Prevalence and Diversity of Trypanosoma cruzi in Triatomine Vectors and Their Blood Meal Sources from South Central Texas, USA
Rebecca J. Kilgore, Trina Guerra, Heather Beck, Andrea Villamizar Gomez, Michael R. J. Forstner, Dittmar Hahn

TL;DR
This study explores how triatomine bugs in Texas feed on various animals and how far they travel, which helps understand the spread of Chagas disease.
Contribution
The study provides novel insights into the foraging distances of triatomine vectors and their diverse blood meal sources in Texas.
Findings
Triatomine bugs in Texas fed on a wide range of hosts, including raccoons, humans, and chickens.
Vector foraging distances exceeded 400 meters, indicating potential for wide pathogen spread.
Trypanosoma cruzi was detected in 59% of triatomine insects tested.
Abstract
Chagas disease is endemic to the state of Texas in the United States but does not have consistent surveillance or reporting. We utilized multiple sampling sites and different species of triatomine to gain data on the blood meal sources found by DNA testing for the host and vector species identities. From domestic, peridomestic, and rural sites, we found a breadth of blood meal origins including mammals, chickens, and reptiles. Unique non-native taxa utilized for blood meals enabled us to also report on extensive foraging distances for the vectors. Understanding the diversity of blood meal sources and the distances the vectors travel between meals and daytime refuges are both important aspects for understanding the spread of this disease. The prevalence of Trypanosoma cruzi was assessed in 117 triatomine insects from central Texas. The qPCR-based results revealed T. cruzi in 59% of the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
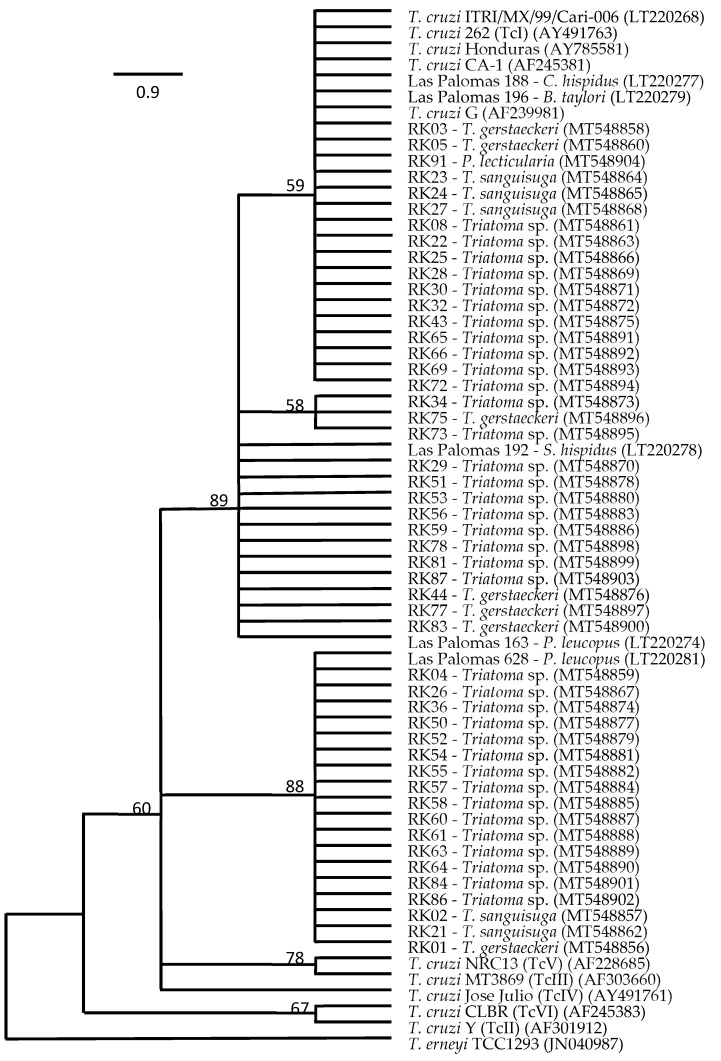 Figure 1
Figure 1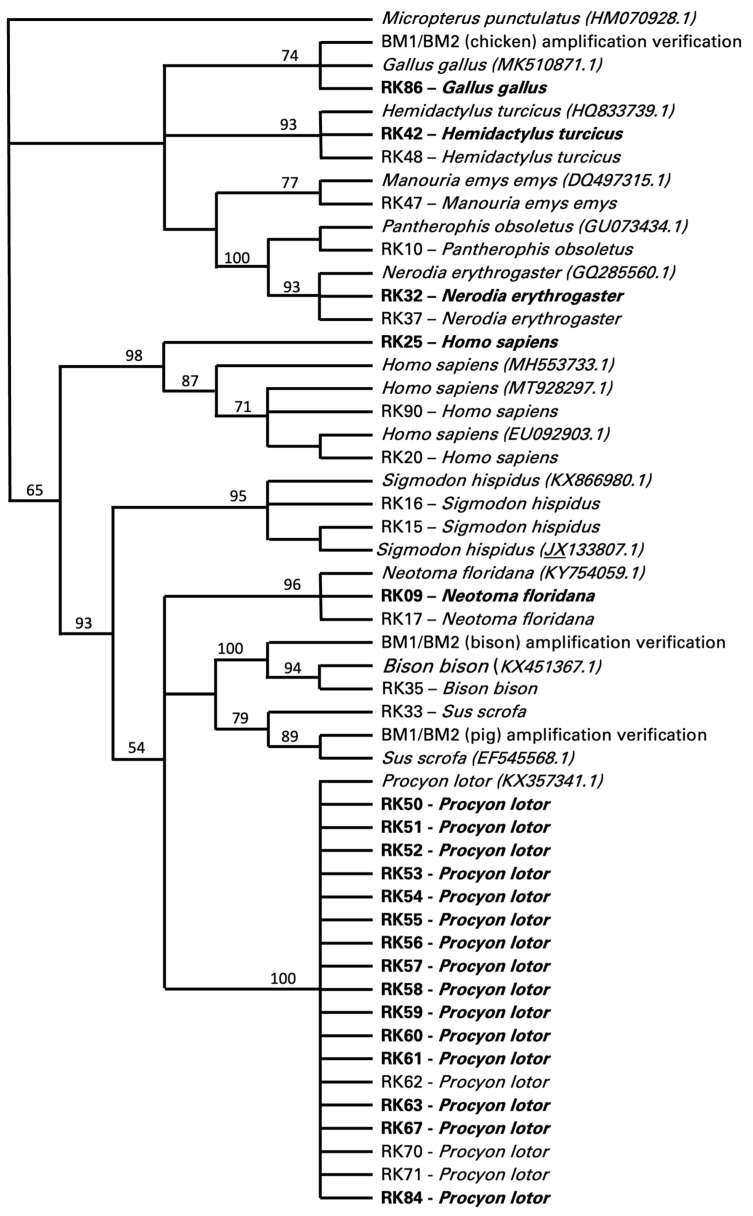 Figure 2
Figure 2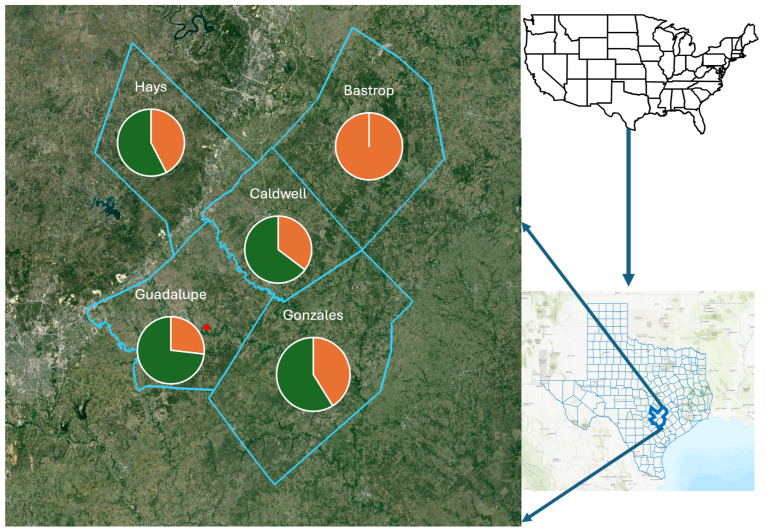 Figure 3
