Loss of transcriptional regulator of phospholipid biosynthesis alters post-translational modification of Sec61 translocon beta subunit Sbh1 in Saccharomyces cerevisiae
Jacob M. Miller, Mary E. Tragesser-Tiña, Samantha M. Turk, Eric M. Rubenstein

TL;DR
Deleting a gene that regulates phospholipid production in yeast changes a protein's modification, possibly affecting cell function.
Contribution
Shows that lipid biosynthesis disruption alters protein modification, not degradation, in yeast ER.
Findings
INO4 deletion does not destabilize Sbh1 or Cue1 proteins.
Sbh1 shows altered electrophoretic mobility in ino4Δ yeast.
Phospholipid changes may affect post-translational modifications.
Abstract
We recently discovered that disrupting phospholipid biosynthesis by eliminating the Ino2/4 transcriptional regulator impairs endoplasmic reticulum (ER)-associated degradation (ERAD) in Saccharomyces cerevisiae , but the mechanism is unclear. Phosphatidylcholine deficiency has been reported to accelerate degradation of Sec61 translocon beta subunit Sbh1 and ERAD cofactor Cue1. Here, we found that, unlike targeted phosphatidylcholine depletion, INO4 deletion does not destabilize Sbh1 or Cue1. However, we observed altered electrophoretic mobility of Sbh1 in ino4 Δ yeast, consistent with phospholipid-responsive post-translational modification. A better understanding of the molecular consequences of disrupted lipid homeostasis could lead to enhanced treatments for conditions associated with perturbed lipid biosynthesis.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
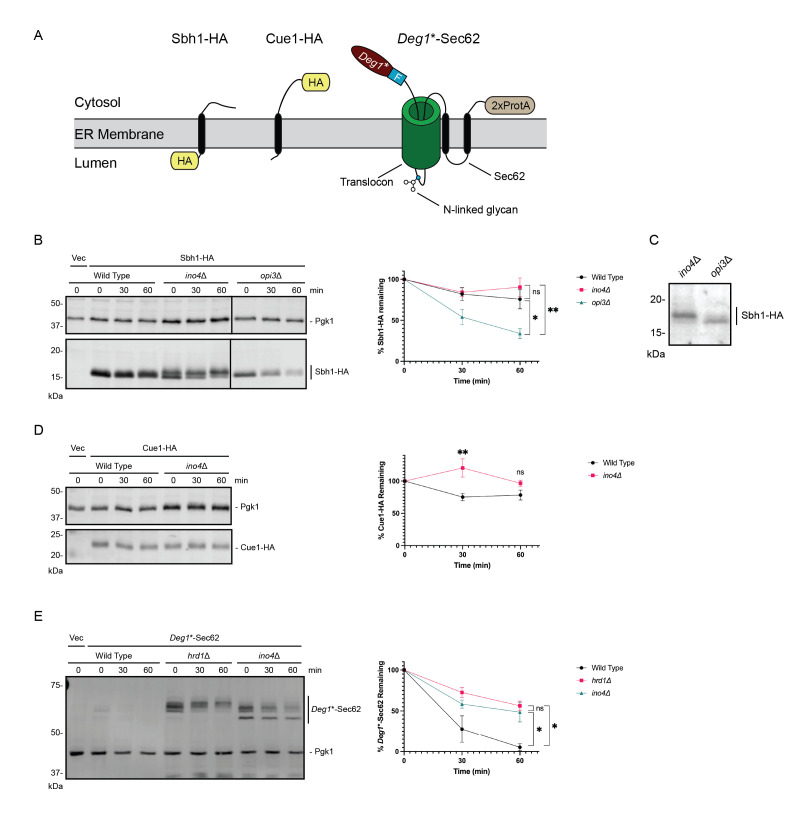 Figure 1
Figure 1|
|
|
|
VJY474 |
|
|
VJY476 (alias BY4741) |
|
|
VJY511 |
|
|
VJY1071 |
|
|
|
|
|
|
|
pVJ27 |
pRS316 |
Empty vector (CEN,
|
|
|
pVJ40 |
pRS315 |
Empty vector (CEN,
|
|
|
pVJ317 |
|
| |
|
pVJ667 |
pGT181 |
Cue1-HA driven by
|
|
|
pVJ668 |
pGT183 |
Sbh1-HA driven by
|
|
- —National Institutes of Health (United States)https://ror.org/01cwqze88
- —Ball State University (United States)https://ror.org/00k6tx165
- —Ball State University (United States)https://ror.org/00k6tx165
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA Research and Splicing · Endoplasmic Reticulum Stress and Disease · RNA and protein synthesis mechanisms
Description
