Two novel members of Onygenales, Keratinophyton kautmanovae and K. keniense spp. nov. from soil
Roman Labuda, Vanessa Scheffenacker, Andreas Schüller, Broňa Voleková, Alena Kubátová, Hazal Kandemir, Winnie Cherotich Maritim, Josphat Matasyoh, Markus Gorfer, Christoph Schüller, Joseph Strauss

TL;DR
Two new fungi species, Keratinophyton kautmanovae and K. keniense, were discovered in soil samples from Africa and Europe.
Contribution
The discovery of two new Keratinophyton species from distinct geographical regions adds to the fungal diversity of the Onygenales order.
Findings
K. kautmanovae and K. keniense are phylogenetically distinct and supported by ITS and LSU rDNA sequences.
K. keniense is the first Keratinophyton species reported from Africa.
Both species exhibit unique morphological traits, such as conidial shapes and growth characteristics.
Abstract
Two new Keratinophyton species, K. kautmanovae sp. nov. and K. keniense sp. nov., isolated from soil samples originating from two different geographical and environmental locations (Africa and Europe) are described and illustrated. Phylogenetically informative sequences obtained from the internal transcribed spacer (ITS) region and the nuclear large subunit (LSU) rDNA, as well as their unique phenotype, fully support novelty of these two fungi for this genus. Based on ITS and LSU combined phylogeny, both taxa are resolved in a cluster with eight accepted species, including K. alvearium, K. chongqingense, K. hubeiense, K. durum, K. lemmensii, K. siglerae, K. submersum, and K. sichuanense. The new taxon, K. kautmanovae, is characterized by clavate, smooth to coarsely verrucose conidia, absence of arthroconidia, slow growth at 25 °C, and no growth at 30 °C, while K. keniense is…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
