A clinical metagenomic study of biopsies from Mexican endophthalmitis patients reveals the presence of complex bacterial communities and a diversity of resistance genes
Miguel Ángel Vences-Guzmán, Martín Jiménez-Rodríguez, Luis Lozano, Sergio Rojas-Juárez, Juan Abel Ramírez-Estudillo, Ángeles Yahel Hernández-Vázquez, Ingrid Yazmín Pita-Ortiz, Karol Guadalupe Ramírez-Ceballos, Silvia Medina-Medina, Christian Sohlenkamp

TL;DR
This study used metagenomic sequencing to find diverse bacteria and antibiotic resistance genes in eye infections of Mexican patients, challenging the belief that only a few bacteria are involved.
Contribution
The study reveals complex bacterial communities and resistance genes in endophthalmitis biopsies using metagenomic sequencing.
Findings
Metagenomic analysis identified a wide variety of bacteria in endophthalmitis biopsies.
Resistome analysis detected resistance genes for ceftazidime, vancomycin, and efflux pumps.
Findings contradict the assumption that only a few bacterial strains cause endophthalmitis.
Abstract
Infectious endophthalmitis is a severe ophthalmic emergency. This infection can be caused by bacteria and fungi. For efficient treatment, the administration of antimicrobial drugs to which the microbes are susceptible is essential. The aim of this study was to identify micro-organisms in biopsies of Mexican endophthalmitis patients using metagenomic next-generation sequencing and determine which antibiotic resistance genes were present in the biopsy samples. In this prospective case study, 19 endophthalmitis patients were recruited. Samples of vitreous or aqueous humour were extracted for DNA extraction for metagenomic next-generation sequencing. Analysis of the sequencing results revealed the presence of a wide variety of bacteria in the biopsies. Resistome analysis showed that homologues of antibiotic resistance genes were present in several biopsy samples. Genes possibly conferring…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
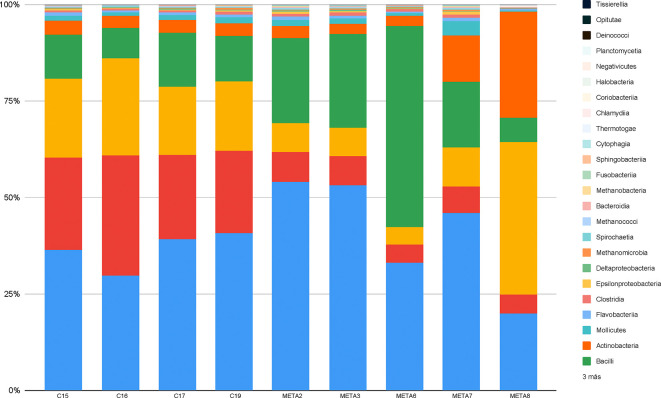 Fig. 1
Fig. 1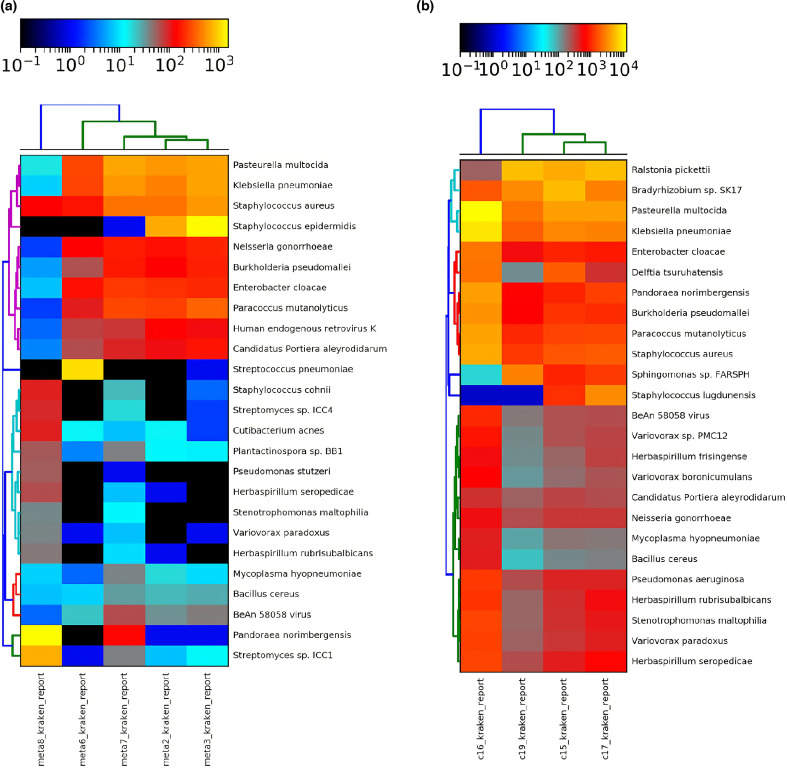 Fig. 2
Fig. 2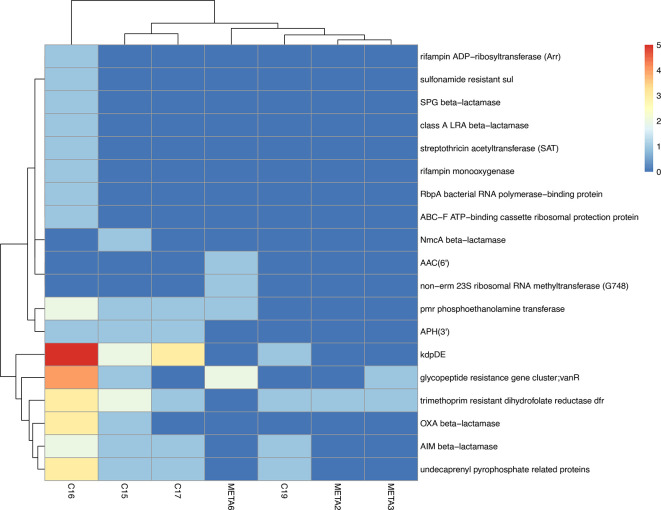 Fig. 3
Fig. 3| No. | Sex | Eye | Age (years) | Aetiologies | Initial VA | Time of symptoms(days) | Final VA | IVI (No.) | Antibiotic | Complementarytreatment | Sample | Most abundant bacteria identified by mNGS |
| 1 | M | R | 44 | Penetrating keratoplasty | HM | 6 | NLP | 2 | Ceftazidime+vancomicin | Evisceration | Vitreous | Not sequenced |
| META2 | M | R | 79 | Corneal ulcer | LP | 1 | LP | 6 | Ceftazidime+vancomicin | None | Vitreous |
|
| META3 | M | L | 71 | Intravitreal injection | LP | 7 | HM | 2 | Moxifloxacin | Vitrectomy | Vitreous |
|
| 4 | F | R | 63 | Cataract surgery | LP | 30 | 20/20 | 2 | Ceftazidime+vancomicin | Vitrectomy | Vitreous | Not sequenced |
| 5 | M | L | 48 | Vitrectomy | LP | 3 | LP | 1 | Moxifloxacin | Vitrectomy | Vitreous | Not sequenced |
| META6 | M | R | 38 | Open wound | HM | 6 | 20/400 | 2 | Moxifloxacin | Wound closure | Vitreous |
|
| META7 | M | R | 51 | Cataract surgery | LP | 30 | 20/70 | 2 | Ceftazidime+vancomicin | Vitrectomy | Vitreous |
|
| META8 | M | L | 87 | Cataract surgery | 20/40 | 4 | 20/30 | 1 | Moxifloxacin | None | Vitreous |
|
| 9 | M | L | 83 | Intravitreal injection | CF | 4 | 20/200 | 2 | Moxifloxacin | Phacovitrectomy | Vitreous | Not sequenced |
| 10 | M | L | 35 | Corneal ulcer | LP | 3 | CF | 2 | Moxifloxacin | None | Vitreous | Not sequenced |
| 11 | M | R | 82 | Cataract surgery | LP | 3 | CF | 1 | Moxifloxacin | Vitrectomy | Vitreous | Not sequenced |
| 12 | M | R | 70 | Intravitreal injection | 20/80 | 1 | 20/80 | 1 | Moxifloxacin | None | Vitreous | Not sequenced |
| 13 | F | L | 81 | Cataract surgery | LP | 3 | HM | 2 | Moxifloxacin | Vitrectomy | Vitreous | Not sequenced |
| 14 | M | R | 68 | Cataract surgery | HM | 7 | 20/200 | 2 | Moxifloxacin | Vitrectomy | Vitreous | Not sequenced |
| C15 | F | L | 69 | Intravitreal injection | LP | 10 | HM | 2 | Moxifloxacin | Phacovitrectomy | Vitreous | |
| C16 | F | R | 68 | Cataract surgery | LP | 1 | NLP | 1 | Moxifloxacin | None | Vitreous |
|
| C17 | M | L | 65 | Cataract surgery | CF | 6 | 20/50 | 1 | Moxifloxacin | Vitrectomy | Vitreous |
|
| 18 | M | L | 65 | Cataract surgery | CF | 6 | 20/50 | 1 | Moxifloxacin | Vitrectomy | Aqueous | Not sequenced |
| C19 | M | R | 42 | Open wound | LP | 6 | HM | 2 | Moxifloxacin | Phacovitrectomy | Vitreous |
|
| Total reads | Filtered reads | Contigs | Assembly (Mb) | Predicted genes | Classified contigs | |
| META2 | 12 556 288 | 9 918 778 | 107 336 | 37 | 13 105 | 4 157 |
| META3 | 16 001 022 | 13 349 320 | 128 614 | 44 | 16 150 | 5 749 |
