Infection of Phytophthora palmivora Isolates on Arabidopsis thaliana
Mariandrea García-Gaona, Hernán Mauricio Romero

TL;DR
This study shows how different isolates of Phytophthora palmivora infect Arabidopsis thaliana, revealing differences in infection processes and host responses.
Contribution
The study establishes a new model pathosystem using Arabidopsis to study P. palmivora isolates from oil palm.
Findings
Two P. palmivora isolates caused lesions and sporangia in Arabidopsis, while one isolate failed to penetrate tissue.
Infection-related genes were overexpressed in compatible isolates but not in the incompatible one.
Arabidopsis defense-related genes were regulated during interactions with the isolates.
Abstract
Phytophthora palmivora, a hemibiotrophic oomycete, causes diseases in several economically important tropical crops, such as oil palm, which it is responsible for a devastating disease called bud rot (BR). Despite recent progress in understanding host resistance and virulence mechanisms, many aspects remain unknown in P. palmivora isolates from oil palm. Model pathosystems are useful for understanding the molecular interactions between pathogens and hosts. In this study, we utilized detached leaves and whole seedlings of Arabidopsis thaliana Col-0 to describe and evaluate the infection process of three P. palmivora isolates (CPPhZC-05, CPPhZC-04, CPPhZOC-01) that cause BR in oil palm. Two compatible isolates (CPPhZC-05 and CPPhZOC-01) induced aqueous lesions at 72 h post-inoculation (hpi), with microscopic visualization revealing zoospore encysting and appressorium penetration at 3 hpi,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
