Adeno-associated virus-mediated intraprostatic suppression of MIR375 inhibits tumor progression in the TRAMP mouse model of prostate cancer
Xianyanling Yi, Jin Li, Zeyu Han, Tianyi Zhang, Dazhou Liao, Jia You, Jianzhong Ai

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
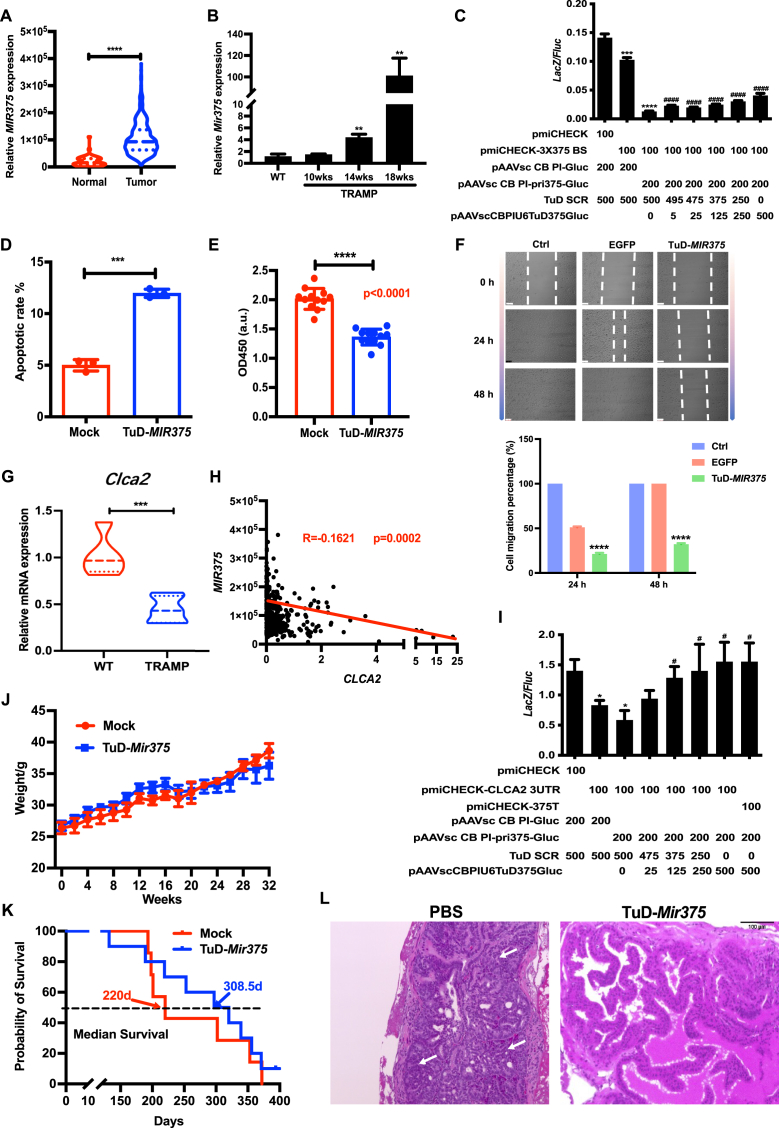 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsComparative Literary Analysis and Criticism · Literary and Philosophical Studies
The treatment of prostate cancer (PCa) needs to be improved.1 Micro-RNAs (miRNAs) are a subtype of non-coding, single-stranded RNAs that influence cellular survival and death by modulating mRNAs. Among these miRNAs, MIR375 has a critical role in the regulation of tumorigenesis2 and holds promise as a novel therapeutic target for future PCa treatment. Recombinant adeno-associated virus (rAAV) exhibits non-pathogenicity, low-grade inflammation, and robust and long-lasting expressions of target genes. We have previously described the rAAV9 as a valid vector to transfer target miRNAs and genes,3 so rAAV9 could be used as a vector to deliver MIR375 into the mouse prostate and PCa cells.
