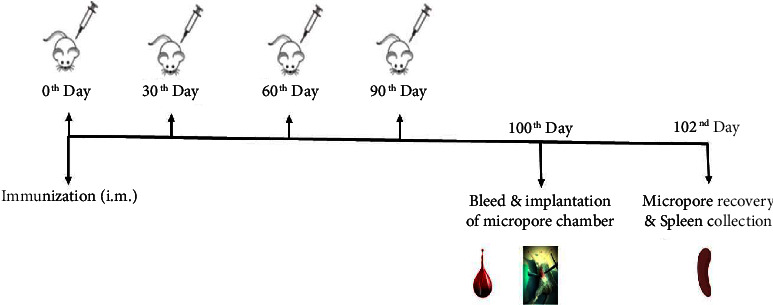 Figure 1
Figure 1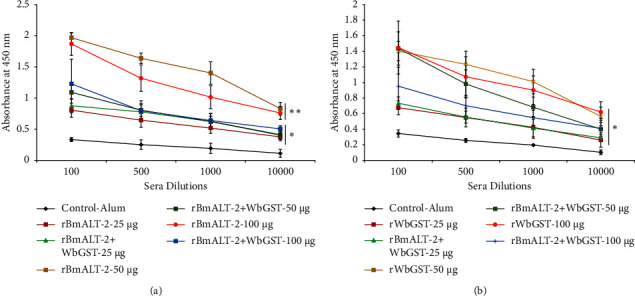 Figure 2
Figure 2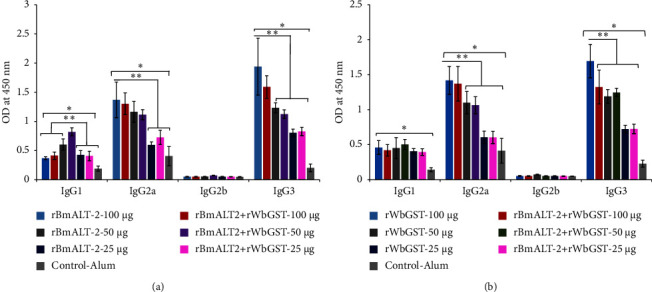 Figure 3
Figure 3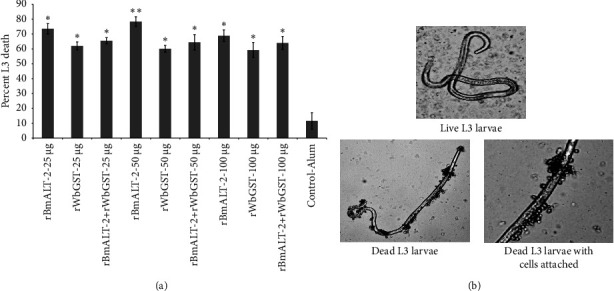 Figure 4
Figure 4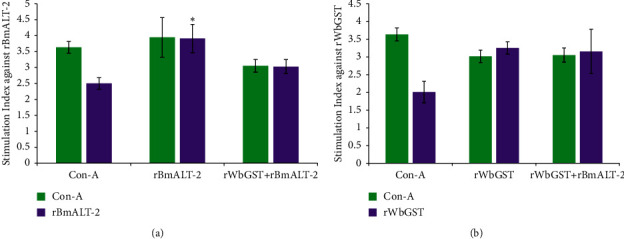 Figure 5
Figure 5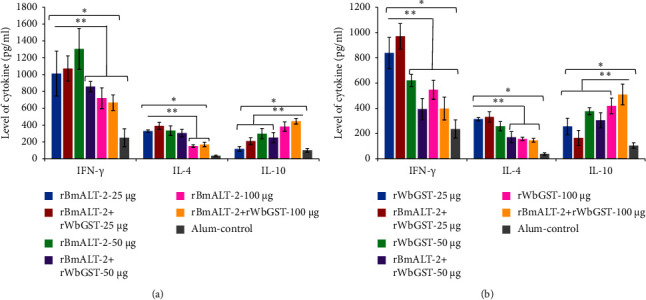 Figure 6
Figure 6- —Department of Biotechnology , Ministry of Science and Technology
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic Diseases Research and Treatment · Insect symbiosis and bacterial influences · Parasites and Host Interactions
1. Introduction
Parasitic worms, specifically Wuchereria bancrofti, Brugia malayi, and B. timori, cause lymphatic filariasis (LF), influencing roughly 863 million individuals worldwide [1]. Mass drug administration (MDA) is one of the strategies used by the World Health Organization (WHO) in the Global Programme to Eliminate Lymphatic Filariasis, which aims to eradicate the disease by 2030 [1]. Since consistent MDA renders parasites vulnerable to developing drug resistance, several treatment modes should be considered. Prophylactic vaccination is one such approach that may aid in LF control and elimination worldwide [2]. Moreover, LF re-emergence in several countries has highlighted the importance of developing a prophylactic vaccine for LF [3–5].