Figure 3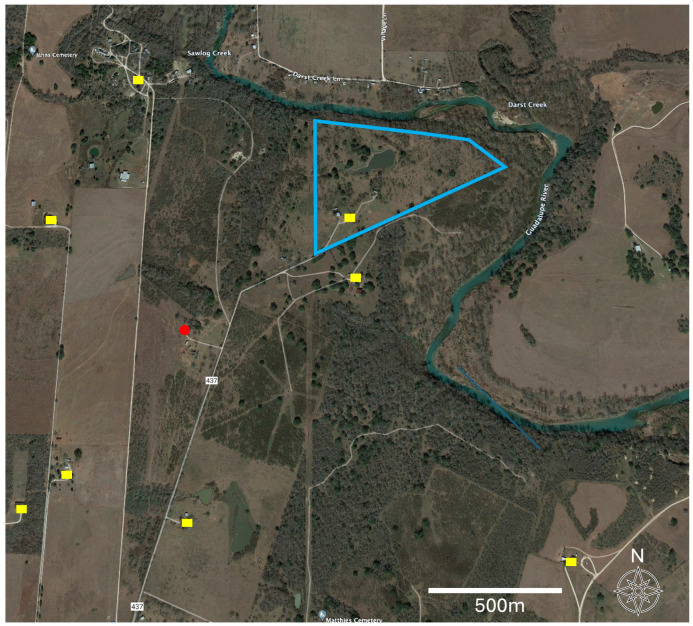 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTrypanosoma species research and implications · Research on Leishmaniasis Studies
1. Introduction
Chagas disease is one of many neglected tropical diseases shared across North, Central, and South America [1,2]. The causative agent of Chagas disease is Trypanosoma cruzi, a parasitic protozoan that has been detected in many mammalian hosts including domestic and wild animals such as rodents, opossums, raccoons, armadillos, bats, dogs, cats, goats, and pigs [2,3,4,5,6]. The parasite is naturally transmitted to mammals through infected triatomine fecal material introduced into a wound but also by oral ingestion or congenital routes [7].
In central Texas, a region with established populations of infected triatomine vectors [8,9,10,11], 70% of raccoons were found to be positive for T. cruzi, while other animals such as bobcats, ocelots, coyotes, and foxes revealed much a lower prevalence of 14% each [3,12]. T. cruzi infections were also reported from feral hogs (Sus scrofa), even though the prevalence was relatively low at 6% [13]. The Triatomine vector species in Texas have mainly been identified as Triatoma gerstaeckeri Stål 1859, though occasionally Triatoma sanguisuga Leconte 1855, Paratriatoma lecticularia Stål 1859, Triatoma rubida Uhler 1894, or Triatoma protracta woodi Uhler 1894 were observed as well [8,9,10,14]. Up to 50% and more of the insects have been reported to be infected with T. cruzi [8,10,14], with adult insects generally 5 to 10 times more likely to be infected than nymphs [10,15].