| META6 | 7 131 962 | 5 390 718 | 58 883 | 22 | 8 768 | 2 933 |
| META7 | 2 282 582 | 1 585 174 | 84 756 | 30 | 12 422 | 4 806 |
| META80 | 18 880 270 | 8 929 374 | 12 944 | 3 | 2 150 | 4 949 |
| C15 | 143 536 508 | 54 014 296 | 1 714 050 | 583 | 108 786 | 45 872 |
| C16 | 191 525 902 | 76 121 928 | 1 371 027 | 1 323 | 321 403 | 86 959 |
| C17 | 135 897 938 | 52 257 342 | 1 834 288 | 592 | 111 796 | 45 122 |
| C19 | 116 364 188 | 45 113 668 | 1 265 077 | 389 | 72 102 | 32 133 |
|
|
|
|
|
|
| |||
| 5 624 | 30 320 | Proteobacteria/Alphaproteobacteria/Rhizobiales/Bradyrhizobiaceae | |
|
| 4 724 | 71 370 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
|
| 4 030 | 102 394 | Proteobacteria/Gammaproteobacteria/Pasteurellales/Pasteurellaceae |
|
| 3 068 | 108 983 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 1 864 | 11 959 | Proteobacteria/Betaproteobacteria/Burkholderiales/Comamonadaceae |
|
| 1 782 | 80 197 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 1 414 | 37 479 | Proteobacteria/Alphaproteobacteria/Rhodobacterales/Rhodobacteraceae |
|
| 1 151 | 31 727 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
|
| 1 120 | 8 093 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
| 994 | 113 763 | Proteobacteria/Alphaproteobacteria/Sphingomonadales/Sphingomonadaceae | |
|
| |||
|
| 12 412 | 144 821 | Proteobacteria/Gammaproteobacteria/Pasteurellales/Pasteurellaceae |
|
| 9 085 | 155 777 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 4 531 | 6 723 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 4 325 | 51 030 | Proteobacteria/Alphaproteobacteria/Rhodobacterales/Rhodobacteraceae |
|
| 4 038 | 24 963 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
|
| 3 518 | 42 284 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
|
| 2 502 | 50 080 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 2 443 | 16 603 | Proteobacteria/Betaproteobacteria/Burkholderiales/Comamonadaceae |
| 1 779 | 70 608 | Proteobacteria/Alphaproteobacteria/Rhizobiales/Bradyrhizobiaceae | |
|
| 1 430 | 6 723 | Proteobacteria/Betaproteobacteria/Burkholderiales/Oxalobacteraceae |
|
| |||
|
| 5 869 | 43 215 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
|
| 4 026 | 98 841 | Proteobacteria/Gammaproteobacteria/Pasteurellales/Pasteurellaceae |
|
| 3 230 | 22 24 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 2 939 | 102 706 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
| 2 875 | 13 542 | Proteobacteria/Alphaproteobacteria/Rhizobiales/Bradyrhizobiaceae | |
|
| 1 848 | 76 732 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 1 462 | 34 541 | Proteobacteria/Alphaproteobacteria/Rhodobacterales/Rhodobacteraceae |
|
| 1 340 | 11 481 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
| 1 270 | 103 007 | Proteobacteria/Alphaproteobacteria/Sphingomonadales/Sphingomonadaceae | |
|
| 1068 | 29225 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
|
| |||
|
| 5 887 | 46 946 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
| 3 256 | 14 164 | Proteobacteria/Alphaproteobacteria/Rhizobiales/Bradyrhizobiaceae | |
| 2 947 | 75 473 | Proteobacteria/Alphaproteobacteria/Sphingomonadales/Sphingomonadaceae | |
|
| 2 491 | 83 872 | Proteobacteria/Gammaproteobacteria/Pasteurellales/Pasteurellaceae |
|
| 1 900 | 88 916 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 1 219 | 65 525 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 1 009 | 30 730 | Proteobacteria/Alphaproteobacteria/Rhodobacterales/Rhodobacteraceae |
|
| 688 | 7 040 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
|
| 655 | 25 737 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
|
| 555 | 32 366 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| |||
|
| 704 | 5 054 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 574 | 23 897 | Proteobacteria/Gammaproteobacteria/Pasteurellales/Pasteurellaceae |
|
| 474 | 22 816 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 411 | 22 920 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 247 | 9 692 | Proteobacteria/Alphaproteobacteria/Rhodobacterales/Rhodobacteraceae |
|
| 220 | 10 142 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 161 | 1 943 | Proteobacteria/Betaproteobacteria/Neisseriales/Neisseriaceae |
| Human endogenous retrovirus K | 144 | 2 048 | Viruses/Ortervirales/Retroviridae |
|
| 137 | 7 926 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
| Candidatus Portiera aleyrodidarum | 120 | 8 580 | Proteobacteria/Gammaproteobacteria/Oceanospirillales/Halomonadaceae |
|
| |||
|
| 1 472 | 10 039 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 655 | 31 951 | Proteobacteria/Gammaproteobacteria/Pasteurellales/Pasteurellaceae |
|