Two filarial antigens, B. malayi abundant larval transcript-2 (BmALT-2) and W. bancrofti glutathione-S-transferase (WbGST), have been proposed as promising vaccine candidates [6, 7]. The ALT-2 protein stimulates high protective immune responses against B. malayi [7–11]. The filariae express these proteins at the late second stage larvae2 (L2) and peak in the third stage larvae3 (L3) [12], playing a pivotal role in transmission and infectivity [8, 13]. The ALT-2 protein has no mammalian homologue [7, 10], and it is unique among host components and has been shown to have an immunomodulatory function in the primary host immune response [14, 15]. Studies suggest that ALT proteins from B. malayi are involved in activating Th2 immune responses that can aid in the survival of parasites [13]. GSTs are one of the crucial enzymes that efficiently inhibit the host's oxidative free radicals [7, 16]. GSTs, primary phase II detoxification enzymes, and antioxidants are crucial for parasite survival in the host [7–9, 13, 17]. Previous vaccination studies in Jirds and Mastomys using rBmALT-2 or rWbGST antigens reported substantial protection against filariasis [7, 10, 18].
This study aimed to evaluate the effective dose of both antigens (rBmALT-2 and rWbGST) either as a single or combination vaccine in Mastomys coucha.
2. Materials and Methods
2.1. Animals and Filarial Parasites
All experiments involving male M. coucha (males, 6–8 weeks) were carried out according to guidelines described by the National Institutes of Health (NIH) and Office of Laboratory Animal Welfare (OLAW) USA [19, 20] and Institutional Animal Ethics Committee (IAEC) of Mahatma Gandhi Institute of Medical Sciences (MGIMS), Sevagram, Maharashtra, India (MGIMS/IAEC/July/8/2014). Baermann's method was employed to obtain B. malayi infective L3 as described [21, 22] and used for further experimental procedures.
2.2. Protein Expression and Purification
The rBmALT-2 and rWbGST proteins were expressed as previously described [8]. Briefly, the bmalt-2 and wbgst genes in the pRSET-A vector were expressed in the BL21 (DE3) pLysS expression host. Isopropyl β-D-1-thiogalactopyranoside of 1 mM (IPTG; Merck Millipore, Bangalore, India) was used to induce these six-histidine-tagged recombinant proteins and further purified by nickel-charged sepharose column (Thermo Fisher Scientific, Mumbai, India). Limulus amebocyte lysate (LAL) chromogenic quantification kit (Thermo Fisher Scientific, Mumbai, India) was used to detect and eliminate endotoxins from purified protein preparations. Protein expression and purity were determined by 12% SDS-PAGE and western blotting with anti-6XHis antibodies (Qiagen, New Delhi, India).