The diversity of potential blood meal source prey may be poorly characterized in Texas compared to the potential breadth of prey available. Similarly, while previous work has characterized the prevalence of T. cruzi detection in triatomines, concurrent tests for blood meal sources for the vectors are less frequently reported [8,14]. We sought to address some of these knowledge gaps for central Texas, USA. The goal of this study was to assess the prevalence of T. cruzi in triatomines in South Central Texas from a variety of sampling locations, characterize triatomine vectors, and subsequently determine their blood meal sources.
2. Materials and Methods
Triatomine insect specimen collection and preparation. Triatomine insects were iteratively collected opportunistically within five central Texas municipal counties (Bastrop, Caldwell, Gonzales, Guadalupe, and Hays), generally from peridomestic sites surrounding buildings and woodpiles of residential structures, except for two insects in Hays County and four insects in Caldwell County that were captured inside residences. We characterized these sites by the density of human habitation, alongside the captures (n = 6) within a home or outside it: domestic (>100 homes/km^2^), peridomestic (>10 homes/km^2^), and rural (<1/km^2^). Active search efforts were not made in non-disturbed or sylvatic environments. Additional triatomines were collected from peridomestic sites using intentionally placed wooden lumber debris piles outside a barn in Guadalupe County, Texas, or as bycatch in a sylvatic herpetofaunal study using pitfall arrays with associated bucket traps on the rural Griffith League Ranch in Bastrop County, Texas. Three adults of Arilus cristatus (wheel bug) and two nymphs of Pyrrhocoris apterus (fire bug) were used as negative controls for DNA-based analyses of T. cruzi and triatomine identification. Insect specimens were stored at −20 °C until extractions were performed on all available tissue recovered from the abdominal contents (about 25 mg) using the Qiagen DNeasy Blood and Tissue Kit (Qiagen, Germantown, MD, USA), following the instructions for tissue samples. DNA was resuspended in a final volume of 200 µL [8].