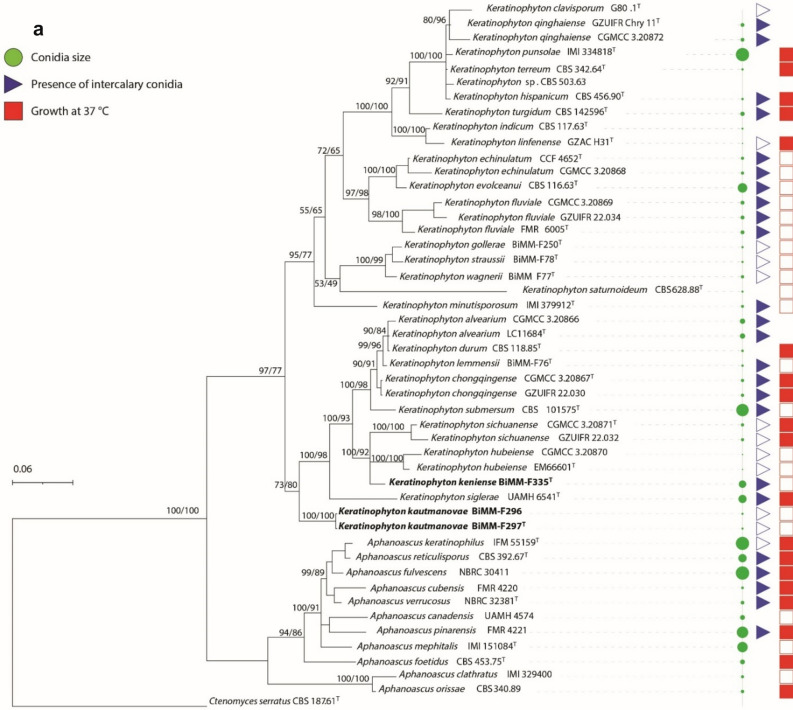 Figure 1
Figure 1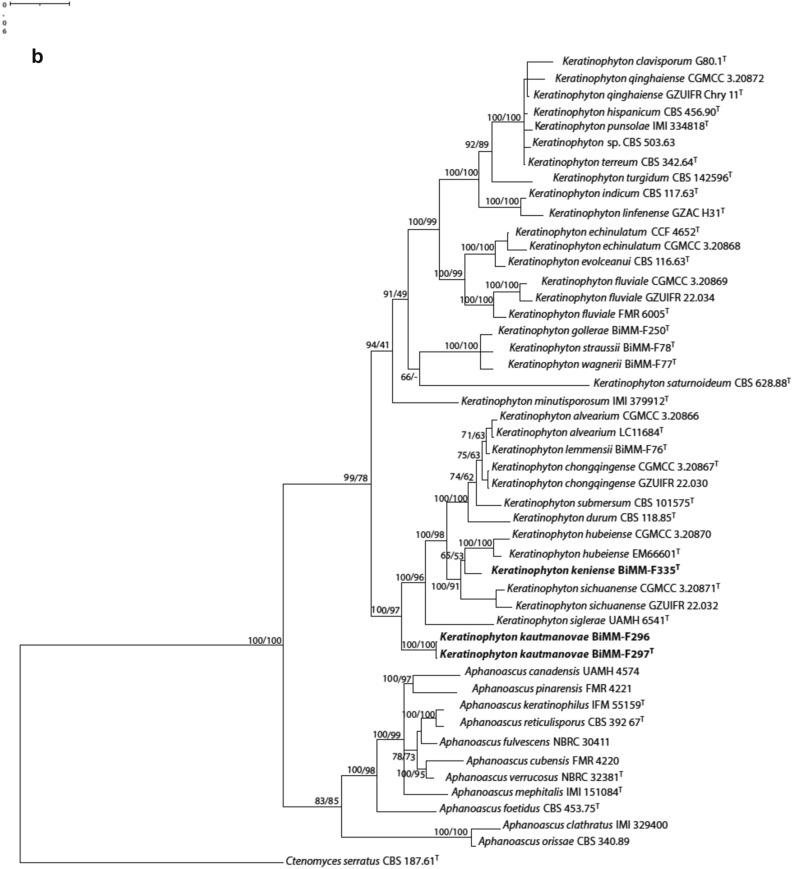 Figure 1
Figure 1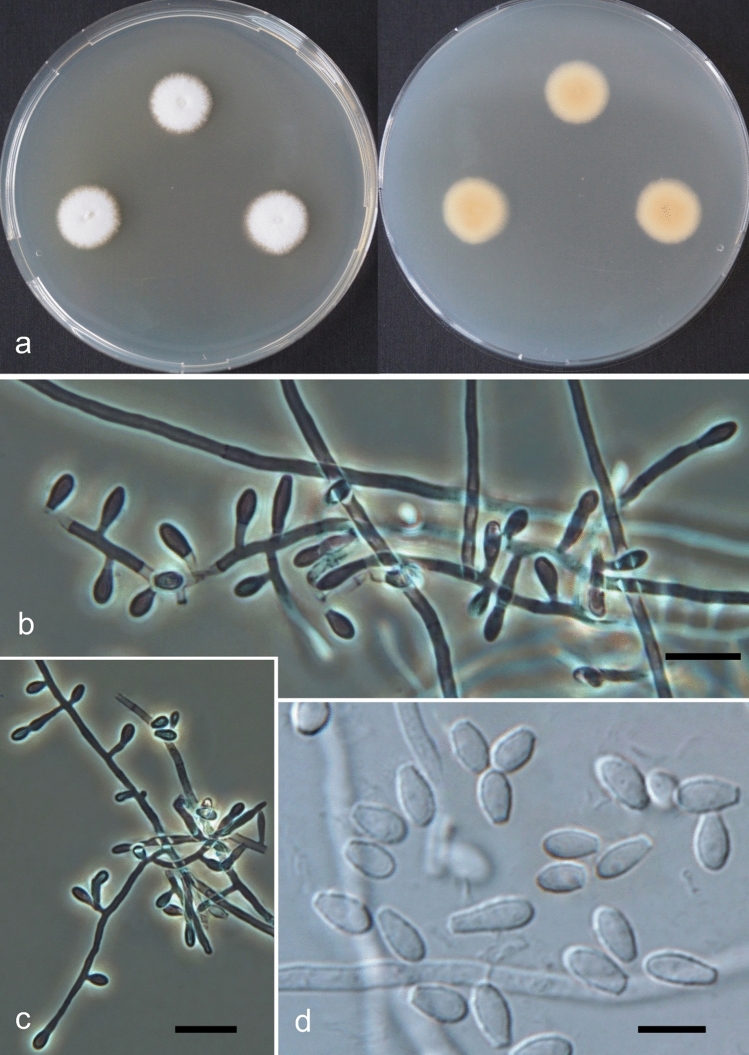 Figure 2
Figure 2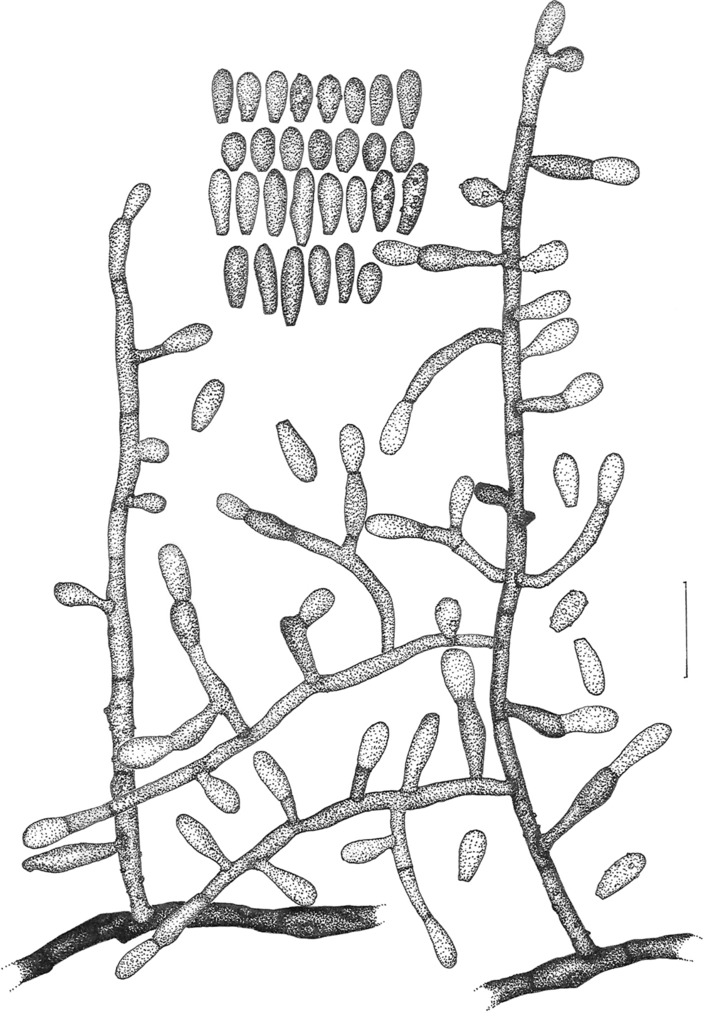 Figure 3
Figure 3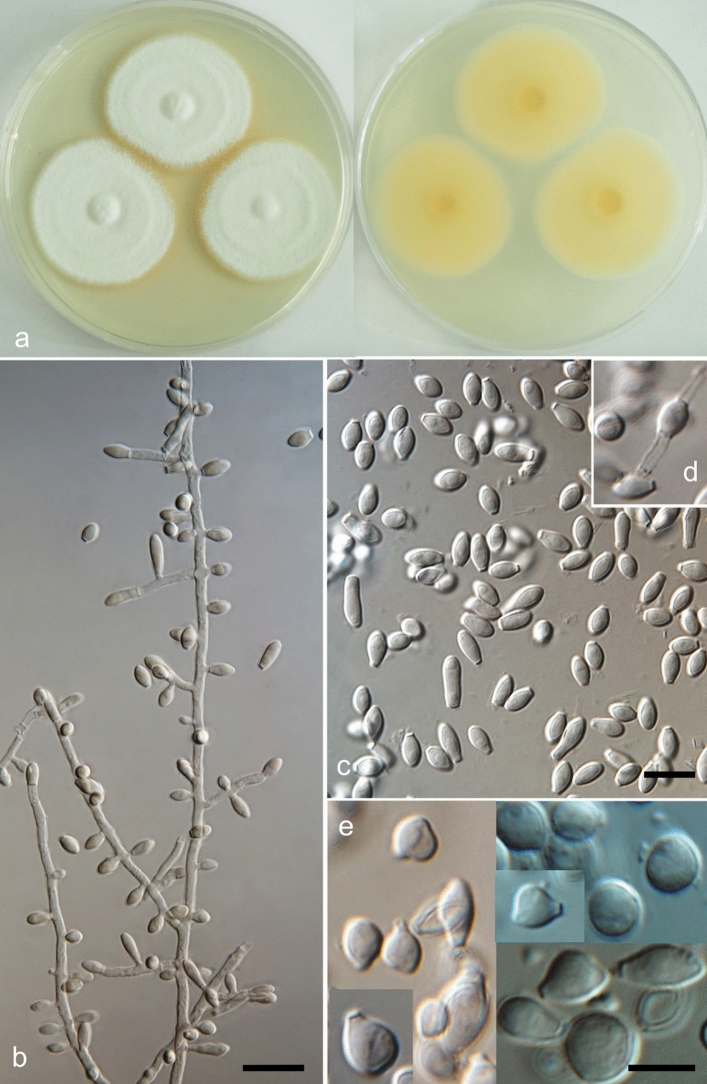 Figure 4
Figure 4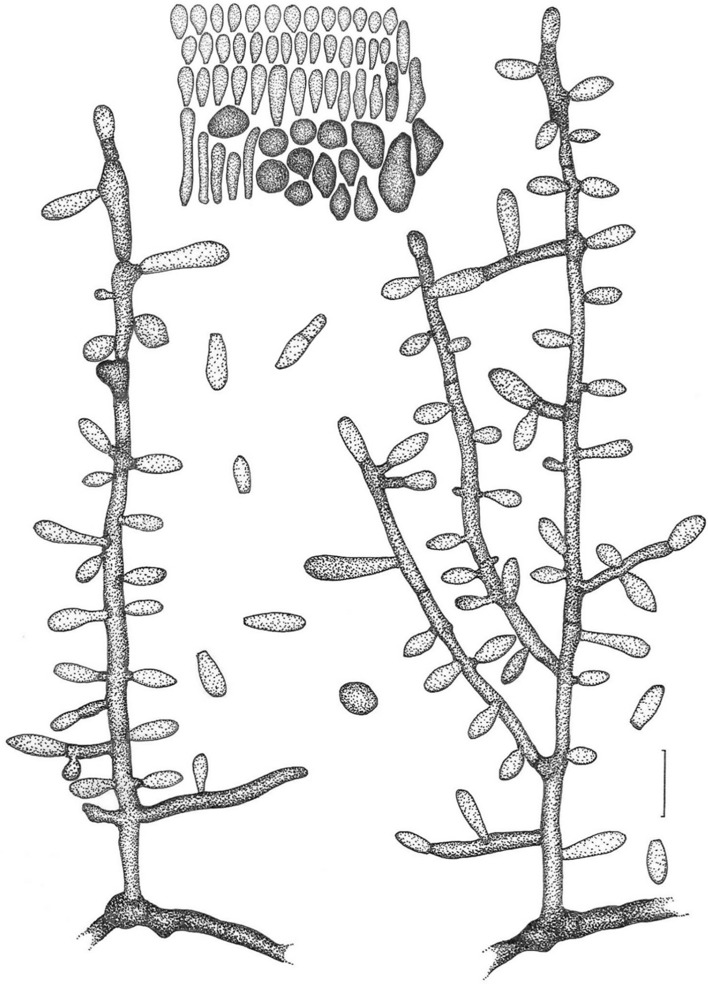 Figure 5
Figure 5- —Lower Austria Science and Education Fund (NfB/GFF)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Lichen and fungal ecology · Mycorrhizal Fungi and Plant Interactions
Introduction
The genus Keratinophyton was introduced in 1964 by Randhawa & Sandhu^1^ However, the nomenclatural history of this genus was confusing since untill recently all known Keratinophyton species were classified in Aphanoascus (Cooke) Apinis^2–4^. Members of Keratinophyton and Aphanoascus share cleistothecia with a pseudoparenchymatic peridium, and they can be found on keratinous substrata and dung^3,5,6^. Sutton et al. (2013) re-established the genus Keratinophyton with K. terreum as the type species, based on ascospore morphology^5^. These two genera are distinguished by their ascospore morphology, where ascospores with a conspicuous equatorial rim and pitted wall are characteristic for Keratinophyton, while Aphanoascus species produce reticulate ascospores without a rim^5^, and by result of phylogenetic analysis using ITS (the internal transcribed spacer) and LSU [the D1/D2 domains of the large-subunit (28S) rDNA gene] loci^5–7^. The monophyletic Keratinophyton clade currently encompasses six