| 650 | 31 442 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 573 | 32 303 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 354 | 13 693 | Proteobacteria/Alphaproteobacteria/Rhodobacterales/Rhodobacteraceae |
|
| 203 | 13 356 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 193 | 2 459 | Proteobacteria/Betaproteobacteria/Neisseriales/Neisseriaceae |
|
| 185 | 11 100 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
| Candidatus Portiera aleyrodidarum | 163 | 11 983 | Proteobacteria/Gammaproteobacteria/Oceanospirillales/Halomonadaceae |
| Human endogenous retrovirus K | 121 | 2 698 | Viruses/Ortervirales/Retroviridae |
|
| |||
|
| 1 155 | 42 636 | Firmicutes/Bacilli/Lactobacillales/Streptococcaceae |
|
| 266 | 12 824 | Proteobacteria/Gammaproteobacteria/Pasteurellales/Pasteurellaceae |
|
| 263 | 12 239 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 163 | 10 520 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 158 | 5 653 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 135 | 1 011 | Proteobacteria/Betaproteobacteria/Neisseriales/Neisseriaceae |
|
| 104 | 5 165 | Proteobacteria/Alphaproteobacteria/Rhodobacterales/Rhodobacteraceae |
| Human endogenous retrovirus K | 74 | 1 139 | Viruses/Ortervirales/Retroviridae |
| Candidatus Portiera aleyrodidarum | 67 | 2 752 | Proteobacteria/Gammaproteobacteria/Oceanospirillales/Halomonadaceae |
|
| 65 | 4 255 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
|
| |||
|
| 667 | 11 656 | Proteobacteria/Gammaproteobacteria/Pasteurellales/Pasteurellaceae |
|
| 566 | 9 186 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 416 | 9 613 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 272 | 4 103 | Proteobacteria/Alphaproteobacteria/Rhodobacterales/Rhodobacteraceae |
|
| 235 | 3 174 | Proteobacteria/Gammaproteobacteria/Enterobacterales/Enterobacteriaceae |
|
| 181 | 760 | Proteobacteria/Betaproteobacteria/Neisseriales/Neisseriaceae |
|
| 171 | 2 912 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
|
| 149 | 2 038 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
| Candidatus Portiera aleyrodidarum | 99 | 3 834 | Proteobacteria/Gammaproteobacteria/Oceanospirillales/Halomonadaceae |
| Human endogenous retrovirus K | 84 | 670 | Viruses/Ortervirales/Retroviridae |
|
| |||
|
| 1 566 | 88 901 | Proteobacteria/Betaproteobacteria/Burkholderiales/Burkholderiaceae |
| 735 | 27 546 | Actinobacteria/Actinobacteria/Streptomycetales/Streptomycetaceae | |
|
| 138 | 5 270 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 103 | 5 415 | Firmicutes/Bacilli/Bacillales/Staphylococcaceae |
|
| 102 | 32 414 | Actinobacteria/Actinobacteria/Propionibacteriales/Propionibacteriaceae |
| 94 | 3 583 | Actinobacteria/Actinobacteria/Streptomycetales/Streptomycetaceae | |
|
| 68 | 3 383 | Proteobacteria/Betaproteobacteria/Burkholderiales/Oxalobacteraceae |
| 60 | 5 296 | ctinobacteria/Actinobacteria/Micromonosporales/Micromonosporaceae | |
|
| 58 | 4 605 | Proteobacteria/Gammaproteobacteria/Pseudomonadales/Pseudomonadaceae |
|
| 44 | 2 694 | Proteobacteria/Betaproteobacteria/Burkholderiales/Oxalobacteraceae |
- —http://dx.doi.org/10.13039/501100006087 Dirección General de Asuntos del Personal Académico, Universidad Nacional Autónoma de México
- —http://dx.doi.org/10.13039/501100006087 Dirección General de Asuntos del Personal Académico, Universidad Nacional Autónoma de México
- —http://dx.doi.org/10.13039/501100003141 Consejo Nacional de Ciencia y Tecnología
- —http://dx.doi.org/10.13039/501100003141 Consejo Nacional de Ciencia y Tecnología
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOcular Infections and Treatments · Bacterial Identification and Susceptibility Testing · Antibiotic Resistance in Bacteria
Data Summary
The authors confirm that all supporting data, code and protocols have been provided within the article or through supplementary data files. The sequencing data were deposited at NCBI with the SRA accession number: SRP424451. Bioproject: PRJNA937315PRJNA937315. Biosamples: SAMN33401590, SAMN33401589, SAMN33401588, SAMN33401587, SAMN33401586, SAMN33401585, SAMN33401584, SAMN33401583, SAMN33401582.
Introduction