Detection of T. cruzi. The presence of T. cruzi in triatomine samples was assessed by qPCR targeting a 166 bp fragment of satellite DNA with primers Cruzi 1 and Cruzi 2 (Table 1) [16]. Standards used for qPCR reactions were PCR amplified products from T. cruzi ITRI/MX/99/Cari-006 originally isolated from Triatoma picturata and representing lineage TcI, obtained from Universidad Autonoma del Estado de Morelos, Mexico [17]. All DNA samples were diluted ten-fold for amplification and analyzed in triplicate. qPCRs were performed on an Eco Real-Time PCR System (Illumina, San Diego, CA, USA) in a 10 µL volume reaction with 5 µL SYBR^®^ Green (SsoAdvanced Universal SYBR^®^ Green Supermix), 0.2 µL of 10 µM Cruzi primer 1, 0.2 µL of 10 µM Cruzi 2 primer, 3.6 µL H_2_O, and 1 µL of DNA solution (Bio-Rad, Hercules, CA, USA). The qPCR parameters consisted of initial denaturation at 95 °C for 5 min, followed by 40 cycles at 95 °C for 15 s and 58 °C for 60 s (Table 1) [8].
Identification of T. cruzi. Samples with positive qPCR detection of T. cruzi using primers Cruzi 1 and Cruzi 2 were used to amplify a 667 bp fragment of the 18S rRNA gene of T. cruzi in a nested PCR, initially with primers of SSU4F and 18Sq1R (followed by using the amplicons as a template for primers SSU561F/SSU561R [19]. The initial amplicons were generated on a PTC-200 DNA Thermocycler (MJ Research, Watertown, MA, USA) of 25 µL with 1 µL of specimen sample, 12.5 µL of Green Taq polymerase, 0.8 µL of 10 µM SSU4F, 0.8 µL of 10 µM 18Sq1R, and 9.9 µL of nuclease-free H_2_O. PCR reaction parameters consisted of initial denaturation at 95 °C for 5 min, followed by 35 cycles at 95 °C for 30 s, 50 °C for 45 s, and 72 °C for 60 s (Table 1). Amplicons were cleaned using a PCR clean-up kit following standard protocols (Promega, Madison, WI, USA). The nested PCR reaction was performed with the same conditions as those listed above, with the exception of an increase in DNA template to 2 µL and a corresponding decrease in water to 8.9 µL.
Amplicons of the nested PCR were visualized on a 2% agarose gel, cleaned using Shrimp Alkaline Phosphate and Exonuclease I (Affymetrix, Santa Clara, CA, USA) following the manufacturer’s protocols, and then sequenced bidirectionally using BigDye Terminator v3.1 (Applied Biosystems, Foster City, CA, USA), with the same primers used for PCR. Sequences were analyzed on a 3500 Genetic Analyzer (Life Technologies, Carlsbad, CA, USA) and deposited at GenBank/EMBL under accession numbers MT548855-MT548904.
Identification of Triatomines. Triatomine insects were identified by comparative sequence analyses of partial cytochrome B or COI amplicons of mitochondrial DNA [8,27] using DNA extracts from insect intestinal samples. All COI results were obtained using identical primers and methods in our previous work [8] and utilized the same specimens. The cytochrome B amplicons reactions were performed on a PTC-200 DNA Thermocycler of 25 µL in volume with 1 µL of DNA template, 14.25 µL of nuclease-free H_2_O, 4.125 µL of 15 mg mL^−1^ BSA (Thermo Fisher Scientific, Waltham, MA, USA), 2.5 µL of 10× Taq Buffer, 0.5 µL of 10 µM each of 7432F forward primer and 7433R reverse primer (Table 1), 0.5 µL of 10 mM dNTP stock, 1.5 µL of 50 mM MgCl_2_, and 0.125 µL of 5 U µL^−1^ Taq polymerase [27]. Thermocycler parameters were 95 °C for 5 min followed by 34 cycles of 95 °C for 30 s, 45 °C for 45 s, 72 °C for 60 s, and, finally, 72 °C for 10 min [28] (Table 1). Amplicons were cleaned and sequenced bidirectionally using BigDye Terminator v3.1 and sequences were analyzed on a 3500 Genetic Analyzer as described above.