2.3. Immunization, Challenge, and Sera Collection of Mastomys
Mastomys were divided into ten groups (5–7/group). Vaccinated groups received intramuscular (i.m.) injections of 25, 50, or 100 μg/dose of the respective antigens plus alum as adjuvant four times at an interval of 4 weeks as follows: Groups 1, 2, and 3: Mastomys immunized with alum plus rBmALT-2-25 μg, rBmALT-2-50 μg, and rBmALT-2-100 μg, respectively; Groups 4, 5, and 6: Mastomys immunized with alum plus rWbGST-25 μg, rWbGST-50 μg, and rWbGST-100 μg, respectively; Groups 7, 8, and 9: Mastomys immunized with alum plus rBmALT-2 + rWbGST-25, 50, and 100 μg (1 : 1 concentration), respectively; and Group 10: Mastomys administered with alum adjuvant alone (control group). Ten days after the last immunization sera were collected by intraorbital bleeding to be used for serum analysis for antibody titers. Vaccine-induced protection, cytotoxicity, were assessed by inserting a plexiglass diffusion chamber containing 20 B. malayi L3 (Millipore Sigma, Bedford, MA) into Mastomys peritoneal cavity as previously mentioned [10]. After 48 hr, the chambers were retrieved, and their contents were microscopically examined to assess larval killing and cell adhesion. The results were expressed as a percentage cytotoxicity value (Figure 1).
2.4. Splenocyte Proliferation and Cytokine Expression in Culture Supernatant
Mastomys were euthanized by an overdose of ketamine, and spleens were aseptically removed at day 102 (Figure 1), minced, and washed thrice using RPMI-1640 medium, along with 10% heat-inactivated FBS fetal calf serum (FBS) and 1×antibiotic/antimycotic solution (Sigma-Aldrich, Mumbai, India). Single-cell suspensions were prepared (2 × 10^6^ cells/well in 200 μL RPMI media); stimulated with 1 μg/well of the respective antigen (rBmALT-2 or rWbGST), 1 μg/well of positive control concanavalin A (ConA; Sigma-Aldrich, Mumbai, India) and negative control (unstimulated cells) were plated in triplicate in cell culture plates (96-well plate) and incubated for 48 hr (37°C in 5% CO_2_). The MTS assay kit (Promega, New Delhi, India) was used to measure cell proliferation, and the stimulation index (SI) was calculated as previously described [11].
For cytokine analysis, similar sets of single cell suspension (2 × 10^6^ cells/mL/well) were prepared and plated in cell culture plates (24-well plate), which were further stimulated with 10 μg/well of the respective antigens or 2 μg/well of ConA (positive control) and incubated for 72 h (37°C in 5% CO_2_). The cytokine ELISA kits (Invitrogen Bioservices, Bengaluru, India) were used to assess the amounts of IL-4, IL-10, and IFN-γ in the culture supernatants after incubation following the manufacturer's instructions.
2.5. Antigen-Specific IgG Titer and Isotype in the Sera of Vaccinated Mastomys
An indirect ELISA was used to quantify the amounts of rBmALT-2- or rWbGST-specific total IgG antibodies in the sera of Mastomys [11]. The serum samples collected on day 100 (Figure 1) were diluted at dilutions of 1 : 100, 1 : 500, 1 : 1000, and 1 : 10000 and tested for the presence of IgG antibodies using a goat anti-mouse HRP-conjugated secondary antibody diluted 1 : 10000. (Thermo Fisher Scientific, Mumbai, India). TMB substrate was used to develop the reaction, and adding 2M H_2_SO_4_ stopped the color development. A spectrophotometer (Biotek, New Delhi) was used to measure the absorbance at 450 nm.
The concentrations of antigen-specific isotypes (such as IgG1, IgG2a, IgG2b, and IgG3) against rBmALT-2 or rWbGST were measured in the blood samples using an indirect ELISA method, as explained in a previous study [11], with the use of the specific HRP-labeled antibodies for each isotype.