species, namely K. durum, K. hispanicum, K. multiporum, K. punsolae, K. saturnoideum, and K. terreum, known to form in vitro ascosporic state (sexual morph) as well as 19 taxa known only as asexual morphs, characterized mainly by a chrysosporium-like conidiogenesis^6–8^. Actually, the genus Keratinophyton includes several species described in Chrysosporium^9–14^. These species however, have been regarded as having a doubtful identity^6^.
The genus Keratinophyton is represented by a large group of keratinolytic soil-borne fungi rather common in areas with high animal activity resulting in transfer of keratinous material to the soil. Ecology and distribution of the genus has been reviewed in a previous study, stating that soil and soil-like substrata are primary habitats for this fascinating group of onygenalean fungi^7^. Currently, this genus comprises a total of 27 recognized and accepted taxa^6,8^.
We performed a mycological survey with emphasis on keratinophilic fungi in environmental samples taken from Republic of Kenya, Africa and Slovak Republic, Europe. Herein, we present description of two novel morphologically and phylogenetically distant species within the genus Keratinophyton being characterized by their unique chrysosporium-like morphs.
Material and methods
Sample collection and isolation of the fungi
The soil sample was collected in front of the Budúcnosť adit at the abandoned antimony (Sb) deposit Pezinok—Kolársky Vrch, 48°19′03.3" N 17°14′20.9" E, locality Rudné Mines, Pezinok (Slovak Republic) in December 2021. Sampling site was situated approximately one meter from neutral mine drainage contaminated by potentially toxic elements.