Identification of food sources. Blood meals of triatomines were analyzed from insect intestine samples by comparative sequence analyses of partial cytochrome B amplicons of mitochondrial DNA, with three different primer sets targeting vertebrates in general, mammals, or birds, respectively (Table 1) [22]. DNA extracted from tissue samples of bison, chicken, swallow, turkey, alligator, rat, mouse, turtle, and pig tissues were used as positive controls for these primer sets. While the primer set for mammals amplified all controls, the vertebrate primer set failed to detect turtles, and the avian primer set detected swallow, turkey, alligator, mouse, and rat but did not amplify chicken. Reactions were performed on a PTC-200 DNA Thermocycler of 25 µL with 3 µL of DNA template, 12.175 µL of nuclease-free H_2_O, 4.125 µL of 15 mg mL^−1^ BSA, 2.5 µL of 10× Taq Buffer, 0.5 µL of 10 µM each of forward primer and reverse primer (Table 1), 0.5 µL of 10 mM dNTP stock, 1.5 µL of 50 mM MgCl_2_, and 0.25 µL of 5 U µL^−1^ Taq polymerase. Amplicons were cleaned and sequenced bidirectionally using BigDye Terminator v3.1 and sequences were analyzed on a 3500 Genetic Analyzer as described above.
Phylogenetic Analyses. Sequences were assembled in Geneious 8.1.8 (Biomatters Ltd., Auckland, New Zealand) and checked in the GenBank/EMBL databases using the BLAST algorithm [29]. The identities and relationships among the sequences amplified were compared to available GenBank sequences using Neighbor-Joining (NJ) [30] and maximum likelihood (ML) [31] analyses, including bootstrap consensus for 1000 replicate analyses [32,33]. NJ trees used the HKY substitution method and 1000 bootstrap replicates. All trees included the percent consensus support from ML analysis where appropriate. Trees were generated to identify the strain identity of the T. cruzi sequences as well as phylogenetic results from the mitochondrial DNA sequences that enabled species identification of the insect vector.
3. Results
A total of 117 insects were classified as triatomines by sight identification, with 90 individuals characterized as adults and 27 as nymphs (Table 2). Further determination of instar stage was not performed. Morphological keys could not enable species-level determination because the frozen, individually stored specimens were in flexible plastic bags and experienced considerable damage to appendages and color changes attendant to such storage. Table 2 also includes arthropods we previously identified [8]. Those samples had been previously identified as species using the same methods as in this project but had not had blood meal assessments [8]. Those 30 insects were included in the blood meal assessments here. In 59% (70/117) of these insects (62 adults and eight nymphs), qPCR-based analyses indicated the presence of T. cruzi (Table 2). The prevalence of T. cruzi detected in these insects was 59% (19/33), 73% (30/41), 64% (11/17), 58% (10/17), and 0% (0/9), respectively, for the Hays, Guadalupe, Caldwell, Gonzales, and Bastrop counties (Table 2). Two of the five non-triatomines, i.e., one wheel bug and one fire bug, used as negative controls showed weak amplification, with Ct values close to the detection limit, while the remaining two wheel bugs and one fire bug did not show any amplification.
Comparative sequence analyses of a 667 bp fragment of the 18S rRNA gene targeting T. cruzi in a nested PCR allowed us to generate sequences from T. cruzi in 49 of the 70 samples, with T. cruzi from 31 triatomine insects from Hays, Guadalupe, and Caldwell counties clustering with representatives of Tc1 and the remaining related to Tc1, but distinct to TcII to TcVI (Figure 1). The presence of T. cruzi in our non-triatomine insects, i.e., the wheel bug and the fire bug, was not confirmed as T. cruzi by sequence data and thus was characterized as false-positive qPCR detections. Comparative sequence analyses of the 18S rRNA gene fragment targeting T. cruzi revealed a 98.9% pairwise identity match to Trypanomastidae sp. isolate PNG85 (MK929454) for the wheel bug sample and a 99.8% pairwise identity match to Blastocrithidia papi isolate Pa3 (KX641340) for the fire bug sample. Another sample, RK35, with a strong qPCR signal indicating the presence of T. cruzi, was also identified as a false-positive detection, with a 99.4% pairwise identity match to the same Blastocrithidia papi isolate Pa3 (KX641340). Table 3 provides the results for triatomine species, T. cruzi status, and Texas county of origin. Twenty-one of the triatomine insects positive for T. cruzi were identified as Triatoma gerstaeckeri by comparative CytB sequence analyses, with a 99.7 to 100% pairwise identity match to the sequence of specimen San Marcos I-13 (LT630441), 100% pairwise identity match to the sequence of I-JAM02 (LT630443), or a 98.2% pairwise identity match to the sequence of Parker 026 (KY305701). Twenty-five other individuals, negative for T. cruzi, were also identified as T. gerstaeckeri. Eight insects carrying T. cruzi and another five insects without T. cruzi were identified as Triatoma sanguisuga, with a 99.1 to 99.7% pairwise identity to the sequence of specimen PS099 (KY305708). Two adult insects, both positive for T. cruzi, were identified as Paratriatoma leticularia, with a 98.6 to 99.1% pairwise identity match to the sequence of specimen PS100 (KY305709). The remaining 39 insects positive for T. cruzi could not be identified at the species level and were further classified as Triatoma species (Figure 1).