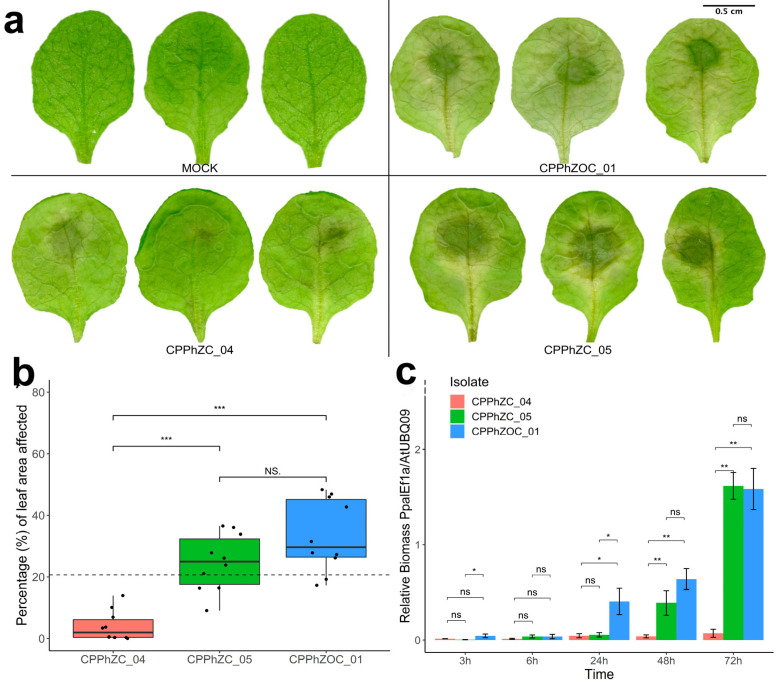 Figure 1
Figure 1 Figure 2
Figure 2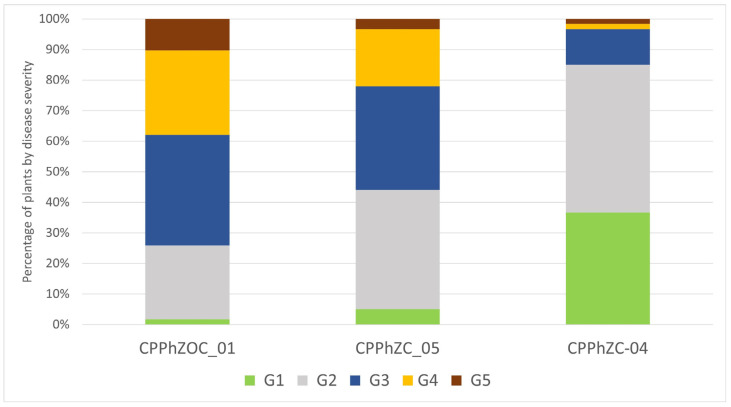 Figure 3
Figure 3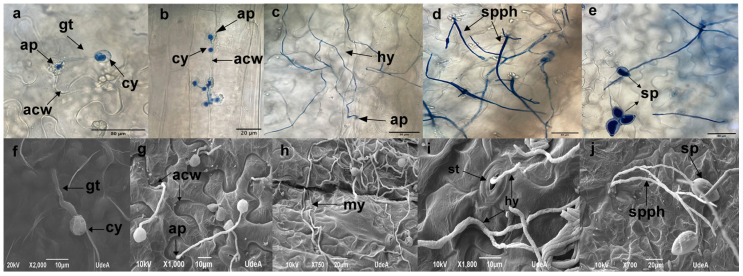 Figure 4
Figure 4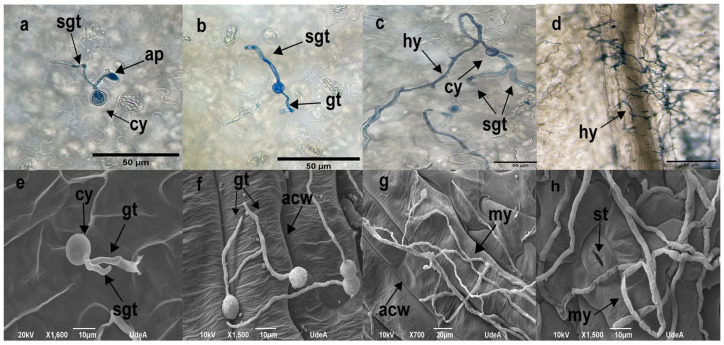 Figure 5
Figure 5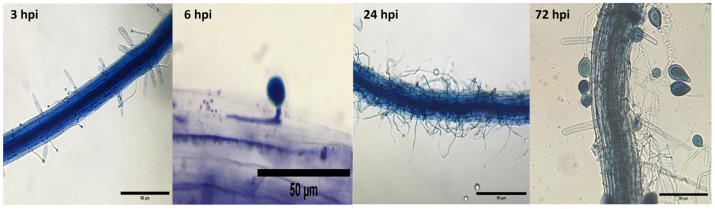 Figure 6
Figure 6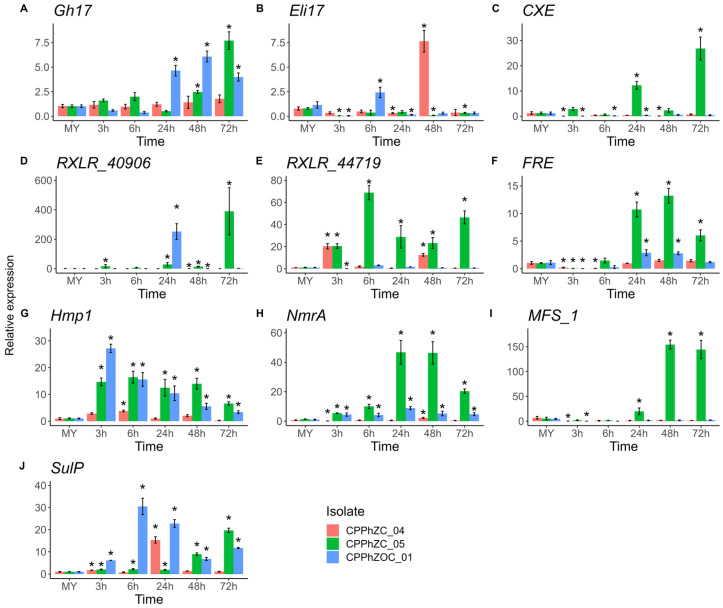 Figure 7
Figure 7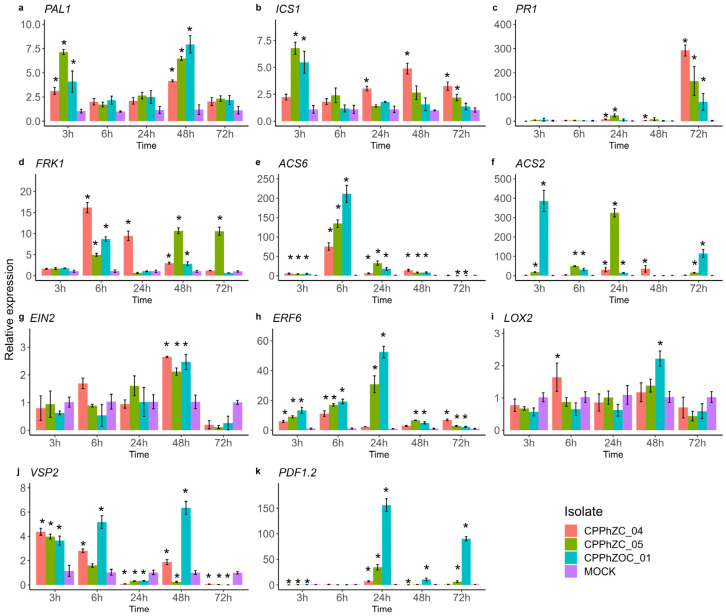 Figure 8
Figure 8- —Colombian Oil Palm Promotion Fund (FFP)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Resistance · Plant-Microbe Interactions and Immunity · Plant Pathogens and Fungal Diseases
1. Introduction
Phytophthora palmivora is a generalist pathogen that causes root, bud, and fruit rot diseases in various economically important tropical crops, including durian, papaya, coconut, rubber, and cocoa [1,2]. It is especially devastating in oil palm [3], causing bud rot disease [2,4], the main limiting factor for crop expansion in Latin America. P. palmivora is an oomycete characterized by having a hemibiotrophic life cycle. Like many oomycetes, it has an asexual life cycle, where sporangia capable of dispersing through wind or water are generated. These structures can germinate directly or by releasing zoospores [5].