Many diseases are characterized by defects in phospholipid homeostasis (Arendt et al., 2013; Bargui et al., 2021; Kim et al., 2016; Kosicek & Hecimovic, 2013; Li et al., 2006; Mitsuhashi & Nishino, 2011) , and a growing body of literature functionally links phospholipid metabolism and protein degradation (Halbleib et al., 2017; Ho et al., 2020; Hwang et al., 2023; Jonikas et al., 2009; Promlek et al., 2011; Shyu et al., 2019; Thibault et al., 2012; To et al., 2017; Volmer et al., 2013; Xu & Taubert, 2021) . It is therefore important to understand how cellular lipid composition impacts protein dynamics. We recently identified INO4 in a screen for genes required for endoplasmic reticulum (ER)-associated degradation (ERAD) in *Saccharomyces cerevisiae * (Turk et al., 2023) .
- INO4* encodes one subunit of the heterodimeric Ino2/4 master transcriptional regulator of phospholipid biosynthetic genes (Ambroziak & Henry, 1994; Henry et al., 2012) . *INO4 * deletion stabilizes a broad panel of ERAD substrates of both major yeast ERAD ubiquitin ligases (Hrd1 and Doa10) without globally impairing the ubiquitin-proteasome system (Turk et al., 2023) . The precise molecular mechanism(s) underlying ERAD disruption by INO4 deletion remain(s) to be elucidated.
Loss of INO4 broadly disrupts lipid homeostasis, as Ino2/4 promotes expression of genes required for synthesis of phosphatidylcholine, phosphatidylinositol, and phosphatidylethanolamine, among other phospholipids. Rescue of the ino4 Δ ERAD defect by supplementation with phospholipid metabolites and intermediates whose uptake or synthesis are controlled by Ino2/4-regulated genes confirmed the degradation impairment is linked to altered phospholipid metabolism (Turk et al., 2023) . Genetic disruption of several lipid biosynthetic pathways (including those specifically responsible for phosphatidylcholine, phosphatidylinositol, and sterol synthesis) impairs ERAD, indicating ER protein quality control is highly sensitive to alterations in membrane composition (Turk et al., 2023) .
We found loss of the phospholipid methyltransferase Opi3 (required for phosphatidylcholine synthesis) significantly stabilizes Deg1 *-Sec62, a model translocon-clogging ERAD substrate of the Hrd1 ubiquitin ligase (Turk et al., 2023) . Conversely, others have observed OPI3 deletion accelerates Doa10-dependent ERAD of a panel of transmembrane ER proteins, including Sec61 translocon beta subunit Sbh1 and ERAD cofactor Cue1 ( Figure 1A ) (Shyu et al., 2019) . We reasoned that depletion of either Sbh1 or Cue1 might explain the profound ERAD defect in yeast lacking Ino4.
