Characteristics of the chloroplast genome and genetic divergence of Tamarix hispida Willd. 1816 (Tamaricaceae)
Haowen Tian, Xiaojun Shi, Hongxiang Zhang

TL;DR
This study examines the chloroplast genome of Tamarix hispida to understand its genetic divergence and evolutionary history in northwestern China.
Contribution
The study provides new insights into the genetic structure and evolutionary divergence of Tamarix hispida using chloroplast genome sequencing.
Findings
The T. hispida chloroplast genome is 156,164–156,186 bp long with a quadripartite structure and 131 annotated genes.
Phylogenetic analysis identified two lineages within T. hispida that diverged 3.15 million years ago.
The species exhibits low genetic diversity, highlighting the need for conservation strategies.
Abstract
Tamarix hispida Willd. 1816, a crucial native plant species in the arid desert region of northwestern China, plays a significant role in maintaining ecological stability. It is instrumental in addressing soil salinity–alkalinity and heavy metal pollution. This research aims to analyze the phylogenetic divergence pattern and evolutionary history of T. hispida by comparing chloroplast genome structures across different populations. Despite the minimal differences in chloroplast genome structure due to conserved genes and junction regions, sequencing was conducted using the Illumina NovaSeq platform to verify the historical evolutionary processes between different populations, followed by assembly and annotation. The results revealed that the T. hispida chloroplast genome is approximately 156,164–156,186 bp in length, with a quadripartite structure and 131 annotated genes. Phylogenetic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1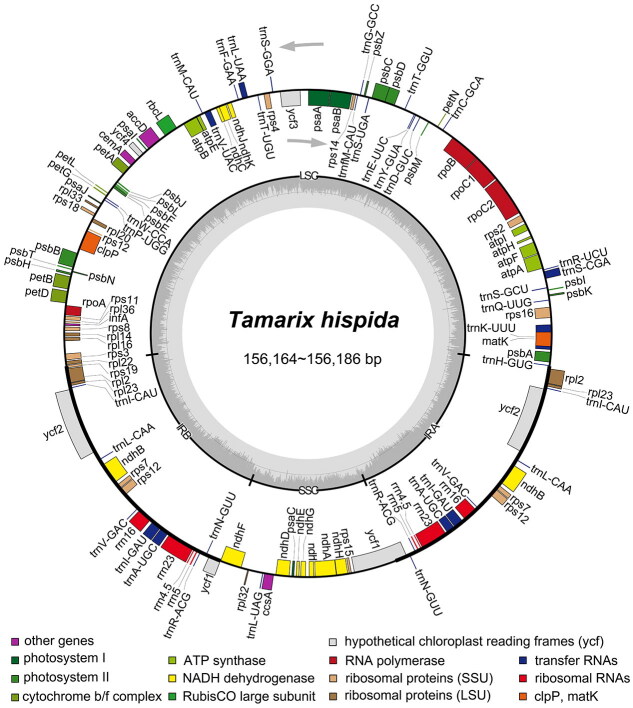 Figure 2
Figure 2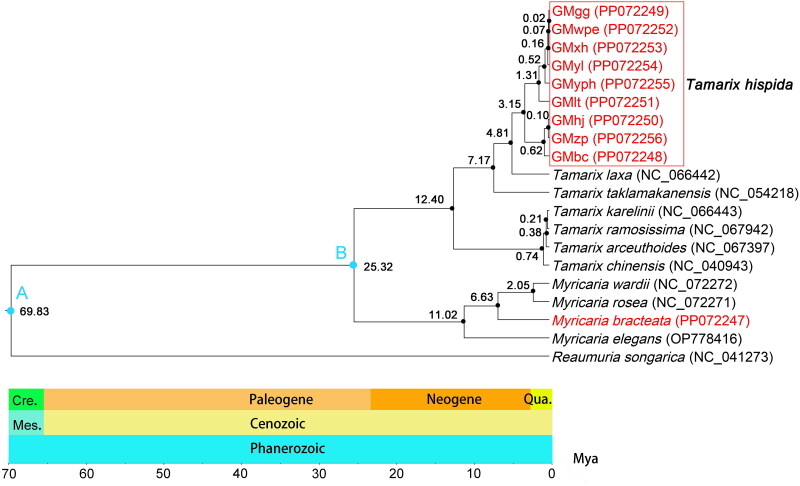 Figure 3
Figure 3| Population code | Locality | Longitude | Latitude |
|---|---|---|---|
| GMbc | Bachu, Xinjiang | 78.5028 | 39.7985 |
| GMgg | Tuokexun, Xinjiang | 88.5531 | 42.5945 |
| GMhj | Hejing, Xinjiang | 86.4750 | 42.2106 |
| GMIt | Bugur, Xinjiang | 84.7892 | 41.6112 |
| GMwpe | Shufu, Xinjiang | 75.5468 | 39.2530 |
| GMxh | Xinhe, Xinjiang | 82.5736 | 41.6587 |
| GMyl | Yuli, Xinjiang | 86.4083 | 41.1596 |
| GMyph | Yopurga, Xinjiang | 76.4980 | 39.2727 |
| GMzp | Poskam, Xinjiang | 77.2671 | 38.2362 |
- —Third Xinjiang Scientific Expedition Program
- —“Western Light” program of the Chinese Academy of Sciences
- —Second Tibetan Plateau Scientific Expedition and Research [STEP] Program