In addition to the different agricultural and biological alternatives to mitigate P. palmivora infections in oil palm [6,7], studying the interaction between plants and pathogens provides a comprehensive overview of the defense mechanisms of the plant and pathogen infection. This knowledge can help to develop durable, effective, and environmentally friendly strategies [8].
Establishing model pathosystems in controlled experimental setups that mimic natural interactions is central to advancing our understanding of these interactions. Among the available model organisms, Arabidopsis thaliana stands out due to its genetic tractability, well-documented genome, and ease of transformation. Over time, Arabidopsis has become a favored choice for elucidating the complexities of host–pathogen interplay [9,10].
Within the genus Phytophthora, the infection processes of several species have been elucidated in A. thaliana. These species include Phytophthora brassicae [11], P. parasítica [12], P. capsici [13], P. cinnamomi [14], and P. palmivora [9,15]. Despite this progress, specific insights into the infection processes of different P. palmivora isolates remain unexplored, particularly in the context of economically relevant hosts like oil palm.
The aims of the study were (i) to elucidate the infection processes of three P. palmivora isolates in their interactions with Arabidopsis thaliana, (ii) to distinguish between compatible and incompatible interactions by analyzing the microscopic stages of infection, and (iii) to characterize the gene expression profiles of P. palmivora’s candidate effectors and the defense response markers in A. thaliana.
2. Materials and Methods
2.1. Plant Material
Arabidopsis thaliana ecotype Col-0 (CS667) was obtained from the ABRC collection. Maintenance of seeds and plant culture conditions followed a standard protocol [16]. Plants grew in pots with a mix of Floragard^®^ (Oldenburg, Germany) peat moss and perlite in a growth chamber at 22 °C, under a photoperiod of 10 h of light and 14 h of darkness, with a relative humidity of 60–70%.
2.2. Pathogen Growth and Infection Assay
Three Phytophthora palmivora isolates (CPPhZC-05, CPPhZC-04, CPPhZOC-01) from the Cenipalma collection which had been previously characterized [17] were grown on clarified V8 juice medium consisting of 20% V8^®^ Campbell (Camden, NJ, USA) juice, 5 g L^−1^ CaCO_3_ (Merck, Darmstadt, Germany), 50 mg L^−1^ B-sitosterol (Merck), 1.5% Bacto™ agar, and rifampicin 1 μg mL^−1^ (Merck), as previously reported [18]. The cultures were maintained under a 12 h photoperiod at 25 °C until 8 days of growth had been completed.
The detached leaf inculation was performed by following Wang et al. [12]. Twenty fully expanded leaves (leaf No 3, 4, or 5 of the rosette) from 4-week-old (growth stage 1.10–1.12) Col-0 plants were excised and placed onto moist filter paper in a tray (22 × 14 × 7 cm), then introduced into a Ziploc^®^ bag (San Diego, CA, USA). Zoospore suspensions of P. palmivora isolates (9 × 10^5^ zoospores/mL) were prepared, and a droplet of 10 µL was applied to the abaxial side of the leaf; sterile water was used as a control (mock inoculation). The release of zoospores was induced by thermal shock using sterile distilled water at 4 °C, and pathogen quantification was performed using a Neubauer Chamber. The inoculated trays were maintained at 25 °C with a 12 h light/dark photoperiod.
Twenty detached leaves per treatment (CPPhZC-05, CPPhZC-04, CPPhZOC-01 and Mock) were scanned at 72 hpi. The total area of each leaf and the infected area were calculated using Assess 2.0 [19]. Ten independent replicates were performed.
For whole plant infection, we used the methodology described by Wang et al. [12] with modifications. Ten-day-old Arabidopsis seedlings (growth stage: 1.04) were floated in a suspension of 9 × 10^5^ zoospores mL^−1^ for five seconds and then transferred onto Petri plates containing half-strength MS medium without sugar. A total of 60 plants per each treatment, distributed in 10 petri plates, were used for inoculation. Then, the plates were incubated at 23 °C with a 12 h light/dark photoperiod for 96 h.