Infectious endophthalmitis is a severe ophthalmic emergency. It is a rare but serious complication of cataract surgery with an incidence of about 0.1 %, frequently leading to poor visual outcomes [14]. This infection can be caused by bacteria and fungi, and for efficient treatment, the administration of antimicrobial drugs to which the microbes are susceptible is essential. Since the publication of the Endophthalmitis Vitrectomy Study (EVS), the current recommendations for empirical therapy have not changed [5]. Many of the common causative organisms are sensitive to the broad-spectrum antibiotic combination vancomycin and ceftazidime, which is the current standard, but there are reports of cases of infectious endophthalmitis resistant to these antibiotics. According to the EVS, only 89.5 % of Gram-negative isolates are sensitive to amikacin and ceftazidime. Furthermore, the increased prevalence of methicillin-resistant bacteria has become a clinical problem. New antibiotics that can be administered intravitreally are required, and other types of antibiotics could be used to control intraocular infections [6]. One alternative antibiotic that could be used in endophthalmitis patients is the fourth-generation fluoroquinolone moxifloxacin (MXF). MXF has been tested in in vitro studies using cell culture and in animal models. Previous studies have established that 150 µg ml^−1^ MXF is a safe concentration for rabbit eyes and in vitro human pigmented cells. Intravitreal MXF caused no electroretinographic or retinal histological abnormalities [78].
The recognition of the causative pathogens of acute endophthalmitis is fundamental and can have wide-reaching implications because a more specific treatment can be selected [9]. Traditional culture- and microscope-based methods remain the gold standard, but the vitreous tap volume is limited, and traditional culture-based methods need a relatively large sample volume to grow bacteria [10]. Most pathogens that were cultured from biopsies are Gram-positive, such as Staphylococcus aureus, Staphylococcus epidermis and Streptococcus viridans, but some Gram-negative isolates such as Pseudomonas aeruginosa have also been grown from biopsies [10]. Using these classical methods, if successful, it takes several days to determine the infectious agent, but an additional complication is that a large proportion of the micro-organisms cannot be cultured [911]. In earlier studies when culture-based methods for pathogen identification were used, often only a single pathogen was identified, and these results seemed to imply that infectious endophthalmitis is caused by a single bacterium or fungus. Using PCR-based methods for pathogen identification, only pathogens that will be looked for can be identified, so the general idea of a single pathogen being responsible did not change [1214]. More recently, detection methods based on next-generation sequencing (NGS) have allowed for a more detailed and hypothesis-free analysis of the micro-organisms present in a sample. One of two general strategies is followed: targeted amplicon sequencing, for example of the 16S rRNA gene, or microbial whole-genome shotgun sequencing [1516]. Deep sequencing of fragments of the 16S rRNA gene was used with success to detect unculturable bacteria and to reduce the time of detection of pathogens [15]. Pathogens were detected in several samples from which it had not been possible to culture bacteria. A disadvantage is that the detection of a specific 16 rRNA only shows if a certain genus or species is present, but it does not reveal which resistance genes are present in the microbial population. To tackle this problem, shotgun metagenomic approaches are the best way to predict to which antibiotics a certain population is resistant and which treatment to choose [1720]. To gain a comprehensive understanding of the different types, mechanisms, transmission and evolution of antibiotic resistance one needs to understand the contribution of the whole microbiota. While our study was in progress, Zhu et al. [21] published a metagenomic NGS study to detect pathogens in endophthalmitis patients. They detected thousands of species of micro-organisms, making it difficult to identify the causal agent, but despite these results they claimed to have identified one or few causal pathogens per sample. It was concluded that NGS was more efficient than culture-based methods to identify pathogens, but they also noted that the NGS results were not always concordant with culture-based approaches [21].
Here, we present a metagenomic DNA shotgun sequencing study of biopsies obtained from Mexican endophthalmitis patients to describe the micro-organisms present and to identify the causal agents. After the sample was obtained patients were treated with a combination of ceftazidime/vancomycin or with MXF. Total DNA was purified from biopsies and sequenced. The resulting sequencing dataset was analysed by assembly of the short reads into larger contiguous DNA fragments and by mapping the sequencing reads to reference sequences. This method allowed the determination of the microbiota composition and was used to detect the antibiotic resistance genes present.