2.6. In Vitro Cytotoxicity Experiment Using B. malayi L3
To assess the cytotoxicity of antigen-specific protective antibodies against B. malayi L3, an in vitro cytotoxicity assay (antibody-dependent cellular assay; ADCC) was used as previously described [7]. Briefly, peritoneal exudate cells (PEC; 2 × 10^5^ cells/well) collected from nonimmunized Mastomys were incubated with 10–12 B. malayi L3 and 50 μL pooled sera samples from immunized Mastomys into 96-well cell culture plates (Thermo Fisher Scientific, Mumbai, India) for 48 hr at 37°C in 5% CO_2_. After 48 hr incubation, the viability of L3 was checked, and percentage cytotoxicity was calculated as previously described [7].
2.7. Statistical Assessment
The statistical studies were conducted using SPSS V21.0 software (IBM, SPSS Inc., India). Survival data were examined using one-way analysis of variance (ANOVA) with a Bonferroni post hoc test. p values <0.05 were considered significant. Sera and cytokine were analyzed by the Kruskal–Wallis test followed by Bonferroni post-test multiple comparisons, p values <0.05 and < 0.005 were considered significant.
3. Results
3.1. All Immunized Mastomys Had a High Antigen-Specific IgG Antibody Titer
All immunized groups showed high antigen-specific IgG antibody titers after the last dose of the vaccination cycle (Figure 2). The Mastomys immunized with 50 or 100 μg rBmALT-2 generated a significant (p < 0.005) IgG antibody titer compared with the Mastomys immunized with 25 μg rBmALT-2 alone and all formulations of rBmALT-2 + rWbGST (Figure 2(a)). Mastomys immunized with 25, 50, or 100 μg rWbGST alone and in combination with rBmALT-2 generated a substantially higher (p < 0.005) IgG antibody titer than control Mastomys (Figure 2(b)).
3.2. All Immunized Mastomys Showed Different Antigen-Specific IgG Isotype Levels
The serum (1 : 100 diluted) isotype profile showed that all immunized Mastomys had significantly (p < 0.005) higher IgG1, IgG2a, and IgG3 isotype levels than the control Mastomys (Figure 3). There was a progressive increase in IgG2a and IgG3 levels with the increasing dose of test antigens (Figure 3). The Mastomys immunized with 50 μg rBmALT-2 alone and in combination with rWbGST showed significantly higher IgG1 levels (p < 0.005) compared with Mastomys immunized with 25 or 100 μg rBmALT-2 alone or the combined rBmALT-2 + rWbGST immunizations (Figure 3(a)). In contrast, immunization of Mastomys with a 100 μg dose of rBmALT-2, rWbGST or the combination rBmALT-2 + rWbGST with alum showed significantly higher IgG2a and IgG3 levels (p < 0.005) compared with the 25 or 50 μg antigen-immunized group (Figure 3).
3.3. ADCC Assay Showed Larvicidal Activity by Antigen-Specific Antibodies in the Sera of Immunized Mastomys
The serum of immunized Mastomys, with different doses, 25, 50, or 100 μg rBmALT-2, rWbGST or their combination, promoted PEC adherence to the L3. All sera of the immunized groups showed significantly higher ADCC than that of the control group; 50 μg rBmALT-2 induced the highest protection (Table 1). Furthermore, regardless of the antigen dose, only Mastomys sera immunized with rBmALT-2 had the highest activity against L3 (p < 0.005) compared to other test antigens.
3.4. Diffusion Chamber Assay Showed Significant Prophylactic Efficacy of Antigens
All immunized groups induced significantly higher (p < 0.005) larval death compared with the control group (Figure 4(a)). When compared with the control group, Mastomys immunized with 25 μg rBmALT-2, rWbGST, and their combination induced 75%, 64%, and 67% protection (p < 0.005), respectively. While 50 μg rBmALT-2 provided higher protection with 79%; p < 0.005 compared with 50 μg rWbGST or their combination (Figure 4(a)). Cells were found to be attached to the surface of many recovered dead L3 (Figure 4(b)).