2.3. Microscopic Visualization of the Infection Process
Leaves or whole seedlings of A. thaliana were inoculated as decribed above. Samples for each treatment were collected at 3, 6, 24, 48, and 72 hpi. They were stained using diluted trypan blue solution of 10 g phenol (Merck), 10 mL glycerol (Promega, Madison, WI, USA), 10 mL lactic acid (Merck), 10 mL destiled water, and 10 mg trypan blue (Merck) [20]. Subsequently, the samples were destained in 96% ethanol for 48 h, replacing the ethanol every 12 h until the leaves cleared. Observations were made using an Olympus (Tokyo, Japan) CX3 microscope. Three independent replicates were conducted.
For SEM visualization, three leaves from each treatment were fixed in 2.5% glutaraldehyde for 24 h and sequentially dehydrated in ethanol concentrations of 30%, 50%, 70%, 80%, 90%, and 95% for 10 min each, then finally at 100% overnight. The samples were then dried, coated with gold [4], and prepared for imaging.
2.4. Extraction of Total RNA, cDNA Synthesis, and qRT-PCR Analysis
After inoculation, ten leaves were collected at each time point, at 3, 6, 24, 48, and 72 hpi, then were homogenized with liquid nitrogen and stored at −80 °C. The extraction of RNA from the homogenized leaves for each treatment was carried out with the InviTrap^®^ Spin Plant RNA Mini Kit (Invitek, Tondela, Portugal) following the manufacturer’s protocol. The cDNA synthesis was performed using 1 ng of RNA with the SuperScript IV Reverse Transcriptase (RT) kit (Thermo Fisher Scientific, Waltham, MA, USA). All cDNA samples were diluted 3X with nuclease-free water and stored at −20 °C until further use. The gene expression quantification by RT-qPCR was performed with 4 μL of dilute cDNA and EvaGreen^®^ (Biotium, Fremont, CA, USA) dye in a final volume of 10 μL, according to the manufacturer’s instructions. The qRT-PCR reactions were performed using a LightCycler 480 from Roche^®^ (Mannheim, Germany). The fold change (FC) values of the qRT-PCR reactions were calculated using the Livak method [21], and statistical analyses were performed using R Studio version 4.1.0.
Ten genes of Phytophthora palmivora and ten of Arabidopsis thaliana were analyzed. Genes of P. palmivora were selected based on previous analyses that showed key genes for the pathogenesis process [22], and the Arabidopsis genes were selected as hallmarks for plant defense response [23]. Primers for qRT-PCR analyses were designed using Primer3 (v.0.4.0) and are listed in Table 1.
The relative biomass of P. palmivora was calculated using the PpalEF1a/AtUBC9 genes ratio. The relative expression for each gene was calculated using PpalEF1a and AtUBC9 genes as normalizers for P. palmivora and Arabidopsis, respectively. Controls for the calculation of ΔΔCt consisted of cDNA from axenically cultivated P. palmivora containing mycelia and sporangia (MY) and sterile water-inoculated leaves of Arabidopsis (Mock). Two technical replicates were analyzed for each of the three independent sample replicates at any given time point/treatment.
3. Results
3.1. Infection Dynamics of P. palmivora on Arabidopsis Col-0
Three P. palmivora isolates were assessed for their infection capabilities on Arabidopsis Col-0. The isolates CPPhZC-05 and CPPhZOC-01 produced lesions covering 30% to 60% of the leaf area 72 h post-inoculation (Figure 1a). In contrast, CPPhZC-04 produced significantly smaller lesions, covering only 0% to 15% of the leaf area (Figure 1b). The biomass quantification of P. palmivora confirmed increased colonization over time for CPPhZC-05 and CPPhZOC-01, whereas CPPhZC-04 exhibited minimal growth (Figure 1c). These results show that visible lesions are correlated with pathogen development.
3.2. Root Inoculation and Symptom Evaluation
P. palmivora is a soil-borne pathogen; thus, the roots of 10-day-old Col-0 seedlings were immersed in a zoospore suspension. Symptoms across the whole plant were observed three to four days post-inoculation, ranging from no visible symptoms to complete wilting. As a result, we categorized the disease severity on a scale: G1 (healthy plants), G2 (wilting and yellowing of the oldest leaves), G3 (brown coloration on two leaves), G4 (more than two brown-colored leaves), and G5 (dead plants) (Figure 2).