The second soil sample was collected from Egerton University campus, 0°22′09.3" S 35°55′24.2" E, Njoro (Republic of Kenya) in February 2022.
The samples from the surface layer (up to 20 cm deep) were dried and stored in at 5‒8 °C. Isolation of the keratinophilic fungi was performed as previously described^15^, with a modification according to^7^. A sample was divided into five subsamples. The subsamples (5 g each) were poured into Petri dishes (50 mm in diameter) and mixed with 0.5 g Vermiculite, then soaked with 3‒4 mL (depending on moiety of the sample) antibiotic solution containing 0.5 g/L cycloheximide and 0.1 g/L chloramphenicol. Sterile defatted horse hair fragments (10 pieces of ca 2.0 cm per plate) were used as baits. The Petri dishes were then incubated at laboratory temperature (23‒25 ± 1 °C), in dark, for a period of 2–3 months and remoistened with sterile deionized water when necessary^7,15^. The Petri dishes were checked weekly for the presence of fungi, and colonies were transferred on Sabouraud 4% dextrose agar (SDA; Merck, Darmstadt, Germany) supplemented with 0.5 g cycloheximide and 0.05 g chloramphenicol. Pure cultures were then transferred onto potato dextrose agar (PDA; Van Waters and Rogers (VWR) International, Leuven, Belgium)^7^.
Morphological analysis
The preliminary identification of the resulting keratinophilic fungi was carried out based on their phenotypic characteristics^9–11^.
For phenotypic determination, the strains were transferred by three-point inoculation onto PDA, Malt Extract Agar (MEA; Merck, Darmstadt, Germany), and SDA, and incubated for 14 d in the dark at 25 °C. Christensen´s urea agar (Sigma-Aldrich, St Louis, MO, USA) was used for additional physiological and biochemical characteristics (25 °C, 14 days, in the dark)^7^.
Colony growth rate (mm), colony structure and characteristics such as production of exudates and pigments were noted after 14 days (on PDA, MEA, and SDA). However, the cultivation was extended up to 3 months to observe and record changes in pigmentation of the colonies as well as to determine the onset of sexual reproduction^7^. In order to determine the optimal and minimum/maximum temperatures for growth, PDA, MEA and SDA plates were incubated at 5 °C, 8 °C, 10 °C, 12 °C, 15 °C, 18 °C, 20 °C, 25 °C, 28‒32 °C, 35 °C, and 37 °C, and the growth rate was measured on the 14th day of cultivation. For comparative descriptions of the macroscopic and microscopic characteristics, PDA was used according to^7,11,16^.
For observation of microscopic traits 14‒18 days growth on PDA was used. Conidiophore and conidia formation were observed in situ under low magnification (50–100×). Details of conidiophores, conidia (aleurioconidia) and other microscopic structures, such as width of hyphae, were observed in Melzer´s reagent and lactic acid with cotton blue^7^. Photomicrographs were taken using phase and Nomarski contrast optics on an Olympus BX51 microscope with Olympus DP72 camera and QuickPHOTO Micro 3.0 software. Photographs of the colonies were taken with a Sony DSC-RX100.
Dried fungal specimens were deposited as holotypes in the collections of the Mycological Department, National Museum in Prague, Czech Republic (PRM); ex-type cultures were deposited in the Bioactive Microbial Metabolites (BiMM) Fungal Collection, UFT- Tulln in Austria and in the Culture Collection of Fungi in Prague (CCF)^7^.
Keratinolytic activity
A hair perforation test was performed following de Hoog et al. (2020) using 25 mL water containing 2–3 drops 10% yeast extract (YEW)^17^. The hairs were examined microscopically 14 and 21 days after inoculation at 25 °C in the dark. At the end of the incubation period, a few pieces of hair were taken out from the testing medium. The overgrowing fungus was deactivated with 70% ethanol and then removed from the hair surface mechanically in a stream of a tap water^7^. The degree of hair digestion-degradation (keratinolytic activity) was assessed in the light microscope under 100× and 400× magnification. Water was used as mounting fluid for the observation and microphotography of the hair samples. Intensity of degradation^18^ of hair was estimated on a scale of 0 to 4: 0 = no degradation; 0–1 = light degradation on the cuticle; 1 = moderate degradation on the cuticle and/or rare formation of boring hyphae; 2 = degradation of cuticle and cortex, with about 20% degradation; 3 = degradation of cuticle and cortex, with about 50% degradation; 4 = degradation of cuticle and cortex, with about 80% degradation.