We therefore performed cycloheximide chases and western blots to analyze degradation of Sbh1-HA and Cue1-HA in ino4 Δ yeast. As previously reported, we observed destabilization of the tail-anchored, single-pass, transmembrane Sbh1-HA in phosphatidylcholine-deficient opi3 Δ yeast ( Figure 1B ). By contrast, loss of INO4 did not alter Sbh1-HA degradation kinetics. However, a modified, higher molecular-weight species of Sbh1-HA reproducibly accumulated in ino4 Δ yeast, and to a more modest extent in opi3 Δ yeast, relative to wild type yeast ( Figure 1B, 1C). We speculate slowed migration reflects phosphorylation, as multiple proteomic analyses have identified three phosphorylated residues of Sbh1 (Lanz et al., 2021; MacGilvray et al., 2020; Swaney et al., 2013; Zhou et al., 2021) . Additional analyses will be required to definitively characterize the nature of Sbh1 modification. We did not observe destabilization of the single-pass transmembrane Cue1-HA in ino4 Δ yeast ( Figure 1D ). On the contrary, Cue1-HA was marginally stabilized in ino4 Δ yeast, consistent with broad ERAD impairment by this mutation. Finally, we reproduced our previous observation that loss of INO4 stabilizes and impedes post-translational modification of the model translocon-clogging Hrd1 substrate, Deg1 *-Sec62 ( Figure 1E ).
As INO4 deletion does not destabilize either HA-tagged Sbh1 or Cue1, our results suggest the ERAD defect in Ino2/4-deficient yeast is not due to depletion of either protein. A precise mechanism by which altered phospholipid biosynthesis in ino2 Δ or ino4 Δ yeast impacts ERAD remains elusive. We previously observed that INO4 deletion does not globally inhibit ER translocation (Turk et al., 2023) . However, phospholipid-sensitive alteration in post-translational modification of a translocon subunit might subtly or selectively perturb translocation in a manner that compromises ERAD. Indeed, recent work indicates phosphorylation alters the conformation of the Sbh1 N-terminus and promotes translocation of a small subset of ER-targeted proteins, modestly impacting the ER proteome (Barbieri et al., 2023; Yadhanapudi et al., 2024) . Loss of Ino2/4 broadly impacts expression of genes regulating lipid homeostasis. It is therefore likely that deletion of INO2 or INO4 impacts ER physiology in myriad ways, including altered membrane fluidity, disrupted protein-protein interactions, changes in post-translational modifications, and perturbations in structure, localization, or function of ER-resident proteins.
Methods
**Cycloheximide chase and yeast cell lysis. ** Yeast were cultured in Synthetic Defined (SD) growth medium (2% dextrose, 0.67% yeast nitrogen base, 0.002% adenine, 0.004% uracil, 0.002% arginine, 0.001% histidine, 0.006% isoleucine, 0.006% leucine, 0.004% lysine, 0.001% methionine, 0.006% phenylalanine, 0.005% threonine, and 0.004% tryptophan), lacking uracil or leucine (for plasmid selection). Cycloheximide chase experiments were conducted as described (Buchanan et al., 2016) . In brief, 2.5 OD 600 units (1 OD 600 unit is equivalent to 1 mL of yeast at an OD 600 of 1.0) per time point were harvested by centrifugation and resuspended in fresh medium. Cycloheximide was added to a final concentration of 250 µg/mL. 2.4-OD 600 aliquots were collected 0, 30, and 60 minutes following cycloheximide addition and transferred to stop solution (10 mM sodium azide, 0.25 mg/mL bovine serum albumin). Proteins were extracted using the alkaline lysis method (Kushnirov, 2000; Watts et al., 2015) .
**Western blotting. ** Western blotting was conducted as described (Watts et al., 2015) . Proteins were separated by SDS-PGE and transferred to polyvinylidene difluoride (PVDF) membranes at 20 V for 1 hr at 4°C. Membranes were blocked in Tris-buffered saline (TBS; 50 mM Tris, 150 mM NaCl) with 5% skim milk powder for 1 hr at room temperature or overnight at 4°C. Antibody incubations were conducted for 1 hr at room temperature in TBS supplemented with 1% Tween 20 (TBS/T) and 1% skim milk powder. Each antibody incubation was followed by three five-minute, room-temperature washes with TBS/T. To detect HA-tagged proteins (Sbh1-HA and Cue1-HA), mouse anti-HA.11 primary antibody (Clone 16B12; BioLegend) was used at a dilution of 1:1,000. To detect Pgk1, mouse anti-Pgk1 primary antibody (Clone 22C5D8; LifeTechnologies) was used at a dilution of 1:20,000. Primary antibody incubations were followed by incubation with AlexaFluor-680-conjugated rabbit anti-mouse secondary antibody (LifeTechnologies) at a dilution of 1:20,000. The C-terminal tandem *Staphylococcus aureus * Protein A tags on Deg1 *-Sec62 were directly detected by AlexaFluor-680-conjugated rabbit anti-mouse secondary antibody (LifeTechnologies) at a dilution of 1:20,000. Membranes were imaged using the Li-Cor Odyssey DLx Imaging System and quantified using ImageStudio software (Li-Cor). To determine percent protein remaining, the ratio of signal intensity for each protein of interest (Sbh1-HA, Cue1-HA, or Deg1 *-Sec62) to the signal intensity of Pgk1 at each time point was compared to this ratio at 0 minutes for each culture.