The Arabidopsis plants exhibited different levels of disease severity after infection with P. palmivora. Around 60 to 70% of the plants inoculated with CPPhZC-05 and CPPhZOC-01 showed symptoms of wilting or complete collapse (G3, G4, G5) within four days (Figure 3), whereas 40% of the plants treated with CPPhZC-04 exhibited no symptoms or only mild symptoms (G1).
3.3. Microscopic Analysis of Infection Mechanisms
Detailed microscopic observations of detached leaves using optical and SEM microscopy revealed that CPPhZC-05 and CPPhZOC-01 zoospores germinated and formed appressoria, penetrating the tissue at anticlinal cell wall junctions within 3 to 6 h post-inoculation (Figure 4a,b,f,g). During this biotrophic phase, frequent callose formations were observed at penetration sites (Figure 4b), although no macroscopic reactions were evident. By 24 and 48 h post-inoculation, the tissue showed extensive colonization by a dense network of hyphae (Figure 4c,h), and by 72 h, sporangiophores with ovoid sporangia had emerged from the tissue or stomata (Figure 4d,e,i,j), indicating that CPPhZC-05 and CPPhZOC-01 completed their life cycles in Arabidopsis.
In contrast, CPPhZC-04 demonstrated a reduced ability to penetrate tissue. Initial germ tube formation was observed between 3 and 6 h post-inoculation (Figure 5a,b,f), with continued growth on the leaf surface by 24 and 48 h to form hyphae (Figure 5c,g). By 72 h, although mycelium had developed, no sporangiophores or hyphae emerged from the tissue or stomata (Figure 5d,h), a behavior similar to other Phytophthora species which are unable to infect Arabidopsis or tobacco, as described in previous studies [11,13,24,25].
Furthermore, CPPhZOC_01 and CPPhZC_05 were able to grow internally, producing haustoria and sporangia (Figure 6), whereas CPPhZC_04 exhibited only vegetative growth with sporangia formation, failing to induce symptoms comparable to the other two isolates.
3.4. The Expression of Infection-Related Genes of P. palmivora Is Upregulated during Colonization of Arabidopsis Leaves
Effectors are defined as molecules that manipulate cell structure and function. They are often assigned a dual function, facilitating infection or inducing the plant’s defense response [26]. Usually, the transcriptional induction of pathogen genes encoding secreted effector proteins is a hallmark of plant–pathogen interaction [27]. We examined the expression of five candidate effectors, two with hydrolytic domains (Gh17 (Glycosyl hydrolase family 17) and CXE (Coesterase type B)), an elicitin (hereafter called Eli17), and two RXLR motif effectors (RXLR_40906 and RXLR_44719). These are involved in oil palm colonization of the isolate CPPhZC_05. Additionally, we evaluated five other genes that included the transcription factor NmrA (nitrogen metabolic regulation A), transmembrane proteins such as Hmp1 (Haustorium-specific membrane protein), SulP (Sulfate Permease), MFS_1 (major facilitator, sugar transporter), and FRE (ferric reductase), which are co-expressed with candidate effectors during oil palm infection [22]. Our results show statistical differences in the expression of each gene compared to the controls (Figure 7).
At the early stages, i.e., 3 and 6 h post-infection (hpi), the genes Hmp1, NmrA, RXLR_44719, and SulP were significantly upregulated compared to axenically cultured conditions. However, these expressions were variable among isolates. For instance, the isolate CPPhZC_04 only co-expressed RXLR_44719 and SulP during this period, without any other genes. On the other hand, the isolate CPPhZOC_01 did not express the effector RXLR_44719; instead, it expressed Eli17.
By 24 hpi, there were more notable changes in expression. In addition to the previously mentioned genes, there was also expression of genes such as FRE, MFS_1, CXE, Gh17, and RXLR_40906. At 48 h and 72 h, compatible isolates continued expressing FRE and MFS_1 genes, co-expressing with Hmp1, NmrA, and Gh17.
The results point out that, during P. palmivora infection of Arabidopsis, the isolates CPPhZC_05 and CPPhZOC_01 secrete hydrolytic effectors and RXLR motif effectors, which are co-expressed with genes responsible for haustoria generation, nitrogen metabolism, and acquisition of nutrients such as sugars and sulfates. However, specific gene expression differs between the two compatible isolates, indicating that successful infection may occur through a distinct set of genes over time.
Regarding the incompatible isolate CPPhZOC_04, it is important to note that the expression of Hmp1, associated with the germination of cysts, appressorium, and haustoria generation, remained low during the infection. This confirms the microscopic visualization results, where appressorium and tissue penetration were rarely seen. Only two effectors, RXLR_44719 and ELI17, were upregulated at 3 and 48 hpi, respectively.