3.5. Immunized Mastomys Exhibited Increased Cellular Immune Response and Antigen-Recall Response
The splenocytes collected from rBmALT-2-immunized animals showed a significantly (p < 0.05) higher proliferative response compared with those from the control group with a stimulation index of SI) 3.91 ± 0.44 vs. 2.50 ± 0.17 when measured by the MTS assay (Figure 5(a)). Splenocytes collected from rWbGST immunized animals showed higher SI compared with those from the control group 3.26 ± 0.17 vs. 2.01 ± 0.30 when measured by MTS assay (Figure 5(b)). The splenocytes from Mastomys immunized with rBmALT-2 + rWbGST responded to both rBmALT-2 SI 3.02 ± 0.22 and rWbGST SI 3.15 ± 0.62, indicating a strong cellular immune response against these antigens in Mastomys vaccinated with rBmALT-2 + rWbGST (Figure 5).
The restimulated splenocytes of Mastomys immunized with 25 or 50 μg rBmALT-2 alone or in combination with rWbGST significantly released increased (p < 0.005) levels of IFN-γ and IL-4 compared with Mastomys treated with 100 μg rBmALT-2 or in combination with rWbGST. Mastomys treated with 100 μg rBmALT-2 alone or in combination with rWbGST released significantly increased IL-10 levels (p < 0.005) compared with other rBmALT-2 only treated groups (Figure 6(a)). Mastomys treated with 25 μg rWbGST alone or in combination with rBmALT-2 showed significantly elevated (p < 0.005) levels of IFN-γ and IL-4 compared with other rWbGST-only treated groups (Figure 6(b)). In contrast, Mastomys treated with 50 or 100 μg of rWbGST alone or in combination with rBmALT-2 displays increased (p < 0.005) IL-10 levels compared with the control group (Figure 6(b)).
4. Discussion
The present work focuses on two well-documented and potent vaccine candidates: rBmALT-2 and rWbGST. Previous studies showed that immunization with these proteins individually can induce significant (p < 0.05) protection against filariasis in Jirds and Mastomys [6, 7, 18]. Antigen immunogenicity is influenced not only by its physical and biological activities but also by its dose [23]. Several findings have suggested that a modest amount of immunogen may produce cell-mediated immunity (CMI), whereas high doses may elicit humoral immune responses [24–26]. However, some research groups indicate that the type and dose of immunogen utilized mostly regulates the immune response [27, 28]. Hence, this study used purified rBmALT-2 and rWbGST at different doses, 25, 50, and 100 μg, since administering large doses may induce a systemic immune response [29].
Using an in vitro and in vivo, ADCC and diffusion chamber, approach for this investigation, we found higher protective efficacy with 50 μg rBmALT-2 than with 25 and 100 μg rBmALT-2, all doses of rWbGST or the combinations of rBmALT-2 + rWbGST when alum was used as an adjuvant against B. malayi L3. Combination of these two antigens induced approximately 64–68% protection against B. malayi L3 in vitro [2, 10, 30, 31]. In our study, rBmALT-2 induced the highest protective efficacy (69–80% in vitro) regardless of the dose administered.
ADCC is thought to be an essential immunological mechanism in animals and is primarily responsible for circulating parasite disappearance [32–34]. In vitro, cytotoxicity against microfilaria and infective larvae is caused by antibody and complement-mediated effector ADCC pathways [35]. In our in vitro study, antisera produced against these two antigens (BmALT-2 or WbGST) elicited cytotoxicity against L3 within 48 hr. 50 μg of anti-rBmALT-2 (82% protection) serum increased PEC's adherence to L3 as compared to anti-rWbGST (65% protection) or anti-rBmALT-2 + rWbGST (69% protection) serum. Similarly, 25 or 100 μg anti-rBmALT-2 (69–75% protection) serum showed higher cell adhesion than anti-rWbGST (64–65% cytotoxicity) or anti-rBmALT-2 + rWbGST (67-68% protection) serum.