- —National Natural Science Foundation of China10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Genetic diversity and population structure · Chromosomal and Genetic Variations
Introduction
The Tamarix hispida Willd. 1816, a significant native plant species in the arid desert region of northwestern China within the Tamarix genus, plays a vital role in maintaining ecological stability (Gaskin 2003). It aids in the restoration of soil salinity–alkalinity and heavy metal pollution (Pang et al. 2022; Xie et al. 2023). Nevertheless, self-pollination tendencies and occasional secondary flowering in the Tamarix genus have caused significant hybridization, complicating the accurate identification of Tamarix species (Terrones and Juan 2023). The Tamarix genus, belonging to the family Tamaricaceae, comprises approximately 70–75 recognized species, many of which are adapted to extreme environmental conditions (Villar et al. 2019). Chloroplast genome sequences unravel extensive sequence and structural diversities within and among plant species, offering valuable insights into comprehending climate adaptation in economically crucial crops, facilitating the breeding of closely related species, and identifying and conserving valuable traits (Llorente et al. 2021). Despite the significant role of chloroplast genomes in phylogenetic studies, there is a lack of comprehensive phylogenetic research on Tamarix species, making it essential to explore their genetic diversity and evolutionary history.
To enhance the differentiation of T. hispida and investigate potential discrepancies in chloroplast genomes among distinct lineages, we conducted a comprehensive study to establish a benchmark for future chloroplast genome research on other species within the Tamaricaceae family. Given the highly preserved nature of chloroplast genes, our sampling encompassed individuals from various locations, comprising a total of nine T. hispida specimens. Each specimen was deliberately selected at a minimum distance of 100 km from one another to evaluate the possible impact of geographic isolation on chloroplast genomes. Genetic diversity in chloroplast genomes reflects the historical evolution and geographical dispersion of plants, along with the phylogenetic associations among various species (Figure 1).
Tamarix hispida: vegetative morphology. (A) Plant; (B) capsule (conic); (C) petals (petals 5, spreading, reflexed in upper part, purple-red or red); (D) inflorescence. Photo taken by Haowen Tian in Yopurga, Xinjiang.
Materials and methods
Experimental materials, sequencing, and chloroplast genome assembly
2.1.