DNA extraction, PCR amplification and sequencing
DNA was extracted using a standard cetyltrimethyl ammonium bromide (CTAB) procedure, as described previously^7,19^. The internal transcribed spacer (ITS) region was amplified with primers ITS1-F^20^ and ITS4^21^ using Taq-polymerase (GoTaq G2 Green Master Mix from Promega). The D1/D2 domains of the large-subunit (28S) rDNA gene (LSU) were amplified and sequenced using the primer pair ITS1/TW14^21,22^. All reactions were performed in an Eppendorf Gradient MasterCycler (Eppendorf, Hamburg, Germany). Conditions for amplification of ITS and LSU domains: 95 °C for 5 min; 35 cycles of 95 °C for 30 s, 54 °C for 30 s, 72 °C for 90 s, and finally 5 min at 72 °C^7^. The PCR products were sequenced with the same primers used for the PCR amplifications (LGC, Berlin, Germany). All sequences obtained in this study were deposited in GenBank nucleotide database (Table 1).Table 1. List of the strains included in the study.Species nameStrainSourceGenBank accession numbersITSLSUA.* canadensisUAMH 4574Carnivore dung, CanadaAJ439435–A. clathratusIMI 329400Arable soil, SpainAJ439436–A. cubensisFMR 4220Soil of tobacco field, CubaAJ439432–A. foetidusCBS 453.75^T^Myomys daltoni coat, NigeriaKT155907KT155252A. fulvescensNBRC 30411Soil of rice paddy field, JapanJN943432JN941547A. keratinophilusIFM 55159^T^Pasture land soil, Papua New GuineaNR165936NG064030A*.* mephitalisIMI 151084^T^Dung of wolf, CanadaAJ439439AY176725A. orissaeCBS 340.89Soil in animal husbandry, KuwaitAJ390393–A. pinarensisFMR 4221Forest soil, CubaAJ439433–A. reticulisporusCBS 392.67^T^Soil, New ZealandMH859002MH870704A. verrucosusNBRC 32381^T^Arable soil, SpainNR131309NG057011K. alveariumLC 11684^T^Hive-stored pollen, ChinaMF939598MF939580CGMCC 3.20866Soil, ChinaOM952124OM952112K. clavisporumG80.1^T^Plant root soil, ChinaKY026601–K. chongqingenseCGMCC3.20867^T^Green belt soil, ChinaOM952125OM952113GZUIFR 22.030OM952126OM952114K. durumCBS 118.85^T^Soil, NepalMH861856AB075345K. echinulatumCCF 4652^T^Sole of the foot, CzechiaLT548276LT548276K. evolceanuiCBS 116.63TSoil, IndiaAJ005368MH869834K. fluvialeFMR 6005^T^River sediments, SpainAJ005367MT875000CGMCC 3.20869Soil, ChinaOM952132OM952120GZUIFR 22.034Soil, ChinaOM952133OM952121K. golleraeBiMM F250^T^Forest soil, SlovakiaMN633084MT874997K. hispanicumCBS 456.90^T^Beach soil, SpainKT155910MT875003K. hubeienseEM66601^T^Soil under the chicken feather, ChinaKJ849227–CGMCC 3.20870Soil, ChinaOM952128OM952116K. indicumCBS 117.63^T^Soil, IndiaNR_145203KT155044K. kautmanovaeBiMM F297^T^**BiMM 2296Soil nearby a Sb stream Slovakia**Soil nearby a Sb stream Slovakia***PP062810PP062811PP062954PP062955K. kenienseBiMM F335**^T^Soil, Republic of Kenya**PP062809PP062953K. lemmensiiBiMM F76^T^Compost soil, AustriaMN633082MT874998K.* linfenenseGZAC H31^T^Rhizosphere soil, ChinaNR158289–K. minutisporosumIMI 379912^T^River sediments, SpainKT155616MT875001K. punsolaeIMI 334818^T^Arable soil, SpainAJ439440–K. qinghaienseGZUIFR Chry 11^T^Farmland soil, ChinaJX868607–CGMCC 3.20872Soil, ChinaOM952134OM952122K. saturnoideumCBS 628.88^T^Arable soil, SpainNR077135AB075347K. sichuanenseCGMCC 3.20871^T^Green belt soil, ChinaOM952130OM952118GZUIFR 22.032Green belt soil, ChinaOM952129OM952117K. sigleraeUAMH 6541^T^Garden soil, SpainAJ131684MT875002Keratinophyton sp.CBS 503.63Forest soil, IndiaKT155929KT155274K. straussiiBiMM F78^T^Garden soil, ItalyMN633081MT874996K. submersumCBS 101575^T^River sediments, SpainNR157445NG064180K*.* terreumCBS 342.64^T^Lawn soil, IndiaKT155876KC989709K. turgidumCBS 142596^T^Barber shop soil, IndiaKY290503KY962732K. wagneri*BiMM F77^T^Forest soil, SlovakiaMN633083MT874999Ct. serratusCBS 187.61^T^Soil, AustraliaNR144890AY176733Data in bold generated in the present study.BiMM bioactive microbial metabolites unit, UFT-Tulln, Austria, CBS (Westerdijk Fungal Biodiversity Institute), Utrecht, The Netherlands, CCF culture collection of fungi, Charles University, Prague, Czech Republic, EM, and GZUIFR = CGMCC = LC strains The Institute of Fungus Resource, Guizhou University, China, FMR Facultat de Medicina in Ciències de la Salut, Reus, Spain, IMI CAB International Biosciences, Egham, UK, NBRC IFO, Institute for Fermentation, Osaka, Japan, NITE Biological Resource Centre, Japan, UAMH University of Alberta Microfungus Collection and Herbarium, G, A, Aphanoascus, K Keratinophyton, Ct Ctenomyces, ^T^ ex-type culture.
Phylogenetic analysis
All sequences were aligned with MAFFT v7 with default settings. The percent similarity between strains was determined using BioEdit v7.2^23^. ModelFinder^24^ on IQ-TREE web server^25^ was used to find the best-fitting model for ITS and LSU datasets according to the Bayesian Information Criterion (BIC). Phylogenetic trees were constructed using the maximum likelihood (ML) methods implemented in IQ-TREE web server. Branch support values were measured using ultrafast bootstraps. Additionally, MrBayes v3.2.7^26^ with default settings on the CIPRES portal (http://www.phylo.org/) was used for both datasets. Ctenomyces serratus CBS 187.61 (Arthrodermataceae, Onygenales) was used as an outgroup. Phylogenetic trees were displayed and edited using Treeview v1.6.6^27^ and iTOL v6^28^.
Results
Morphological analyses and keratin degradation
The results of the morphological analyses are given for each novel species under the Taxonomy section below. Temperature dependent growth of the new Keratinophyton species on PDA, MEA and SDA after 14 days are provided in Table S1a‒c. Briefly, K. keniense grew better than K. kautmanovae on the same type of media and at the same incubation temperatures. All species showed good growth at 20‒25 °C on all three media. K. kautmanovae does not grow at 30 °C, while K. keniense reached up to 38 mm after 14 days at this temperature.
Ability to digest keratin was observed in the two new species after 21 days on testing medium (YEW). However, attack intensity on the hair according to the scale of^18^ Marchisio et al. was detected to be very weak in both species (= 0–1). Urease activity was negative in both new species on Christensen ´s urea agar.