Food sources could only be determined in 34 of the 117 specimens, 20 of which were positive for T. cruzi and 14 were negative for T. cruzi (Figure 2). These data were all retrieved with the cytB primer set targeting vertebrates, with a few also confirmed by the cytB primer set targeting mammals. The cytB primer set designed to target only avian samples did not produce any results. The cytB primer set targeting vertebrates also retrieved non-target sequences from twenty-six insects (i.e., 23 sequences identifying Triatoma sp. and 3 sequences identifying Gryllus sp.). Of the food sources identified as vertebrates, 53% (18/34) were identified as Procyon lotor (raccoon), with 15 individuals positive and 3 negative for T. cruzi (Figure 2). Other food sources included Homo sapiens (human, 9%, 3/34), with one insect positive and two insects negative for T. cruzi; Hemidactylus turcicus (house gecko, 6%, 2/34), Neotoma floridana (Eastern woodrat, 6%, 2/34), and Nerodia erythrogaster (plain-bellied water snake, 6%, 2/34), all with one insect positive for T. cruzi and one negative; and Sigmodon hispidus (hispid cotton rat, 6%, 2/34), with two insects negative for T. cruzi (Figure 2). One insect feeding on Gallus gallus (chicken, 3%, 1/34) was positive for T. cruzi, while insects feeding on Manouria emys emys (Asian forest tortoise, 3%, 1/34), Bison bison (bison, 3%, 1/34), and Sus scrofa (pig, 3%, 1/34) were negative for T. cruzi (Figure 2).
Several of these blood meal taxa provided serendipitous information for the foraging distances of these vector insects. Specifically, the detection of these blood meals was novel among reported hosts for the vector insects, and detecting them enabled the calculation of the minimum distances between the blood meal source and the arthropod collection site using straight-line measurement. First, the Asian forest tortoise (Manouria emys) was captively held in an indoor/outdoor paddock. The minimum measured distance between the corner of that paddock and the insect vector collection site was 40 m. However, the tortoise could move as much as 110 m from the site of the vector. Next, a blood meal was detected from bison (Bison bison). These animals were held in a large, fenced facility that was a 600 m minimum distance from the insect vector location in the corner of that facility, proximal to the collection site. Importantly, this was the minimum distance, and this facility enabled the bison to travel as far as 1210 m from the site of insect collection. Finally, we also detected three instances of humans as the blood meal; one of these vector insects was also positive for T. cruzi. There were no human-occupied buildings, campsites, or other such occupancies near the vector collection site. The nearest potential occupied location was 430 m from the collection site, but other alternative locations were found at distances of 621–698 m (n = 5 sites). These all represent neighboring tracts of land adjacent to the insect collection location.
4. Discussion
Trypanosoma cruzi was detected in 59% of the 117 triatomine insects (76% adults and 23% nymphs), with 68% of the adults and 29% of the nymphs being positive. These results are in agreement with those of previous studies in Central Texas in which 50% or more of the triatomine insects analyzed were infected with T. cruzi [8,10,11,14,33]. A higher prevalence in adult insects compared to nymphs has also been reported [10,15], even though one study reported a much lower overall prevalence of 23% in adults and 4% in nymphs compared to our results [15]. High prevalence in triatomines was also found in other Texas areas [6], and the prevalence in triatomines collected in Texas was generally higher than in other states, including Oklahoma [34], Nebraska [35], or specific areas in Arizona, while other areas in Arizona were comparable to Texas values [36].
Triatomines were predominantly collected from peridomestic habitats in five counties in South Central Texas (Figure 3), with similar prevalence values for T. cruzi in four counties (Hays, Guadalupe, Caldwell, Gonzales), with 57% (19/33), 73% (30/41), 64% (11/17), and 58% (10/17), respectively, while T. cruzi was not detected in samples from Bastrop County (0/9). While prevalence values for Hays, Guadalupe, Caldwell, and Gonzales counties represented values generally found in Texas, the lack of any detection in neighboring Bastrop County was unexpected, and most likely a consequence of the small sample size. A small sample size might also have been the cause of the low prevalence detection in triatomes from Nebraska [35], but this was not considered to explain the samples from Arizona [36]. Here, the sample size was comparable, but the sites differed with respect to the most frequently encountered triatomine species, Triatoma rubida and Triatoma recurva, respectively [36].