The in vivo diffusion chamber approach, which is performed in a limited physiological environment, has been utilized to investigate parasitic survival, development, and the host effector mechanism. Some experiments using B. malayi have been used to show comparable results between in vitro and in vivo assays [11, 36]. Macrophage participation in cytotoxicity has been verified against different filarial parasites, such as Dipitelonema setariosum adults [37] and the L3 of Litomosoides sigmodontis [38]. In the current study, we discovered that the in vivo diffusion chamber cytotoxicity results were consistent with those obtained from the in vitro ADCC test. The in vivo, diffusion chamber experiment showed 78%, 59%, and 65% protective efficacy with 50 μg rBmALT-2, rWbGST, and their combination, respectively. Similarly, Mastomys immunized with 25 or 100 μg rBmALT-2 showed higher larval killing (approximately 68–73%) than rWbGST or the combination rBmALT-2 + rWbGST (61–65%). Mastomys immunized intramuscularly with 50 μg rBmALT-2 showed better protective efficacy than other immunogens, both in vitro and in vivo. Recombinant BmALT-2 is one of the filarial antigens that can provide substantial protection against L3 [8, 36, 39, 40]. Although the exact mechanism of rBmALT-2 protein is unknown, studies indicate that it may be crucial for host immunomodulation [38, 41].
Notably, the rBmALT-2 + rWbGST combination showed higher and lower protective efficacy (65%) than rWbGST alone (61%) and rBmALT-2 alone (73%) in vivo. This study does not rule out the potential that an instantaneous association among these two antigens (rBmALT-2 and rWbGST) may reduce rBmALT-2 immunogenicity. rWbGST may have concealed the antigenic epitopes on rBmALT-2, reducing rBmALT-2 efficacy in conjugation vaccination [30]. A study on malaria examined the protective potential of two recombinant Plasmodium yoelli merozoite surface proteins (MSPs), namely, PyMSP-8 and PyMSP-142, which has shown a similar result. When mice were treated with a combination of these antigens, the immunogenicity of PyMSP-142 decreased [42]. In a filarial study in Jirds, immunization by conjugating rBmTGA and rBmALT-2 showed merely 47% protection against B. malayi L3 [30]. Therefore, selecting antigens for a successful combination is essential, as not every combination yields a positive outcome.
The T-cell response, typically associated with elevated antibody levels, was investigated using IgG isotypes and cytokine profiles. The result indicated that Mastomys immunized with 50 or 100 μg rWbGST or rBmALT-2 or combination of both antigens (rBmALT-2 + rWbGST) had significant levels of IgG1, IgG2a, and IgG3 antibodies, indicating Th1 and Th2-type reaction. IgG1 and IgG2a isotypes in mice are involved in complement fixation and binding to protein antigens, while IgG3 is involved in carbohydrate-containing epitope identification [36, 41]. Mouse immune complexes with IgG2a and IgG3 can attach to FcγRI and initiate receptor-mediated responses [36, 43]. These isotypes also contribute to the immune response of vaccines and the in vitro reactions (ADCC) against invading pathogens.
Antigen-specific cytokine profiles of Mastomys vaccinated with either BmALT-2 or WbGST or conjugation of both antigens had increased IL-4, IL-10, and IFN-γ levels, indicating a Th1 and Th2 immune response. Moreover, 50 μg BmALT-2 only and in combination with rWbGST induced high IL-10 secretion. IgG1 and IgG3 antibody responses are stimulated explicitly by Th2 cell-produced IL-4 and IL-10, whereas increased IFN-γ reflects the Th1 immune response [44, 45]. Our findings indicate that vaccine candidates and their combination elicited mixed Th1/Th2 immune responses. Some research findings suggest that Th2 immune responses contribute to intestinal helminth resistance [45], although applying these findings to tissue-dwelling nematodes, like LF [45], is challenging. The Th1-type immune response is essential to stimulate immune protection against filarial infection [45]. Intramuscular immunization with 50 μg rBmALT-2 with alum against filarial infection is associated with elevated IFN-γ, IL-4, and IL-10 levels and protective Th1 and Th2 immune responses.