We initially obtained relative expression of MIR375 in PCa from The Cancer Genome Atlas (TCGA) database (http://cancergenome.nih. gov/), and the expression level of MIR375 was compared between tumor and normal tissues. As depicted in Figure 1A, tumor tissues exhibited a significantly higher expression level of MIR375 than normal tissues. Subsequently, transgenic adenocarcinoma mouse prostate (TRAMP) models of different ages were sacrificed, and prostates were harvested. Total RNA was extracted and Mir375 expression was analyzed via quantitative real-time PCR. A remarkable increase in the expression level of Mir375 in TRAMP mice in comparison to wild-type mice. Furthermore, it was observed that the expression level of Mir375 in TRAMP mice increased with age (Fig. 1B). Then, to assay for MIR375 activity, we constructed a dual-gene reporter vector. The data showed that LacZ/Fluc reporter activity was significantly down-regulated by MIR375 (Fig. S1A). It indicated that MIR375 could exert its regulatory function via the vector pAAVsc CB PI- miR-375-Gluc. Moreover, tough decoy (TuD) antisense RNA was used, an efficient and specific inhibitor of miRNA, and the LacZ/Fluc reporter activity was restored (Fig. 1C). Thus, the expression of MIR375 was reduced using TuD in subsequent experiments. The effect of MIR375 on PC3 cell apoptosis was examined with flow cytometry, and the inhibition of MIR375 led to a significant increase in apoptotic cell death, encompassing both early and late apoptosis stages (Fig. 1D; Fig. S1B). Cell viability was detected by CCK8 assay. According to Figure 1E, the proliferation of PC3 cells was attenuated upon MIR375 inhibition. Furthermore, the scratch experiment showed that the scratch width in the TuD-MIR375 group was wider than that of the control group after 24 h and 48 h (Fig. 1F). These findings collectively suggested that MIR375 promotes the proliferation and migration of PC3 cells.Figure 1MIR375 could inhibit apoptosis and promote proliferation and migration of prostate cancer cells via its target gene CLCA2, and rAAV9-TuD*-Mir375* improves the survival in TRAMP mice. (A) The expression level of MIR375 in prostate cancer analyzed by the TCGA database. **(B)**Mir375 expression level evaluated by quantitative real-time PCR in WT and TRAMP mice at different ages. (C) TuD RNA was cloned into a lentiviral vector to inhibit MIR375 function. The pmiCHECK plasmid DNA was used as the positive control, and the sequence of the triple MIR375 binding site was inserted into the downstream region of the LacZ coding sequence (CDS) of pmiCHECK-3 × 375 BS. (D) Annexin V-FITC/PI staining analysis of PC3 cell apoptosis. (E) CCK8 assay was applied to detect cell viability. (F) Migration ability was assessed by wound-healing assay. **(G)**Clca2 expression between TRAMP mice and control mice. (H) Correlation analysis of MIR375 and CLCA2 expression. (I) TuD SCR partly reversed the LacZ/Fluc reporter activity after transfection with 3′UTR of CLCA2. (J) TRAMP mice were injected with rAAV9-TuD-Mir375 or PBS, and mouse weight was recorded every two weeks. (K) The median survival time of mice injected with rAAV9-TuD-Mir375 or PBS. (L) Prostates were assessed by hematoxylin and eosin staining. The white arrow reveals tumorigenesis. TRAMP, transgenic adenocarcinoma mouse prostate; WT, wild-type; TuD SCR, tough decoy-short consensus repeat; PBS, phosphate-buffered saline. ^∗^P < 0.05, ^∗∗^P < 0.01, ∗∗∗P < 0.001, ^#^P < 0.05, ^####^P < 0.0001; ∗and # indicate comparisons with the first column and the third, respectively.Fig. 1
Furthermore, we delved into the expression levels of CLCA2 utilizing TCGA datasets, revealing a noteworthy decrease in CLCA2 expression in PCa compared with normal tissues (Fig. S1C). Likewise, Clca 2 expression level was significantly lower in TRAMP mice via performing quantitative real-time PCR of RNA extracted from TRAMP mice and control mice (Fig. 1G). Subsequently, we explored the potential relationship between MIR375 and CLCA2 in PCa. The MIR375 expression was significantly negatively correlated with that of CLCA2 by analyzing the expression data of the TCGA dataset (Fig. 1H). Additionally, CLCA2 expression was increased through the inhibition of MIR375 (Fig. S1D). Figure 1M shows the sequence alignment of CLCA2 and the evolutionary conservation of 3UTR among different species. CLCA2 3UTR contained three possible MIR375 target sites in the seed region (Fig. S1E). Figure S1F depicts the successful construction of the vector. We co-transfected MIR375 with wild-type 3UTR of CLCA2 or mutated 3UTR. Following transfection with the wild-type 3UTR of CLCA2, the LacZ/Fluc reporter activity was notably repressed, while this repression was diminished after transfection with the mutated 3UTR (Fig. S1G). Additionally, the MIR375 inhibitor partially reversed the LacZ/Fluc reporter activity after transfection with 3UTR of CLCA2 (Fig. 1I). These results indicated that MIR375 can orchestrate CLCA2 expression by binding to its 3′UTR sequence.