**Statistical analysis. ** Statistical analyses were performed using GraphPad Prism (version 10). For experiments depicted in Figures 1B and 1E, means of percent protein remaining at 60 minutes were evaluated by one-way ANOVA followed by Holm-Šídák multiple comparison tests. For the experiment depicted in Figure 1D, means of percent remaining at 30 or 60 minutes were evaluated by unpaired, two-tailed t- tests.
Reagents
Yeast strains used in this study.
**: **
All strains are congenic with BY4741 (Tong et al., 2001) .
Plasmids used in this study.
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ambroziak J Henry SA 1994527 INO 2 and INO 4 gene products, positive regulators of phospholipid biosynthesis in Saccharomyces cerevisiae, form a complex that binds to the INO 1 promoter.J Biol Chem 269210021-925815344153498195172 · pubmed ↗
- 2Arendt BM Ma DW Simons B Noureldin SA Therapondos G Guindi M Sherman M Allard JP 20121015 Nonalcoholic fatty liver disease is associated with lower hepatic and erythrocyte ratios of phosphatidylcholine to phosphatidylethanolamine.Appl Physiol Nutr Metab 3831715-531233434010.1139/apnm-2012-026123537027 · doi ↗ · pubmed ↗
- 3Barbieri G Simon J Lupusella CR Pereira F Elia F Meyer H Schuldiner M Hanes SD Nguyen D Helms V Römisch K 2023111 Sec 61 channel subunit Sbh 1/Sec 61β promotes ER translocation of proteins with suboptimal targeting sequences and is fine-tuned by phosphorylation.J Biol Chem 29930021-925810289510289510.1016/j.jbc.2023.10289536639027 PMC 9947333 · doi ↗ · pubmed ↗
- 4Bargui R Solgadi A Prost B Chester M Ferreiro A Piquereau J Moulin M 2021730 Phospholipids: Identification and Implication in Muscle Pathophysiology.Int J Mol Sci 221510.3390/ijms 2215817634360941 PMC 8347011 · doi ↗ · pubmed ↗
- 5Buchanan BW Lloyd ME Engle SM Rubenstein EM 2016418 Cycloheximide Chase Analysis of Protein Degradation in Saccharomyces cerevisiae.J Vis Exp 11010.3791/5397527167179 PMC 4941941 · doi ↗ · pubmed ↗
- 6Halbleib K Pesek K Covino R Hofbauer HF Wunnicke D Hänelt I Hummer G Ernst R 201776 Activation of the Unfolded Protein Response by Lipid Bilayer Stress.Mol Cell 6741097-2765673684.e 810.1016/j.molcel.2017.06.01228689662 · doi ↗ · pubmed ↗
- 7Henry SA Kohlwein SD Carman GM 201221 Metabolism and regulation of glycerolipids in the yeast Saccharomyces cerevisiae.Genetics 19020016-673131734910.1534/genetics.111.13028622345606 PMC 3276621 · doi ↗ · pubmed ↗
- 8Ho N Yap WS Xu J Wu H Koh JH Goh WWB George B Chong SC Taubert S Thibault G 202076 Stress sensor Ire 1 deploys a divergent transcriptional program in response to lipid bilayer stress.J Cell Biol 21970021-952510.1083/jcb.20190916532349127 PMC 7337508 · doi ↗ · pubmed ↗