Methods
Clinical study and methods
This prospective case series study was approved by the internal review board of the Fundación Hospital Nuestra Señora de la Luz IAP, Mexico City. It was conducted in accordance with the Declaration of Helsinki and the purpose of the Good Clinical Practice Guidelines. All patients were informed of the experimental nature of the intervention, including a detailed description of the ‘Gold standard therapy’ with vancomycin and ceftazidime, possible complications of the disease, and the pros and cons of the experimental therapy. All patients signed an informed consent form before participating in the study.
We included male and female patients over 18 years of age, regardless of the cause [post-surgery, corneal ulcer, intravitreal injection (IVI), open globe trauma], with a clinical diagnosis of acute endophthalmitis. We excluded patients with no light perception visual acuity or with tissue lysis that required evisceration. Demographic data, laterality, type of surgery and days of symptom presentation were recorded.
After enrolment, all patients underwent a full ophthalmological examination which included an assessment of initial best-corrected visual acuity (BCVA), measured in logMAR units, slit lamp examination [conjunctival hyperemia, corneal epithelial damage, intraocular pressure, lens status, anterior chamber cells, anterior chamber haze and vitreous haze (using SUN Grading scheme for each one) [22]. If the anterior chamber or vitreous were not clear for clinical examination, an A/B scan ultrasound examination was realized. Pain as a variable was measured with an analog visual scale score.
In the operating room, vitreous tap was obtained, and 0.1 ml MXF was injected via pars plana into the vitreous under topical anaesthesia with draping in a routine sterile fashion. Samples were immediately refrigerated at 4 °C, then mixed with glycerol to a final concentration of 20 % (w/v) and stored in a deep freezer.
Patients underwent IVI of 0.1 ml MXF every 48 h if required until the ophthalmological examination showed signs of improvement (cornea haze reduction, anterior chamber inflammation decreased). Pars plana vitrectomy 25G with the Stellaris PC Platform (Bausch and Lomb) was then realized under retrobulbar anaesthesia. All patients were draped in a sterile fashion. The eyelid skin was disinfected with 10 % povidone iodine (PI) and the conjunctiva was disinfected with 4 % PI. Silicon oil was used as vitreous tamponade at the end of the surgery. Sclerotomies were sutured with 7–0 polyglactin (Vicryl; Ethicon). Moxifloxacin drops and prednisolone acetate were used after the surgery.
Patients were followed daily after IVI with MXF, variables of clinical examination were compared day 1 to the day previous to the vitrectomy in each case and final BCVA was determined 1 month after the surgery. Doses of MXF, time to the vitrectomy and follow-up time were recorded in each file.
Preparation of MXF solution for IVI
In this study we used commercial sealed MXF solution (Vigamoxi 0.5 %; Alcon; 500 µg/0.1 ml) for IVI. This formula was reported to be preservative-free and has a low toxic concentration for intraocular use in previous in vitro reports and case reports. A 0.1 ml aliquot of MXF (500 µg/0.1 ml) in the average vitreous volume of 4 ml corresponds to an empiric concentration of 125 µg ml^−1^. It has a half-life time of 1.72 h [78]. The MIC90 of fluoroquinolones in one case report was low, ≤1 µg ml^−1^ for ciprofloxacin and ≤2 µg ml^−1^ for levofloxacin, which might explain the clinical resolution of infections despite the short half-life time of this class of antibiotic [2324].
Samples and DNA extraction
Genomic DNA was isolated from 100 µl of vitreous or aqueous fluid using the Qiagen blood and tissue DNA extraction kit according to the manual. When less than 100 µl sample volume was available, the total volume was used for DNA extraction. The DNA was eluted in the kit elution buffer and stored at −20 °C. DNA concentration was determined using a NanoDrop ND-1 000 spectrophotometer (Thermo Scientific).
Sequencing, metagenome assembly and taxonomic classification
DNA was sequenced at INMEGEN (Nacional Institute of Genomic Medicine, Mexico City; samples 2, 3, 6, 7, 8) or at Omega Bioservices (Norcross, GA, USA; samples 15, 16, 17, 19). Paired-end sequencing was performed on MiSeq (250 bp ×2) or HiseqX10 (150 bp ×2) platforms. Quality analysis of the Illumina sequencing files was performed with FastQC (RRID:SCR_014583, https://www.bioinformatics.babraham.ac.uk/projects/fastqc/) software version 0.11.9; low-quality bases and adapters were removed with Trim Galore (RRID:SCR_011847, https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/) software version 0.6.6, and the first 15 bases of the reads of all fast files were removed because or erroneous base sequence content. The human genome sequence version 38 was obtained from the refseq ftp site of the NCBI [25], this genome was indexed with Bowtie2build and the Illumina reads were aligned with Bowtie2 [26] software v2.3.4.1. (RRID:SCR_016368, http://bowtie-bio.sourceforge.net/bowtie2/index.shtml). The Illumina reads that aligned concordantly to the human genome were excluded from the fastq files to obtain the human filtered fastq files.
De novo genome assembly was performed for each metagenome sample with SPAdes [27] software version 3.14.1 with metagenome option and kmer values of 21, 33, 55, 77, 99, 111 and 127, using the human filtered fastq files. The assembled contigs were used for gene prediction and annotation with Prokka [28] version 1.12 with the metagenome option. Taxonomic classification was performed with Kraken2 [25] software version 2.0.8-beta using the assembled contigs with the mini-kraken2_v1_8 Gb database with the -use-mpa-style option. The Kraken2 results were analysed and visualized with Pavian [29] software version 1.2.0. The protein sequences of all metagenome samples were blasted [30] against the last version of the full dataset Virulence Factor Database (VFDB) [31] and the protein homologue model type of the Comprehensive Antibiotic Resistance Database (CARD) [32]. Blast hits with these two databases were considered significant if the protein query and hit have 80 % alignment coverage and 40 % sequence identity.