The presence of T. cruzi detected by qPCR-based analyses was confirmed by comparative sequence analyses of 18S rRNA gene fragments in 87% of the samples, with most of the sequences clustering with those of representatives of discrete typing unit (DTU) TcI [37] and the remaining related to TcI, but distinct from DTUs TcII to TcVI. DTU TcI is widely distributed in the Americas and found in Texas in Triatoma sp. from domestic and sylvatic areas [2,10,15]. Our phylogenetic analyses clearly identified members of the TcI lineage but could not explicitly characterize the remaining T. cruzi lineage with bootstrap support, even though underlying comparative raw sequence analyses showed a high similarity to TcIV for these isolates. Thus, while our analyses largely corroborate data on TcI lineage abundance in triatomes in Texas, the potential occurrence of TcIV in triatomines necessitates additional informative DNA sites that can enable a better-supported resolution of T. cruzi lineages found in our study.
Vectors mainly belonged to Triatoma gerstaeckeri (35% of which 65% were positive for T. cruzi), Triatoma sanguisuga (21%, 43% positive), and Paratriatoma leticularia (0.3%, 100% positive); however, 56% of the insects could not be identified at the species level using molecular tools. Of those identified, T. gerstaeckeri was confirmed as the most prominently encountered triatomine insect in Central Texas in our study, with T. sanguisuga and P. lecticularia detected much less frequently, as described in other studies [8,9,10,14,38]. Studies on triatomines in other southern states retrieved T. rubida and T. recurva (Arizona) [36], T. sanguisuga (Oklahoma, Nebraska) [34,35], or T. rubida (New Mexico, West Texas) [6] as the most commonly encountered triatomine insects, though studies often dealt with domestic rather than peridomestic or rural habitats.
Food sources were determined in 29% (34/117) of the insects. This was a lower blood meal recovery rate than two other studies from Texas where the recovery was 92% [9] and 63% [22]. The rate of blood meal recovery success has been shown to decline with the satiation state of the vector insect [22] and when dead arthropods as opposed to living individuals are dissected [22]. It is possible that our generally rural sites provided fewer opportunities for regular meals by the insects, decreasing their satiation and our recovery rate. All of our arthropods were frozen and not living prior to dissection. For the blood meals recovered in this study, the majority of hosts were raccoons (P. lotor), which were identified in 42% (15/34) of these blood meals. With 83% (15/18) of those samples being positive for T. cruzi, the results were higher for raccoons than those reported from other states, with 34% and 43% [39,40]. These results are closer to those obtained for Texas in other studies with prevalence values of 62% and 70% [3,41]. In Texas, DTU TcIV has been identified as the sole T. cruzi lineage in raccoons [3,41], with one TcI/TcIV mixed infection as an exception [3]. Comparative raw sequence analyses in our study supported these results since T. cruzi from raccoons showed a high similarity to sequences identifying the TcIV lineage. Blood meal sources included humans, though expectedly represented a much lower prevalence among blood meals at 9% (33% positive) in our peridomestic habitats than was detected in domestic environments, providing up to 67% [9,15,42]. We detected a blood meal from an Asian forest tortoise (Manouria) without T. cruzi infection that was held at the location but contained in an enclosure 40–110 m from the arthropod capture site. Further, we noted that for one of the peridomestic samples, while the blood meal was human, the nearest domicile occupied by people at the time of the sampling was over 400 m away (Figure 4). Finally, we also detected bison as a blood meal source though without T. cruzi infection. Bison were held in paddocks spanning 600–1200 m from the sampling site, indicating the high mobility of triatomine insects (Figure 4). These last two serendipitous blood meal detections uniquely enabled us to report very large distances between the source of the blood meal to the subsequent Triatoma sp. capture sites. These foraging distances for the insect vectors indicate that a better understanding of the insect vector movement ecology is critical to understanding the potential for exposure and the modeling of this emerging disease.