Leaf samples were collected from nine populations of T. hispida in Xinjiang, China, covering the primary distribution area of the species (Table 1). Voucher specimens were stored at the Specimen Museum of Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences (XJBI, Hongxiang Zhang, [email protected]) (Table S1). The samples underwent DNA extraction at Shanghai Personalbio Technology Co., Ltd. (Shanghai, China) using the improved CTAB method (Porebski et al. 1997). Subsequently, we evaluated the DNA extraction quality via 0.8% agarose gel electrophoresis and quantified the DNA with a UV spectrophotometer. A library comprising 400 bp insert fragments was then constructed on the Illumina NovaSeq platform (San Diego, CA), and paired-end sequencing was performed to acquire 150 bp sequences from both ends of each read. The sequencing depth is shown in Figure S1. Raw data quality control was conducted using Fastp v0.23.1 (Chen et al. 2018), and assembly was performed with GetOrganelle v 1.7.5 (Jin et al. 2020) software. The chloroplast genome was annotated using PGA (Qu et al. 2019) software and assembly results were manually refined with Geneious v 9.0.2 (Kearse et al. 2012) software. The assembled chloroplast genome was deposited in the NCBI database (Table S1). OGDRAW was utilized to create a circular map of the chloroplast (https://chlorobox.mpimp-golm.mpg.de/OGDraw.html). CPGview (Liu et al. 2023) was used to generate cis-spliced and trans-spliced gene illustrations, as shown in Figures S2 and S3.
Gene maps of Tamarix hispida chloroplast genomes. Genes located on the inner side of the circle undergo transcription in a clockwise direction, whereas genes on the outer side are transcribed in a counterclockwise direction. Dark gray and light gray color represent guanine and cytosine (GC) content and adenine and thymine (AT) content, respectively.
The phylogenetic tree and divergence time of the genus Tamaricaceae were constructed using BEAST software based on chloroplast genome sequences. Fossil calibration points A and B were used to calibrate the tree. Geological periods are shown at the bottom. Cre., Mes., and Qua. represent Cretaceous, Mesozoic, and Quaternary periods, respectively. The analysis reveals two main lineages of T. hispida, diverging approximately 3.15 million years ago. The following sequences were used: Reaumuria songarica (accession number: NC_041273, Yao et al. 2019), Myricaria elegans (accession number: OP778416, unpublished), Myricaria bracteata (accession number: PP072247, this study), Myricaria rosea (accession number: NC_072271, unpublished), Myricaria wardii (accession number: NC_072272, unpublished), Tamarix chinensis (accession number: NC_040943, Yao et al. 2019), Tamarix arceuthoides (accession number: NC_067397, unpublished), Tamarix ramosissima (accession number: NC_067942, unpublished), Tamarix karelinii (accession number: NC_066443, unpublished), Tamarix taklamakanensis (accession number: NC_054218, unpublished), Tamarix laxa (accession number: NC_066442, unpublished), GMbc (accession number: PP072248, this study), GMzp (accession number: PP072256, this study), GMhj (accession number: PP072250, this study), GMlt (accession number: PP072251, this study), GMyph (accession number: PP072255, this study), GMyl (accession number: PP072254, this study), GMxh (accession number: PP072253, this study), GMwpe (accession number: PP072252, this study), and GMgg (accession number: PP072249, this study).
Phylogenetic analysis
2.2.
To elucidate the phylogenetic relationship among nine populations of T. hispida and other Tamarix species, chloroplast genomes of 10 Tamaricaceae species (Table S1) were also downloaded from the GenBank database, with Reaumuria songarica (NC_041273) as an outgroup. In total, nine T. hispida samples, six other Tamarix species, one Reaumuria specie, and four Myricaria species were used in the phylogenetic tree construction. The MAFFT v7.520 software was subsequently utilized for alignment, and the resulting FASTA file was converted into a NEX file format using Geneious software. The BEAST v1.6.1 software was then employed to estimate divergence time, with two fossil calibration points at 25 Ma and 70 Ma (Zhang et al. 2014). The chain length of MCMC was set to 1,000,000,000, while all other parameters were retained with default values. Finally, the ML phylogenetic tree (Wu et al. 2015) was constructed using IQ-TREE v2.2.2.6 (Nguyen et al. 2015), which selected the best model as TVM + F + I + G4.
Results
Basic characteristics and comparative analysis of the chloroplast genomes
3.1.