3.5. Arabidopsis Induces Defense-Related Genes in Response to P. palmivora Infection
To determine whether Arabidopsis induces its defense system in response to P. palmivora, we examined genes of the salicylic acid (SA), jasmonic acid (JA), and ethylene (ET) pathways. These hormones are the main key players in plants’ innate immunity. We measured marker genes associated with each pathway through qRT-PCR.
ICS1 (Isochorismate synthase1) and PAL1 (Phenylalanine ammonia-lyase 1) are involved in the synthesis of SA [28], while PR1 (Pathogenesis-related gene 1) is a well-established marker gene responsive to the accumulation of SA [29]. Arabidopsis upregulated both the ISC1 and PAL1 pathways in the presence of P. palmivora, indicating that AS synthesis occurred during infection (Figure 8a,b). All three isolates of P. palmivora induce AS biosynthesis; however, the compatible ones, CPPhZC_05 and CPPhZOC_01, induce a stronger response in ISC1 at the beginning of infection, followed by a decrease. However, in the presence of the incompatible isolate CPPhZC_04, ICS1 increases by 2.5-fold over time. PAL1 is also upregulated; however, it has been demonstrated that 98% of pathogen-induced SA in Arabidopsis is derived from the ICS1 pathway [28,30,31]. Hence, we assume that the major contributor to the synthesis of AS is ISC1 in the P. palmivora-Arabidopsis pathosystem. Typically, the accumulation of AS leads to the expression of PR1 [32], which correlates with the strong upregulation of PR1 generated by CPPhZC_04 at 72 hpi (Figure 8c). Also, PR1 proteins are secretory or vacuolar-target proteins with antimicrobial features [33,34].
The FRK1 gene (Flg22-induced receptor-like kinase 1) is often used as a marker for PTI (pattern-triggered immunity). It activates downstream mitogen-activated protein kinase (MAPK) pathways after plant perception of PAMPs (pathogen-associated molecular patterns) [35]. Arabidopsis usually upregulates FRK1 between 30 min and 8 h post-inoculation with a pathogen or peptide, fgl22 [36]. According to our findings, all three isolates caused upregulation of FRK1 at 3 hpi (Figure 8d). Then, at 6 hpi, FRK1 significantly increased in the presence of the isolate CPPhZC_04, but not in the presence of compatible isolates.
Another hormone involved in the plant defense response is ethylene (ET) [37]. To assess the production of ethylene (ET), we measured the expression levels of two genes: ACS2 and ACS6 (Figure 8e,f). These genes encode two different isoforms of 1-aminocyclopropane-1-carboxylic acid synthase, a crucial enzyme in the metabolic pathway for synthesizing ethylene. Our results showed that upregulation occurred between 3 and 24 hpi in the three isolates. However, the fold change was higher in the isolates CPPhZC_05 and CPPhZOC_01, indicating that ethylene is more abundantly produced when inoculating with these isolates.
Our study showed that the gene EIN2 was upregulated (Figure 8g). This gene plays a crucial role in the signaling pathway that activates the nucleus transcription complex of ET-responsive genes [38]. The gene ERF6 encodes a transcriptional factor expressed when ethylene levels rise [39]. Our results showed that this gene was expressed between 3 and 24 hpi in the three isolates (Figure 8h). Once again, the CPPhZC_05 and CPPhZOC_01 isolates showed fold changes 5 to 20 times higher.
ERF6 is known to have a role in the positive regulation of jasmonic acid (JA) [39]. Therefore, its positive expression suggests that JA is produced due to infection of the three isolates of P. palmivora. To verify this, we quantified other markers involved in the synthesis of JA. LOX2 (Lipoxygenase2) encodes a protein that is the main contributor to the synthesis of JA during leaf damage caused by wounds [40]. Although our results showed that its expression remained basal during infection compared to the Mock, in other photosystems, such as Arabidopsis-P. parasitica, this gene’s basal expression is sufficient to participate in JA synthesis [41].
Due to the accumulation of JA, a gene usually upregulated is VSP2, a vegetative storage protein is encoded [42]. This protein is a significant source of nutrients that accumulate in plant tissue under abiotic or biotic stress [43]. Our results showed upregulation after 3 hpi for the three isolates, then downregulation and raising at 48 hpi for CPPhZOC_01 and CPPhZC_04.
Finally, the accumulation of JA led to the expression of the plant defensin gene PDF1.2, which was upregulated at 24 hpi and raised at 72 hpi. Interestingly, upregulation mainly occurred in the isolate CPPhZCO_01.