5. Conclusions
Our main goals for this project were to collect and identify both the triatomines potentially acting as vectors for Trypanosoma cruzi and further contribute to characterizing the blood sources that these insects use for meals in Texas. We were successful in triatomine sampling, with detections in all five counties in Central Texas. Trypanosoma cruzi was routinely detected among the potential triatomine vectors. Two species of Triatoma and one species of Paratriatoma were identified as positive for T. cruzi. The majority of blood meals were identified to have come from raccoons (Procyon lotor), but a wide array of vertebrate blood meals were confirmed including three samples of Homo sapiens. One of the human blood meals was also from a vector positive for T. cruzi. We were not able to successfully identify 44% (51/117) of the vector insects at the species level; simply storing individual specimens within freezer vials until laboratory processing would likely have prevented this difficulty.
The predominant sampling sites varied from peridomestic to rural, and the samples’ unique, non-native sources for the blood meals enabled us to infer foraging distances for the triatomine vectors. In one case this confirmed a 600 m minimum movement distance for the Triatomine from the blood meal to the daytime retreat where the insect was captured. We considered the variety of blood meal sources and the uniquely enabled forage distance measurements for the Triatoma sp. to be particularly relevant for modeling or assessments of the vector side of T. cruzi evaluations. However, these were a limited set of forage distances; moreover, they are unlikely to be the largest distances that occur. In Texas, but not only Texas, future research into these distances will be enabled because of the wide array of captive, enclosed exotic mammals and reptiles held privately across the landscape of the state. The occurrence of these non-native species could be leveraged by future research to reveal additional Triatomine dispersal/foraging data by conducting experiments collecting the vectors at known distances from unique exotic species enclosures.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bern C. Messenger L.A. Whitman J.D. Maguire J.H. Chagas disease in the United States: A public health approach Clin. Microbiol. Rev.201933 e 00023-1910.1128/CMR.00023-1931776135 PMC 6927308 · doi ↗ · pubmed ↗
- 2Bern C. Kjos S. Yabsley M.J. Montgomery S.P. Trypanosoma cruzi and Chagas’ disease in the United States Clin. Microbiol. Rev.20112465568110.1128/CMR.00005-1121976603 PMC 3194829 · doi ↗ · pubmed ↗
- 3Curtis-Robles R. Lewis B.C. Hamer S.A. High Trypanosoma cruzi infection prevalence associated with minimal cardiac pathology among wild carnivores in central Texas Int. J. Parasitol. Parasites Wildl.2016511712310.1016/j.ijppaw.2016.04.00127330982 PMC 4900435 · doi ↗ · pubmed ↗
- 4Munoz-San Martin C. Campo Verde Arbocco F. Saavedra M. Actis E.A. Rios T.A. Abba A.M. Morales M.E. Cattan P.E. Jahn G.A. Superina M. High rates of Trypanosoma cruzi infection in goats from Mendoza province, Argentina: Parasite loads in blood and seasonal variation Acta Trop.202020810549310.1016/j.actatropica.2020.10549332371222 · doi ↗ · pubmed ↗
- 5Herrera L. D’Andrea P.S. Xavier S.C. Mangia R.H. Fernandes O. Jansen A.M. Trypanosoma cruzi infection in wild mammals of the National Park ‘Serra da Capivara’ and its surroundings (Piaui, Brazil), an area endemic for Chagas disease Trans. R. Soc. Trop. Med. Hyg.20059937938810.1016/j.trstmh.2004.07.00615780345 · doi ↗ · pubmed ↗
- 6Rodriguez F. Luna B.S. Calderon O. Manriquez-Roman C. Amezcua-Winter K. Cedillo J. Garcia-Vazquez R. Tejeda I.A. Romero A. Waldrup K. Surveillance of Trypanosoma cruzi infection in triatomine vectors, feral dogs and cats, and wild animals in and around El Paso County, Texas, and New Mexico P Lo S Negl. Trop. Dis.202115 e 000914710.1371/journal.pntd.000914733600455 PMC 7924784 · doi ↗ · pubmed ↗
- 7Coura J.R. Dias J.C. Epidemiology, control and surveillance of Chagas disease: 100 years after its discovery Mem. Inst. Oswaldo Cruz.2009104(Suppl. 1)314010.1590/S 0074-0276200900090000619753455 · doi ↗ · pubmed ↗
- 8Aleman A. Guerra T. Maikis T.J. Milholland M.T. Castro-Arellano I. Forstner M.R.J. Hahn D. The prevalence of Trypanosoma cruzi, the causal agent of Chagas disease, in Texas rodent populations Ecohealth 20171413014310.1007/s 10393-017-1205-528091763 · doi ↗ · pubmed ↗