Phylogenetical analysis
The ITS dataset consisted of 48 strains with 541 sites, LSU dataset consisted of 38 strains with 564 sites, and the ITS-LSU combined dataset included 38 strains with 564 sites. The best-fitting model was TNe + I + G4 for both datasets. Phylogenetic analyses of ITS (Fig. 1a) and combination of ITS-LSU data (Fig. 1b) of the species described in Keratinophyton and Aphanoascus revealed that strains BiMM-F297 and BiMM-296 (Keratinophyton kautmanovae sp. nov.) formed a basal clade in the genus Keratinophyton with 100% support. Strain BiMM-F335 (Keratinophyton keniense sp. nov.) was clustered together with K. hubeiense and K. sichuanense with 94% and 96% ITS similarity, respectively.Figure 1(a) Phylogenetic tree of Keratinophyton and Aphanoascus strains based on ITS sequences. The new taxa of Keratinophyton are compared with available sequences of the other related species together with their conidium size, presence of intercalary conidia and ability to grow at 37 °C. Empty triangles and squares represent the absence of the characteristics. (b) Phylogenetic tree based on combination of ITS and LSU sequences for the new taxa of Keratinophyton together with available sequences of the other related species. Numbers at nodes indicate Bayesian probability/ maximum likelihood bootstrap values (≥ 60%). New species are shown in bold. Ctenomyces serratus was used as outgroup. A sequence for K. multiporum was not available for the study. T, ex-type culture.
Taxonomy
Keratinophyton kautmanovae B.Voleková, Kubátová, Kandemir & Labuda, sp. nov.
(Figs. 2 and 3)Figure 2Keratinophyton kautmanovae (BiMM-F297). (a) Colonies on PDA (after 14 days) at 25 °C (left—obverse, right—reverse). (b,c) Conidiophores with aleurioconidia (phase contrast microscopy). (d) Aleurioconidia under light microscopy (on PDA, after 14 days). Bars = 10 µm (b,c), 5 µm (d).Figure 3. Line drawing of micromorphology of Keratinophyton kautmanovae (BiMM-F297). Conidiophores with young and mature aleurioconidia on PDA (at 25 °C, after 14 days). Branched conidiophore (at right) and unbranched conidiophore (at left) with sessile aleurioconidia. Fungus is not melanised. Bar = 10 µm.
MycoBank: MB851666
Etymology: Named in honour of Ivona Kautmanová, Department of Botany, Slovak National Museum-Natural History Museum, Bratislava, Slovak Republic, an expert in the fungal ecology and taxonomy of higher fungi.
Type: Slovak Republic, Malé Karpaty, Pezinok, Rudné Mines, (coordinates: 48°19′03.3"N 17°14′20.9"E), from a soil sample close (1 m) to stream of contaminated water of abandoned antimony Sb deposit, coll. B. Voleková, December 2021, isol. R. Labuda, February 2022, holotype PRM 957896 (dried culture in metabolically inactive state), culture ex-type BiMM-F297 = CCF 6679 = CBS 150893, ITS sequence, GenBank PP062810; LSU sequence, GenBank PP062954.
Description: Sexual morph not observed on any of the media used. Asexual morph on PDA (25 °C, 14 days, in dark). Vegetative mycelium of hyaline, septate, smooth-walled, sparsely to pronouncedly branched hyphae, 1.0‒2.0 µm diam. Racquet hyphae not observed. Conidia (aleurioconidia), hyaline, white in mass, thin-walled, mostly smooth to finely roughened, some also coarsely roughened or irregularly warty. Terminal and lateral conidia born on main fertile hyphae or from side branches of variable length, sessile or on short protrusions, often slightly swollen (ampuliform) and of variable length, solitary, 1‒3 (–5) per conidiogenous cell, smooth to verrucose (warty), thick-walled, obovate to clavate, 1-celled, (3.5‒)4.0‒5.0(‒6.5) × (2.0‒)2.0‒2.5(‒3.0) µm (mean = 4.5 ± 0.5 × 2.4 ± 0.2 µm, n = 50). Intercalary conidia (arthroconidia) not observed. Chlamydospores not observed.
Culture characteristics: Colonies on PDA 13‒15 mm diam at 25 °C, after 14 d, floccose to downy (mealy), with abundant sporulation, white to creamy, flat, slightly umbonate at the centre, with slightly radial colony margin submersed into agar, reverse yellowish with dark yellow centre, no pigment or exudate produced. At 30 °C, no growth (germination only). Colonies on SDA 7‒10 mm diam at 25 °C, after 14 days, morphology similar to when on PDA with more floccose and plane colony surface and radial colony margin, poor sporulation, with orange reverse. At 30 °C, no growth (no germination). Colonies on MEA 15‒20 mm diam at 25 °C, after 14 days, morphology similar to PDA with more floccose colonies and plane structure, with moderate sporulation, limited hyaline exudate present, with yellow-orange reverse and vivid orange centre. At 30 °C, no growth (germination only). No ascomata observed after prolonged incubation (3 months).
Optimum temperature for growth on PDA, SDA and MEA at 20–25 °C. Minimum growth (2–5 mm in diam) at 12 °C. Germination of the conidia observed at 10 °C. Maximum temperature for growth (1–3 mm in diam) at 28 °C. Keratinolytic activity very weak, with hair attack intensity = 0–1. Urease activity negative (after 20 days of incubation).
Diagnosis*: *Keratinophyton kautmanovae molecularly can be distinguished from other Keratinophyton species by ITS locus analysis. Combination of the following phenotypic features can be used to differentiate this fungus from other species in the genus: (1) obovoid-clavate, smooth to coarsely verrucose or irregularly warty conidia, (2) absence of arthroconidia, (3) generally slow grow at 25 °C, poor growth at 28 °C and no growth at 30 °C, and (4) orange reverse at 25 °C on MEA and SDA.
Additional material examined: Slovak Republic, Malé Karpaty, Pezinok, Rudné Mines, (coordinates: 48°19′03.3" N 17°14′20.9" E), isolated from a different sub-sample, February 2022, R. Labuda, a strain BiMM-F296 (ITS sequence, GenBank PP062811; LSU sequence, GenBank PP062955). Phenotypically and molecularly (ITS and LSU) identical with ex-type culture BiMM-F297.
Notes: Based on a search of NCBI GenBank nucleotide database, the closest hit for Keratinophyton kautmanovae using the ITS sequence is Keratinophyton lemmensii (ex—type CCF 6359 = BiMM—F 76; GenBank acc. MN633082), with identity = 463/504 (92%) and gaps 16/504 (3%). Phenotypically, K. kautmanovae can be readily distinguished from the K. lemmensii by absence of intercalary conidia (arthroconidia) and filiform 2-celled conidia, presence of orange colony reverse at 25 °C on MEA and SDA, slower growth at 25 °C and inability to grow at 30 °C.