The complete chloroplast genome of T. hispida (accession numbers: PP072247–PP072256) spans a length of 156,164–156,186 bp, encompassing a large single-copy (LSC, 84,791–84,807 bp) region, a small single-copy (SSC, 18,250–18,257 bp) region, and two inverted repeats (IRs, 26,561 bp each) (Figure 2). The GC contents of the LSC, SSC, and IR regions are 34.16%, 29.58–29.61%, and 42.51%, respectively, culminating in an overall GC content of 36.5% for the T. hispida chloroplast genome. The genome possesses 131 genes, comprising 85 protein-coding genes, 37 tRNA genes, and eight rRNA genes. Each of thirteen protein-coding genes (rpoC1, atpF, rps16, petB, petD, two rpl2, rpl16, ndhA, and two ndhB) and seven tRNA genes (two trnA-UGC, two trnI-GAU, trnK-UUU, trnL-UAA, trnS-CGA, and trnV-UAC) contain one intron, while three protein-coding genes (two rps12, ycf3, and clpP) contain two introns.
Phylogenetic relationships
3.2.
The maximum-likelihood (ML) tree obtained from IQ-TREE correlated with the divergence time tree generated by BEAST; hence, the ML tree result was omitted from the phylogenetic analysis (Figure 3). T. hispida owns the closest relationship with T. laxa, and a more distant relationship with Reaumuria than Myricaria, in consistent with the prevailing morphological classification. The nine T. hispida populations were clustered into two lineages. The divergence between T. hispida and related species occurred at 4.81 Ma, whereas the divergence between lineage one and lineage two took place at 3.15 Ma.
Discussion and conclusions
The genus Tamarix originated in the ancient Mediterranean coastal region, with its present-day distribution influenced by factors like the retreat of the Tethys Sea, tectonic movements since the Tertiary period, and the impacts of Quaternary glaciation and interglacial cycles (Daoyuan et al. 2003; Anzidei et al. 2014). Tamarix tends to be easy hybridization. Due to varying degrees of gene flow, species with different morphological characteristics coexist in the same area (Sheidai and Koohdar 2023). Hence, hybridization may contribute to the divergence of the two lineages. Explaining the variation resulting from hybridization at the chloroplast genome level proves to be a challenging task. Therefore, further research in diverse areas is needed to provide a more comprehensive and clear understanding. Importantly, the constructed phylogenetic tree in this study indicates that the complete chloroplast genome can serve as a valuable tool for species identification.
Supplementary Material
Supplemental Material.docx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Anzidei M, Lambeck K, Antonioli F, Furlani S, Mastronuzzi G, Serpelloni E, Vannucci G. 2014. Coastal structure, sea-level changes and vertical motion of the land in the Mediterranean. Geol Soc Lond Spec Publ. 388(1):453–479. doi:10.1144/SP 388.20. · doi ↗
- 2Chen S, Zhou Y, Chen Y, Gu J. 2018. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 34(17):i 884–i 890. doi:10.1093/bioinformatics/bty 560.30423086 PMC 6129281 · doi ↗ · pubmed ↗
- 3Daoyuan Z, Borong P, Linke Y. 2003. The photogeographical studies of Tamarix (Tamaricaceae). Plant Divers. 25:1.
- 4Gaskin JF. 2003. Tamaricaceae. In: Kubitzki K, Bayer C, editors. Flowering plants·Dicotyledons: Malvales, Capparales and non-betalain Caryophyllales. The families and genera of vascular plants. Berlin, Heidelberg: Springer; p. 363–368.
- 5Jin J, Yu W, Yang J, Song Y, De Pamphilis CW, Yi T, Li D. 2020. Get Organelle: a fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 21(1):241. doi:10.1186/s 13059-020-02154-5.32912315 PMC 7488116 · doi ↗ · pubmed ↗
- 6Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, et al. 2012. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 28(12):1647–1649. doi:10.1093/bioinformatics/bts 199.22543367 PMC 3371832 · doi ↗ · pubmed ↗
- 7Liu S, Ni Y, Li J, Zhang X, Yang H, Chen H, Liu C. 2023. CPG View: a package for visualizing detailed chloroplast genome structures. Mol Ecol Resour. 23(3):694–704. doi:10.1111/1755-0998.13729.36587992 · doi ↗ · pubmed ↗
- 8Llorente B, Segretin ME, Giannini E, Lobais C, Juárez ME, Paulsen IT, Blanco NE. 2021. Homecoming: rewinding the reductive evolution of the chloroplast genome for increasing crop yields. Nat Commun. 12(1):6734. doi:10.1038/s 41467-021-26975-5.34795241 PMC 8602674 · doi ↗ · pubmed ↗