Results and discussion
Clinical information
From June 2018 to August 2019, 19 patients with endophthalmitis were evaluated. Their mean age was 63.6±16.2 years; four patients (21.0 %) were female and 15 patients (78.9 %) were male. In nine patients (47.3 %) the left eye was affected. Two patients had an open wound, and 17 cases were associated with ophthalmological procedures. In nine eyes cataract surgery was the potential cause of infection, in four eyes IVI, in two eyes corneal ulcer, in one eye penetrating keratoplasty and in one eye vitrectomy was the cause. The mean time from surgery to the start of symptoms was 7.2±8.4 days. Four patients were treated with a combination of ceftazidime and vancomycin and the other 15 with MXF. The mean initial visual acuity was 2.48 logMAR (hands motion or 20/4 000), and at the end, the mean VA was 1.8 logMAR (count fingers or 20/1 000). However, we registered patients with a 20/20 visual acuity at the end. Only one patient showed a worsened clinical status, but for the others, intravitreal antibiotics improved corneal oedema enough to realize an early pars plana vitrectomy. Demographic and clinical information of the included cases is given in Table 1. The total number of sequenced samples was low, but as this post-surgical complication is not very frequent, samples are very difficult to obtain and frequently are of low volume. In addition, to our knowledge this is only the second study of this type of sample and the first with samples from Latin America.
Effect of moxifloxacin treatment
After the full ophthalmological exam, a vitreous tap sample was obtained, and antibiotic treatment was initiated. Three of the first patients to present as candidates for the protocol accepted vitreous sample collection but not the application of MXF; the remaining patients accepted both interventions. As seen in Table 1, one of the patients treated with the dual antibiotic had to undergo evisceration due to poor progression. Within the MXF group, all patients showed clinical improvement that allowed advancement to a pars plana vitrectomy, enabling a complementary surgical approach for resolution. We conclude that the use of MXF, compared to the use of dual antibiotics, was not inferior, did not worsen the condition, and allowed for surgical intervention to improve the structural and functional conditions of the patient. Therefore, MXF can be considered an additional alternative for treating this highly disabling pathology, aggressive to ocular tissue, and requiring diverse diagnostic and management alternatives due to its presentation with multiple pathological agents, as observed in this study. A detailed description of the cases is beyond the scope of this publication and will be published in a separate article.
DNA extraction and sequencing
A fundamental challenge is that only small volumes of intraocular fluid can be safely obtained, and, in our study, this sometimes was not enough to extract sufficient DNA for successful library construction and sequencing. DNA extraction for metagenomic shotgun sequencing can be difficult in samples with lower microbial biomass and high human DNA content [33]. Some samples contain human tissue fragments, whereas others do not. Some samples may contain high numbers of bacterial cells, whereas others do not. This means that sample characteristics can differ from sample to sample, which could explain the different amounts of DNA that could be extracted from the biopsies. In one case, no DNA could be detected after extraction of the sample and from other samples only minimum amounts not sufficient for library construction were obtained. In the nine samples that could be sequenced we obtained between 2.2 and 191 million sequencing reads (Table 2). Sequencing was done at two different sequencing facilities, which is the main reason for the difference in sequencing depth reported. The first five samples were sequenced at a Mexican non-commercial institution, and the final four samples were sent abroad. As expected, a large fraction of the reads were assigned to human DNA. Depending on the sample between 16.6 and 62.4 % of the sequencing reads were therefore excluded from further analysis. Previous studies had observed between 81 and 99 % human reads [34]. From these filtered reads between 12 000 and 1.8 million contigs could be assembled, but only between 2.4 and 38 % of the assembled contigs could be classified properly by Kraken2 (Table 2).
Metagenome sequencing shows that a complex mixture of bacteria and virus can be detected in the biopsies
After assembling the sequencing reads into contigs, these were assigned to taxonomic classes. By abundance, six major taxonomic classes were detected in each of the biopsies: Betaproteobacteria, Gammaproteobacteria, Alphaproteobacteria, Bacilli, Actinobacteria and Mollicutes. In addition, smaller numbers of reads and contigs could be assigned to other taxonomic classes. The relative amounts of the taxonomic classes varied substantially (Fig. 1) but already at this point in our analysis we noted that the sample compositions were complex and by no means were monocultures dominated by a single bacterium. In previous studies mainly Gram-positive bacteria were isolated from endophthalmitis patients, but in the samples from Mexican patients that we analysed, most bacteria detected were proteobacteria (Fig. 1). We then tried to assign the contigs at the species level and it was confirmed that a large diversity of bacteria was present in the biopsies (Table 3 and Fig. 2). Many of the bacteria detected in this study are known as pathogens or opportunistic pathogens and are identical to typical bacteria identified by earlier studies using culture-dependent approaches or PCR-based approaches as causal agents of the infection. Staphylococcus aureus, Staphylococcus epidermidis, Streptococcus pneumoniae, Pandoraea norimbergensis, Ralstonia pickettii, Pasteurella multocida, Neisseria gonorrhoe, Burkholderia pseudomallei, Enterobacter cloacae, Klebsiella pneumoniae, Paracoccus mutanolyticus, Streptomyces sp. ICC1, Bradyrhizobium sp. SK17, Delftia tsuruhatensis, Sphingomonas sp. FARSPH, Stenotrophomonas maltophilia, Herbaspirillum seropedicae, Variovorax paradoxus and Pseudomonas aeruginosa were among the bacterial species most frequently detected across the samples (Fig. 2 and Table 3). Additionally, some of the bacterial genera we detected are known pathogens but have been also described to be part of the core genera at the ocular surface such as Pseudomonas, Staphylococcus, Streptococcus and Sphingomonas. Other genera that we detected, such as Ralstonia and Delftia, are known to contain pathogenic strains causing infection in other tissues [35]. We were surprised to detect Bradyrhizobium sp. SK17 in many of our samples, but this genus, best known for members that are nodule-forming bacteria on the roots of legume plants [36], has also been described as a typical component of the core microbiome of the ocular surface [35]. In our study, in eight out of nine samples more than half of the detected bacteria were proteobacteria, which is high compared to previous studies. In European and North American reports, in 4–6 % of the cases Gram-negative bacteria were the cause of endophthalmitis, but in Asian, Chinese and Indian studies this number increased to over 20 % [3743]. The normal microbiota of the healthy ocular surface and the pathogens of an infected eye vary widely, depending on the age and immune condition of the patient, geographical location and ambient climate [34].