Keratinophyton keniense V. Scheffenacker, A. Schüller, Kubátová, Kandemir & Labuda, sp. nov.
(Figs. 4 and 5)Figure 4Keratinophyton keniense (BiMM-F335). a Colonies on PDA (after 14 days) at 25 °C (left—obverse, right—reverse). (b) Conidiophores with aleurioconidia. (c–e) Aleurioconidia. (d) Arthroconidium—intercalary conidium (on PDA, after 14 days). Bars = 10 µm.Figure 5. Line drawing of micromorphology of Keratinophyton keniense (BiMM-F335). Conidiophores with young and mature aleurioconidia on PDA (at 25 °C, after 14 days). Branched conidiophore (right) and unbranched conidiophore (left) with sessile aleurioconidia. Fungus is not melanised. Bar = 10 µm.
MycoBank: MB851667
Etymology: Named according to the country of origin, Kenya, where the holotype was collected.
Type: Republic of Kenya, Njoro, Egerton University campus (approximate coordinates: 0°22′09.3" S 35°55′24.2" E), from a soil sample (top layer, 20 cm), February 2022, coll. Andreas Schüller, isol. V. Scheffenacker, July 2023, holotype PRM 960013 (dried culture in metabolically inactive state), culture ex-type BiMM-F335 = CCF 6712 = CBS, ITS sequence, GenBank PP062809; LSU sequence, GenBank PP062953.
Description: Sexual morph not observed on any of the media used. Asexual morph on PDA (25 °C, 14 days, in dark). Vegetative mycelium of hyaline, septate, smooth-walled, sparsely to pronouncedly branched hyphae, 2.0‒5.0 µm diam. Racquet hyphae present, rare. Conidia (aleurioconidia) hyaline, white in mass, thick-walled, smooth to very finely roughened. Terminal and lateral conidia born on main fertile hyphae or from side branches of variable length, sessile or on short protrusions of variable length, solitary, 1‒3(–7) per conidiogenous cell, obovate to clavate, mostly 1-celled, smooth, thick-walled, often apiculate and cymbiform (boat-like shaped), (4.0‒)5.0‒6.0(‒7.0) × (2.0‒)2.5‒3.0(‒3.5) µm, (mean = 5.7 ± 0.8 × 2.8 ± 0.3 µm, n = 60), some also 2-celled, oblong to cylindrical, up to 18 µm long, conidia at colony centre often (sub-)globose and rhomboid, (4.5‒)5.0‒6.0(‒7.0) × (4.0‒)4.5‒5.5(‒6.0) µm (mean = 5.7 ± 0.4 × 4.7 ± 0.3 µm, n = 60). Intercalary conidia (arthroconidia) very rare. Chlamydospores not observed.
Culture characteristics: Colonies on PDA 35‒38 mm diam at 25 °C, after 14 days, floccose to downy (mealy), with abundant sporulation, white to creamy, flat, slightly umbonate at the centre with a few concentric rings, even regular colony margin submersed into agar, reverse yellow with dull yellow centre, no pigment or exudate produced. At 30 °C, 33‒37 mm diam. Colonies on SDA 37‒40 mm diam at 25 °C, after 14 days, morphology similar to when on PDA, good sporulation, with darker yellow reverse at the centre. At 30 °C, 35‒38 mm diam. Colonies on MEA 43‒45 mm diam at 25 °C, after 14 days, powdery to downy (mealy), with abundant sporulation, white to creamy, flat, slightly umbonate at the centre, margin irregular, reverse yellow to dull yellow orange, no pigment or exudate produced. At 30 °C, 29‒31 mm diam. No ascomata observed after prolonged incubation (3 months).
Optimum temperature for growth on PDA, SDA and MEA at 20–30 °C. Minimum growth (2–6 mm in diam) at 10 °C. Germination of the conidia observed at 8 °C. Maximum temperature for growth (2–5 mm in diam) at 33 °C. Germination of the conidia and formation of microcolonies observed at 34 °C. Keratinolytic activity absent, with hair attack intensity = 0. Urease activity negative (after 20 days of incubation).
Diagnosis*: *Keratinophyton keniense molecularly can be distinguished from other Keratinophyton species by ITS locus analysis. Combination of the following phenotypic features can be used to differentiate this fungus from other species in the genus: (1) cymbiform and rhomboid conidia, (2) arthroconidia very rare, (3) good grow at 20–30 °C, (4) dull yellow reverse at 25 °C.
Notes: Based on a search of NCBI GenBank nucleotide database, the closest hit for Keratinophyton keniense using the ITS sequence is Keratinophyton sichuanense (ex—type CGMCC 3.20871; GenBank acc. NR182583), with identity = 529/550 (96%) and gaps 1/550 (0%). Phenotypically, K. keniense can be readily distinguished from the K. sichuanense by presence of rhomboid conidia, slower grow on PDA at 25 °C after 14 days (35‒38 vs 50‒54 mm), and inability to grow at 37 °C.