These results suggest that A. thaliana triggers SA, JA, and ET biosynthesis during infection by P. palmivora. Furthermore, the resistance response to the CPPhZC_04 isolate might have been more closely associated with SA (PR1 and ICS1), as there was a higher expression of genes associated with the SA pathway. In contrast, genes associated with JA and ET were more upregulated in the successful infection. Therefore, they seemed to mediate a susceptible response that facilitated infection by CPPhZC_05 and CPPhZOC_01.
4. Discussion
Phytophthora palmivora has many hosts and induces diseases in several crops. To broaden the alternatives in which P. palmivora can be studied, we examined its capacity to infect the well-known plant model Arabidopsis. Three oil palm—P. palmivora—isolates were used to infect detached leaves and whole seedlings of Arabidopsis Col-0.
Our findings demonstrate that two isolates can infect detached Arabidopsis Col-0 leaves or seedling roots following a 72 h cycle. Infection begins at the biotrophic phase, characterized by cyst germination, appressorium formation, and invasive hyphae formation, and progresses to the necrotrophic phase, where sporangia are generated. This life cycle is similar to other Phytophthora species capable of infecting Arabidopsis [12,14]. Additionally, an incompatible interaction was observed in the CPPhZC_04 isolate on detached leaves and whole seedlings.
This variability provided an opportunity to increase our understanding of the differences among P. palmivora isolates collected from oil palms.
To study differences in isolates, we used a qRT-PCR approach to quantify ten previously detected P. palmivora genes involved in oil palm infection [22]. Hallmarks of the infection, Hmp1 and NmrA, were expressed during compatible infection. Furthermore, the gene MFS_1, associated with a sugar transmembrane transporter, was detected mostly at 24 hpi, 48 hpi, and 72 hpi in the isolate CPPhZC_05 when hyphae were growing and spores were developing, which is consistent with the assumption that at those stages, filamentous microorganisms increase nutrient acquisition for development [44]. Likewise, sulfate permeases (SulP) are involved in the anabolism or removal of waste. In our results, a SulP gene changed dynamically through time and between the tree isolates. Similar patterns have been seen in Phytophthora infestants [44,45], where sulfur is likely to be used to synthesize cysteine, which is important for the production of antioxidants and secretion of effectors through the sulfurization pathway [46]. Furthermore, sulfate transport has been identified as one of the important processes that could be targeted for the control of disease caused by P. infestants in tomato plants [44].
Iron is another element linked to virulence in many pathogens, and is taken up from the host. Transporting iron to pathogens often involves ferric reductases that reduce ferric iron to assimilable ferrous [47]. According to our results, P. palmivora isolates CPPhZC_05 and CPPhZOC_01 upregulated the gene FRE at late stages, reaching a peak at 48 hpi. This suggests that iron was being taken up and that it might be important for P. palmivora growth in plants. In the Arabidopsis, Phytophthora capsici, pathosystem, iron depletion caused less infection in the plant. However, its relationship with virulence has not been proven [48].
Our result showed that genes related to the uptake of nutrients are co-expressed with candidate effectors and have similar pattern expressions as those previously reported when P. palmivora (CPPhZC_05) infected oil palm [22]. Specifically, we evaluated two genes that encode apoplastic effectors (Gh17 and CXE), which contained hydrolytic domains. They were upregulated at 24 hpi when the pathogen invaded the tissue. This might suggest that Gh17 and CXE play a role in degrading compounds of plant cell walls during P. palmivora infection. In the case of Gh17, a protein belonging to the glycosyl hydrolase family 17 has an O-Glycosyl hydrolyzing activity. In other pathosystems, such as Cladosporium fulvum-tomato, genes encoding Glycosyl hydrolase family-17 proteins are upregulated by the pathogen during the later phases of infection, when tomato leaves are necrotic, and the fungus is saprophytic [49].
On the other hand, CBX is a Carboxylesterase type B [50] of the carbohydrate esterase family 1 (CE1). It catalyzes ester bonds, breaks carbon-carbon bonds, and participates in decarboxylation [51]. In plants, it helps with growth, activates hormone signals, responds to biotic stress, and breaks down waxy polymers during germination [52,53,54,55,56]. It is found in hemibiotrophic lifestyle oomycetes and is upregulated in plant pathogenic oomycetes [57]. However, the precise role of Carboxylesterases during phytopathogen infection is still unknown.
We also quantify Eli17, a secreted protein with an elicitin domain. Interestingly, according to our results, the gene Eli17 was only upregulated by the incompatible isolate CPPhZC_04, while it was downregulated in the compatible ones. It is well known that elicitins are a family of small extracellular proteins with sterol-binding activity, which are found in oomycetes such as the Phytophthora and Pythium species [58]. Elicitins are characterized by their ability to induce a hypersensitive response (HR) in Solanaceae and Brassicaceae families when they infiltrate into leaves [59]. Although the Eli17 protein sequences of the three isolates are 98% identical (Figure S1), the high amount of the transcript expressed by CPPhZC_04 could explain part of the incompatibility. In other phytopathogens such as P. parasitica and P. infestans, the low production of elicitin correlates with increased virulence in Nicotiana benthamiana [60]. However, it is beyond this study’s scope to determine the level of recognition of this elicitin by Arabidopsis.