Relative abundance of sequencing reads assigned to taxonomic classes. Samples from left to right: META2, META3, META6, META7, META8, C15, C16, C17 and C19.
Heatmaps showing the 25 most abundant micro-organisms and virus per sample. (a) Samples META2, 3, 6, 7 and 8. (b) Samples C15, 16, 17 and 19. The top bar in both panels represents the mapping of the numerical values to a colour scale.
As the torque teno virus was identified in a few patients with culture-negative endophthalmitis [44], we were also interested in the viral reads we could detect. We looked specifically for reads that could be mapped to virus. In some samples we detected BeAn 58 058 virus, which belongs to the Poxviridiae. It was first isolated in 1963 from the blood of a rodent of the genus Oryzomis in a tropical rain forest, in the region of Belém-do-Pará, Brazil [45]. Interestingly, this virus was also detected in the lung microbiome from patients with chronic obstructive pulmonary disease [46]. Other viral reads were detected in much lower numbers or were only present in very few samples.
We also looked specifically for reads and contigs that could be mapped to fungi. Malassezia restricta was detected in five samples (C15=7 contigs, C16=13 contigs, C17=4 contigs, C19=6 contigs, META6=2 contigs). In a few samples single contigs of Fusarium graminearum (19), Pyricularia pennisetigena (8 and 16), Neurospora crassa (17) and Sugiyamaella lignohabitans (16) were detected. Deshmukh et al. had also observed fungi from the genus Malassezia in several samples of endophthalmitis patients [47] and Fusarium sp. had also been described in previous studies [48].
The results from the assignment of the sequencing contigs to bacterial species were used to prepare two different heat maps to further characterize and compare the different samples. The division was made because it allowed an easier interpretation despite the large differences in sequencing reads between the two groups of samples (Fig. 2). Sequencing depth has a major impact on the results of metagenomic shotgun sequencing [49]. At decreased sequencing depth, the number of undetected microbial species increases together with unclassified and misclassified clades. The large bacterial diversity that we and others [10] detected in the biopsies complicates the identification of the causal agent for the infection. It is important to emphasize that our data showed dramatically different prevalence and much greater diversity at the genus level than typically revealed by classical culture-based methods (Table 3 and Fig. 2a and b). At least for our small number of samples, it is not a single bacterial species that is dominant, but we observed complex microbiomes.
In our shotgun sequencing study, many bacterial species were identified in all nine samples that could be sequenced. This is an observation usually completely missed by culture-based methods [47]. Often, the bacteria involved appear to originate from the patients’ own microbiota but may also be introduced through contaminated solutions or instruments used during ophthalmological procedures [50]. Pathogenic bacteria identified in other studies as causal agents were identified but were often not among the most abundant micro-organisms. A complication of this observation is that it is unclear which bacteria are mainly responsible for the infection and the tissue damage. Other bacterial species normally not known to be involved in pathogenesis might, however, influence the process of infection because they might harbour antibiotic resistance genes that could be transferred horizontally to the pathogens.
The heterogeneity of the biopsies did not allow successful extraction of DNA in all cases. In some cases, enough DNA could be extracted, but in others not. Clearly, this is a problem when suggesting metagenomic shotgun sequencing for diagnosis. Also, metagenomic shotgun sequencing is currently still too time-consuming and methodologically too complex to become a routine method. We think that recent methodological developments such as Nanopore sequencing may have a future application for diagnostic purposes that can improve the quality of care of patients with infectious endophthalmitis. This type of sequencing allows results to be obtained in a shorter timeframe. Recently, an article was published describing a turnover time from 7 to 9 h from sample collection until an informed decision for sepsis treatment using Nanopore sequencing [51].
Vancomycin resistance genes and a large diversity of efflux pumps are detected by metagenome sequencing
Antibiotic resistance is spreading worldwide, and this situation is especially worrying in clinical settings. For the efficient treatment of patients with endophthalmitis, the identification of the causal agent(s) is important in choosing the correct treatment. Most common antibiotics target conserved mechanisms of bacterial metabolism such as cell wall synthesis (beta-lactams, vancomycin), protein synthesis (tetracycline and neomycin), DNA replication (fluoroquinolones) and single carbon metabolism such as folate synthesis (sulfonamides) [19]. Bacteria become resistant by four major mechanisms: enzymatic degradation or modification of the antibiotic, alteration of the antibiotic target, active efflux of the antibiotic and reduced permeability of the bacterial cell surface. The resistance mechanisms can be specific or they can be more general; for example, a beta-lactamase will only act on beta-lactam antibiotics, whereas an efflux pump can export different antibiotics out of the cell or a cell wall modification affecting the cell wall permeability can make a cell less permeable for several antibiotics [19]. Metagenomic deep sequencing allows us to identify specific antibiotic resistance genes present in a microbial population [185253]. Frequently, these resistance genes are located on mobile genetic elements, and this allows rapid spread of the resistance in a bacterial population [54], but which can complicate the assignment of a resistance gene to a specific species.