Importantly, we evaluated the therapeutic effect of injection of rAAV9 delivery TuD-Mir375 in mice with PCa. After the rAAV9-TuD-Mir375 injection, mouse weight was monitored every two weeks, and intriguingly, no substantial difference in body weight was discerned (Fig. 1J). Notably, the median survival time of mice in the AAV9-TuD-Mir375 group (308.5 days) was significantly longer than that of mice in the phosphate-buffered saline (PBS) group (220 days) (Fig. 1K). More obvious tumorigenesis was observed in the PBS group than in the TuD-Mir375 group by hematoxylin and eosin staining (Fig. 1L). The Preliminary experiments suggest that injection of rAAV9 delivery TuD-Mir375 resulted in longer median survival times and less tumor angiogenesis in mice with PCa.
Our study unveiled that the expression of MIR375 in tumor sites is significantly higher than that in normal tissue. Mainly as a tumor promoter, MIR375 is reported to be associated with the prognosis of PCa patients.4 Specifically, apoptosis markers could be strongly inhibited by MIR375 when the expression of downstream epithelial–mesenchymal transition and andrology receptor markers could be up-regulated.2 Consequently, MIR375 emerges as a compelling candidate for novel therapeutic strategies in PCa patients. In vitro experiments were also conducted to systematically assess the impact of MIR375 on the PCa cells. Our findings have elucidated that MIR375 exerts a potent stimulatory effect on PCa progression by modulating the expression of downstream genes. Among these genes, CLCA2 was identified as a potential target of MIR375. CLCA2, a member of the p53 family, is known for its capacity to suppress the proliferation, migration, and invasion of cancer cells.5 We found out that MIR375 could orchestrate CLCA2 expression by binding to its 3′UTR sequence when the expression of CLCA2 was significantly negatively correlated with the MIR375. By this means, the inhibition of MIR375 can activate the downstream tumor suppressor, CLCA2, thereby impeding the progression of PCa cells. Particularly, rAAV9 was proven effective in the transduction of target genes.3 The rAAV9-TuD-Mir375 was constructed and intratumorally injected into the mice to further validate the effectiveness of AAV-based therapy. While no discernible difference in mouse weight was observed between the two groups, the rAAV9-TuD-Mir375 group exhibited a significantly prolonged survival time in comparison to the PBS group. Apart from that, the tumors in both groups were retrieved to perform hematoxylin and eosin staining. Similarly, the tumorigenesis was decreased after the delivery of TuD-Mir375 compared with the PBS group. Thus, the in vivo experiments have corroborated that the down-regulation of Mir375 holds promise for enhancing the survival of TRAMP mice.
In conclusion, targeting MIR375 may effectively down-regulate the proliferation and invasion of PCa cells, and rAAV9 was a robust tool for the transduction of genes into target cells or organs. These findings may pave an avenue for the genetic therapy of PCa, which constitutes the comprehensive treatments and management of PCa.
Ethics declaration
The animal study was reviewed and approved by the Animal Ethics Review Committees of the West China Hospital.
Author contributions
JA and XY conceived the project and drafted the manuscript. JL, XY, ZH, and DL performed the experiments. TZ and JY collected the public data and performed the analysis. JA revised the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of interests
The authors declare no competing interests.
Funding
This study was supported by the National Key R&D Plan (2023YFC3403200), grants from the 10.13039/501100001809National Natural Science Foundation of China (No. 82070784 and 81702536) and a grant from the Science & Technology Department of Sichuan Province, China (No. 2022JDRC0040).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mukherji D.Omlin A.Pezaro C.Shamseddine A.de Bono J.Metastatic castration-resistant prostate cancer (CRPC): Preclinical and clinical evidence for the sequential use of novel therapeutics Cancer Metastasis Rev 33220145555662445275810.1007/s 10555-013-9473-1 · doi ↗ · pubmed ↗
- 2Gan J.Liu S.Zhang Y.micro RNA-375 is a therapeutic target for castration-resistant prostate cancer through the PTPN 4/STAT 3 axis Exp Mol Med 5482022129013053604237510.1038/s 12276-022-00837-6PMC 9440249 · doi ↗ · pubmed ↗
- 3Ai J.Wang D.Wei Q.Li H.Gao G.Adeno-associated virus serotype vectors efficiently transduce normal prostate tissue and prostate cancer cells Eur Urol 69120161791812652696010.1016/j.eururo.2015.10.019 · doi ↗ · pubmed ↗
- 4Huang X.Yuan T.Liang M.Exosomal mi R-1290 and mi R-375 as prognostic markers in castration-resistant prostate cancer Eur Urol 671201533412512985410.1016/j.eururo.2014.07.035PMC 4252606 · doi ↗ · pubmed ↗
- 5Tanikawa C.Nakagawa H.Furukawa Y.Nakamura Y.Matsuda K.CLCA 2 as a p 53-inducible senescence mediator Neoplasia 14220121411492243192210.1593/neo.111700 PMC 3306259 · doi ↗ · pubmed ↗