In addition, the cytoplasmatic effectors (RXLR_40906 and RXLR_44719) were over-expressed, but the expression levels varied among isolates over time. Studies on expression profiles suggest that the regulation of RXLR effector expression is complex. It may be influenced by epigenetic mechanisms, such as RNA interference or histone methylation [61], or may be expressed depending on their functional activities; some are expressed early to suppress effector-triggered immunity (ETI), while others are expressed later to suppress pattern-triggered immunity (PTI) [62]. Therefore, these two genes may play distinct roles during infection. Concerning the response of Arabidopsis against P. palmivora, our findings indicate that SA, JA, and ET undergo regulation during the interaction. The induction of SA seems to be the primary resistance mechanism against P. palmivora. While the early induction of JA and ET impairs resistance, leading to a susceptibility state, it is widely known that a crosstalk between SA and JA signaling pathways occurs to adapt to infection by biotrophic and/or necrotrophic pathogens [63]. For instance, studies into the overexpression of the gene AtRTP5 in Arabidopsis resulted in a higher accumulation of JA and lower SA production than wild-type plants (Col-0). Consequently, the mutants exhibited susceptibility to P. parasitica [41]. Hence, we hypothesize that Arabidopsis’ resistance against CPPhZC_04 is similar to what has been observed in P. infestans [24], P. sojae [62], and P. parasitica [12], where SA is produced in the initial stages to prevent infection.
It is worth noting that we utilized detached leaves to quantify genes. Although we found similar susceptibilities in whole seedlings, we could not assert that the same molecular response would be generated in the roots. As reported in other studies, the resistance in roots differs from that in leaves [64,65,66,67].
Here, we provide initial insight into the interaction of P. palmivora and Arabidopsis Col-0. We found isolates with compatible and noncompatible interactions with Arabidopsis. We showed that detached Arabidopsis leaves are suitable for studying P. palmivora infection and the gene expression profiles of both organisms. The next steps involving this model will include transcriptomics and genomic analysis to elucidate differences in effector expression among the isolates of P. palmivora and Arabidopsis responses.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mohamed Azni I.N.A. Sundram S. Ramachandran V. Pathogenicity of Malaysian Phytophthora palmivora on cocoa, durian, rubber and oil palm determines the threat of bud rot disease For. Pathol.201949 e 1255710.1111/efp.12557 · doi ↗
- 2Torres G.A. Sarria G.A. Martinez G. Varon F. Drenth A. Guest D.I. Bud Rot Caused by Phytophthora palmivora: A Destructive Emerging Disease of Oil Palm Phytopathology 201510632032910.1094/PHYTO-09-15-0243-RVW 26714102 · doi ↗ · pubmed ↗
- 3Sundram S. Intan-Nur A.M.A. South American Bud rot: A biosecurity threat to South East Asian oil palm Crop Protect.2017101586710.1016/j.cropro.2017.07.010 · doi ↗
- 4Sarria G. Martinez G. Varon F. Drenth A. Guest D. Histopathological studies of the process of Phytophthora palmivora infection in oil palm Eur. J. Plant Pathol.2016145395110.1007/s 10658-015-0810-9 · doi ↗
- 5Fawke S. Doumane M. Schornack S. Oomycete Interactions with Plants: Infection Strategies and Resistance Principles Microbiol. Mol. Biol. Rev.20157926328010.1128/MMBR.00010-1526041933 PMC 4468149 · doi ↗ · pubmed ↗
- 6Sarria G. Garcia A. Mestizo Y. Medina C. Varon F. Mesa E. Hernandez S. Antagonistic interactions between Trichoderma spp. And Phytophthora palmivora (Butler) from oil palm in Colombia Eur. J. Plant Pathol.202116175176810.1007/s 10658-021-02363-z · doi ↗
- 7Tupaz-Vera A. Ayala-Diaz I.M. Rincon V. Sarria G. Romero H.M. An integrated disease management of oil palms affected by bud rot results in shorter recovery times Agronomy 202111199510.3390/agronomy 11101995 · doi ↗
- 8Boyd L.A. Ridout C. O’Sullivan D.M. Leach J.E. Leung H. Plant–pathogen interactions: Disease resistance in modern agriculture Trends Genet.20132923324010.1016/j.tig.2012.10.01123153595 · doi ↗ · pubmed ↗