Using the contigs assembled for taxonomic determination, we searched the CARD database for the presence of known antibiotic resistance genes. When comparing the number of antibiotic resistance genes that could be detected, we observed some differences. In the five samples of the first group with a lower sequencing depth (samples 2, 3, 6, 7, 8) between zero and nine resistance genes were detected per sample. In the four samples of the second group with a higher sequencing depth (samples 15, 16, 17, 19), between 14 and 115 resistance genes were detected (Fig. 3). A lower sequencing depth should lead to a decrease in the number of detected resistance genes, and low-abundance resistance genes might not be detected. This might explain why fewer resistance genes were detected in the biopsy samples sequenced with a lower sequencing depth [49].
Heatmap of antibiotic resistance genes present in the samples.
We detected genes conferring resistance against trimethoprim, different aminoglycoside resistance genes (APH, AAC), vancomycin resistance gene clusters, members of different beta-lactamase families (OXA, NmcA, AIM, LRA, SPG), sulfonamide resistance genes, genes encoding enzymes changing cell wall permeability (Fig. 3) and, worryingly, an impressive amount of predicted antibiotic efflux pumps, belonging to the RND, MFS, SMR and ABC transporter families [5557]. Some OXA beta-lactamase family members confer resistance predominantly to ceftazidime and additionally we detected genes conferring resistance to vancomycin, which is used together with ceftazidime in the treatment of endophthalmitis. Given the importance of the putative vancomycin and ceftazidime resistance genes for the treatment of endophthalmitis patients, we investigated them more closely. In three of the samples, we identified putative vancomycin resistance genes with about 40 % identity at the amino acid level and a coverage of 100 % and in one sample we detected vancomycin resistance genes with an identity of more than 90 % at the amino acid level and a coverage of 100 %. Genes encoding putative OXA beta-lactamases were detected in two of the samples, in both cases with identities of more than 90 % at the amino acid level and a coverage of 100 % to known resistance genes.
It is much more difficult to predict the specificity of the efflux pumps. Efflux pumps frequently have a broad substrate specificity and usually it is not possible to predict what the substrates are, although recently important advances have been made in this area [58]. In addition, it has been observed that mutations in the corresponding genes can modify the substrate specificities of the efflux pumps [505960]. Living organisms express a large variety of efflux pumps with broad specificity to deal with a large diversity of xenobiotics in the environment. These transporters, often with overlapping substrate specificities [61], recognize a wide range of substrates that may differ in structure, size or electrical charge and actively remove them from cells, thereby providing an essential survival strategy for the organism [62].
Conclusions
In the present study, we sequenced total DNA extracted from biopsies obtained from eyes of acute endophthalmitis patients. In none of the samples was a single bacterial species dominant, but in all cases, we detected complex metagenomes, an observation which complicates the prediction of the agents causing the infection and the degradation of the tissue. Within this complexity we observed bacterial species that have been described as part of the skin microbiome and part of the ocular surface microbiome.
Using shotgun metagenomics, several resistance genes against a wide range of antibiotics were detected. Among these were resistance genes encoding OXA beta-lactamases which might confer resistance to ceftazidime and genes conferring resistance to vancomycin, two of the antibiotics currently used against endophthalmitis. It is also worrying that many genes encoding efflux pumps were identified.
Clinical metagenomics as a tool in ophthalmology has a large potential to reveal the variety of bacteria, viruses and fungi that can be present in endophthalmitis, but the analysis is still too time-consuming to be used as a routine tool in the clinic. It is probably of value to determine the metagenomes of thousands of patients worldwide to get a complete image of the organisms that can be present in endophthalmitis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bowen RC Zhou AX Bondalapati S Lawyer TW Snow KB et al Comparative analysis of the safety and efficacy of intracameral cefuroxime, moxifloxacin and vancomycin at the end of cataract surgery: a meta-analysis Br J Ophthalmol 20181021268127610.1136/bjophthalmol-2017-31105129326317 PMC 6041193 · doi ↗ · pubmed ↗
- 2Kernt M Kampik A Endophthalmitis: pathogenesis, clinical presentation, management, and perspectives Clin Ophthalmol 2010412113510.2147/opth.s 646120390032 PMC 2850824 · doi ↗ · pubmed ↗
- 3Nakashizuka H Shimada H Hattori T Tanaka K Kitagawa Y et al Intravitreal injection of 1.25% povidone iodine followed by vitrectomy using 0.025% povidone iodine irrigation for treating endophthalmitis Transl Vis Sci Technol 201982110.1167/tvst.8.1.21PMC 637511630783556 · doi ↗ · pubmed ↗
- 4Pathengay A Khera M Das T Sharma S Miller D et al Acute postoperative endophthalmitis following cataract surgery: a review Asia Pac J Ophthalmol 20121354210.1097/APO.0b 013e 31823 e 574b 26107016 · doi ↗ · pubmed ↗
- 5Das T Endophthalmitis management: stain-culture, empirical treatment, and beyond Asia Pac J Ophthalmol 202091310.1097/01.APO.0000617904.11979.ae PMC 700444531990737 · doi ↗ · pubmed ↗
- 6Grzybowski A Turczynowska M Kuhn F The treatment of postoperative endophthalmitis: should we still follow the endophthalmitis vitrectomy study more than two decades after its publication?Acta Ophthalmol 201896 e 651e 65410.1111/aos.1362329197165 · doi ↗ · pubmed ↗
- 7Gao H Pennesi ME Qiao X Iyer MN Wu SM et al Intravitreal moxifloxacin: retinal safety study with electroretinography and histopathology in animal models Invest Ophthalmol Vis Sci 2006471606161110.1167/iovs.05-070216565399 · doi ↗ · pubmed ↗
- 8Kernt M Neubauer AS Ulbig MW Kampik A Welge-Lüssen U In vitro safety of intravitreal moxifloxacin for endophthalmitis treatment J Cataract Refract Surg 20083448048810.1016/j.jcrs.2007.10.04618299076 · doi ↗ · pubmed ↗