In conclusion, Mastomys immunized four times at a 4-week interval with 50 μg rBmALT-2 plus alum as the adjuvant showed an enhanced total IgG antibody titer levels. The isotype profile showed increases in IgG2a, followed by IgG3 and IgG1 levels, demonstrating a mixed Th1 and Th2 response. Immunization with rBmALT-2 plus alum also significantly increased levels of IFN-γ, IL-4, and IL-10 in the spleen cells stimulated ex vivo., indicating a mixed Th1 and Th2 response. Moreover, the immunization presented higher protection of approximately 80% against L3 larvae in in vitro ADCC and in vivo diffusion chamber assay. Therefore, 50 μg/intramuscular dose of rBmALT-2 plus alum as the adjuvant can be used as an effective immunization dose for human LF to explore further preclinical and mechanistic studies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mondiale de la SantéGlobal programme to eliminate lymphatic filariasis: progress report, 2018–programme mondial pour l’élimination de la filariose lymphatique: rapport de situation Weekly Epidemiological Record 2019944457470
- 2Anand S. B. Kodumudi K. N. Reddy M. V. Kaliraj P. A combination of two Brugia malayi filarial vaccine candidate antigens (Bm ALT-2 and Bm VAH) enhances immune responses and protection in jirds Journal of Helminthology 201185444245210.1017/s 0022149 x 100007992-s 2.0-8065514472121208482 · doi ↗ · pubmed ↗
- 3Sunish I. P. Munirathinam A. Kalimuthu M. Kumar V. A. Tyagi B. K. Persistence of lymphatic filarial infection in the paediatric population of rural community, after six rounds of annual mass drug administrations Journal of Tropical Pediatrics 201460324524810.1093/tropej/fmt 1012-s 2.0-8490182170324343822 · doi ↗ · pubmed ↗
- 4Bhattacharjee J. Mass drugs administration in India-a failure story Epidemiology: Open Access 20166310.4172/2161-1165.1000252 · doi ↗
- 5Dyson L. Stolk W. A. Farrell S. H. Hollingsworth T. D. Measuring and modelling the effects of systematic non-adherence to mass drug administration Epidemics 201718566610.1016/j.epidem.2017.02.0022-s 2.0-8501456814528279457 PMC 5340860 · doi ↗ · pubmed ↗
- 6Gupta S. Bhandari Y. P. Reddy M. V. Harinath B. C. Rathaur S. Setaria cervi: immunoprophylactic potential of glutathione-S-transferase against filarial parasite brugia malayi Experimental Parasitology 2005109425225510.1016/j.exppara.2004.12.0032-s 2.0-1474430347915755423 · doi ↗ · pubmed ↗
- 7Anandharaman V. Dakshinamoorthy G. Gnanasekar M. Reddy M. V. R. Kalyanasundaram R. Evaluation of wuchereria bancrofti GST as a vaccine candidate for lymphatic filariasis P Lo S Neglected Tropical Diseases 200936 p. e 45710.1371/journal.pntd.00004572-s 2.0-68349121689 PMC 268597819513102 · doi ↗ · pubmed ↗
- 8Gregory W. F. Atmadja A. K. Allen J. E. Maizels R. M. The abundant larval transcript-1 and-2 genes of brugia malayi encode stage-specific candidate vaccine antigens for filariasis Infection and Immunity 20006874174417910.1128/iai.68.7.4174-4179.20002-s 2.0-003391710310858234 PMC 101719 · doi ↗ · pubmed ↗