Discussion
Phylogeny
Phylogenetic reconstruction using ITS and LSU sequences (Fig. 1) resulted in clustering the both new taxa, Keratinophyton kautmanovae and* K. keniense*, with eight currently accepted species, namely K. alvearium^14^, K. durum^2^, K. chongqingense^8^, K. hubeiense^12^, K. lemmensii^7^, K. sichuanense^8^, K. siglerae^29^, and K. submersum^11^. Apart from the pronounced differences in the ITS regions, the species mentioned above can be distinguished by particular combinations of their phenotypic traits (e.g., colony characteristics, and morphology of conidia) as listed in Table 2. The monophyletic genus Keratinophyton is now extended and includes 29 species including six species known from sexual morphs^5^ and 23 species which are currently known only from asexual morphs^8^ and this study. The ability to produce ascosporic state (sexual morph) in vitro within this cluster is confined to K. durum^2^, characterized by discoid ascospores with flattened poles and with a broad equatorial rim, cruciform in side view, broad-ridged, with reticulate surface^4^. As noted by Li et al. (2022)^8^, the ability to form sexual morph in vitro is not phylogenetically conserved, as it can be seen from the phylogenetic analysis, showing that all known species forming ascosporic (sexual) structures within the genus are not clustered together and they are spread over the phylogenetic tree^6–8^.Table 2. Comparison of the key phenotypic characteristics of the phylogenetically close related Keratinophyton spp.SpeciesReverse colony on PDA 25 °C, after 14 daysIntercalary conidiaConidial shapeConidial dimensions (µm)Conidial surfaceReferencesK. kautmanovaeYellowish, dark yellow centrallyAbsentObovate to clavate3.5‒6.5 × 2.0‒3.0 (1-celled)Smooth to verrucoseThis studyK. kenienseYellowish, dull yellow centrallyPresent^b^Obovate to clavate, cymbiform, filiform, globose, rhomboid4.0‒18.0 × 2.0‒6.0 (1- to 2-celled)SmoothThis studyK. alveariumWhitePresent^c^Globose, pyriform, clavate, cylindrical4.0‒9.0 × 2.0‒7.5 (1-celled)SmoothZhao et al., 2018^14^K. chongqingenseWhitePresentSubglobose, globose, obovate, ellipsoidal apiculate3.0‒10.0 × 2.0‒4.5 (1-celled)Smooth to slightly verrucoseLi et al., 2022^8^K. durum^a^ColorlessAbsentPyriform to clavate4.5‒7.7 × 2.0‒3.0 (1-celled)SmoothGuarro et al. 2012^4^K. hubeienseYellowishAbsentObovoid to ellipsoidal2.0‒4.5 × 1.5‒3.0 (1-celled)SmoothZhang et al. 2016^43^K. lemmensiiLemon yellow (with bright yellow pigment)PresentClavate to filiform3.0‒40.0 × 1.5‒4.0 (1- to 2-celled)SmoothLabuda et al., 2021^7^K. sichuanenseWhiteAbsentObovate to clavate, sometimes curved4.0‒8.5 × 2.0‒4.0 (1-celled)Smooth to slightly verrucoseLi et al., 2022^8^K. sigleraePale brownPresentCylindrical to clavate5.0‒30.0 × 2.0‒3.5 (1- to 2-celled)Smooth to slightly verrucoseCano et Guarro, 1994^29^K. submersumYellowish whitePresent^d^Clavate, pyriform, obovoid and subglobose4.0‒35.0 × 2.5‒5.0 (1- to 4-celled)Smooth to verrucoseVidal et al., 2002^11^^a^Forming sexual morph (ascospores) in vitro.^b^Intercalary conidia observed only very rarely.^c^Intercalary conidia abundant, solitary or in chains.^d^Intercalary conidia present in older cultures.
Ecology and distribution
Almost all known Keratinophyton species have been isolated from soil or soil-like substrates, such as river sediments, compost and sand, and as non-pathogenic fungi as a result of mycological screening for so-called keratinophilic/keratinolytic fungi using a horse-hair baiting method^7,8^. This highly selective method was introduced by Vanbreuseghem (1952) for soil fungi having affinity to keratinous material especially for onygenalean fungi such as dermatophytes^19,30^. According to Papini et al. (1998), Ajello reviewed the taxonomy of keratinophilic fungi for the first time in 1968^31^. Later, Otčenášek et al. (1969) reported on the worldwide distribution of keratinophilic mycobiota in soil, claiming that the occurrence of keratinophilic fungi in soil depends on the presence of mammals, birds, and humans in a variety of ecological sites^32^. It is in fact the only method how this group of keratinophilic fungi can be isolated from the soil-like substrates and studied further. In this study, both fungi originated from the areas which are freely accessible by wild animals typically inhabiting these regions, and thus, these soils might be presumably reach on source of keratin as well as a source of biodiversity for this specific fungal group regardless of geochemical properties of soils^15,33,34^. As for the sample from the abandoned antimony (Sb) deposit (Slovakia, EU), the soil sample is rich in iron oxides and is also characterized by elevated concentrations of arsenic, antimony, aluminum and sulfates. More about mineralogy and geochemistry of the studied site was published previously^35^. On the other hand, the soils in Egerton Njoro area are Vintric Mollic andosols^36,37^. The sampled area is native, not influenced by agricultural or any industrial activities.
As mentioned above, the genus Keratinophyton harbours a total of 29 species, including the two new species described in this study. The majority of the new taxa have been so far described from Europe (15 spp.), followed by Asian continent (China, 8 spp. and India, 5 spp.)^5–8^. To the best to our knowledge, the description of K. keniense represents the first taxonomical novelty of the genus Keratinophyton from African continent. Thus, further research is needed because unknown strains may be isolated from similar environments on the African continent.
Hubálek provided a list of keratinolytic fungi associated with free-living mammals and birds of which ubiquitous K. durum, K. pannicola and K. terreum have been isolated from a variety of animals and from different geographical regions^38^. There are only a few reports of a human or animal clinical isolate belonging to Keratinophyton^16,39^, however, all these cases have doubtful etiological history and with no solid evidence of their pathogenicity^16,40^. On the other side, K. pannicola (as C. pannicola) is included in the Atlas of Clinical Fungi^17^ as a concern in skin infections. Even though the keratinophilic fungi were considered as potential pathogens by several researchers^41,42^; they rarely cause infections. Therefore, soil is proposed as an epidemiological and probably also an evolutionary link, that relates geophilic, zoophilic, and anthropophilic keratinophilic fungi^42^. Although being cycloheximide resistant, a potential pathogenicity to homeothermic vertebrates (mammals and birds) by these fungi seems highly unlikely because both new species are not able to grow at higher temperature (above 30‒34 °C), they are urease negative, and possess none or very mild keratinolytic activity in vitro. Rather contrary, these fungi might be interesting from a metabolic point of view, as they undoubtedly represent a yet unexplored source of new bioactive compounds as there is not much known of these properties in the genus^34^. Metabolic profile and investigation of the potential use of substances produced by these two novel fungi is an object of our further biochemical exploration.
Supplementary Information
Supplementary Table S1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Randhawa HS Sandhu RS Keratinophyton terreum gen. nov. sp. nov., a keratinophilic fungus from soil in India Sabouraudia 1964325125710.1080/003621764851904215896103 · doi ↗ · pubmed ↗
- 2Cano J Guarro J The genus Aphanoascus Mycol. Res.19909435537710.1016/S 0953-7562(09)80361-4 · doi ↗
- 3Cano J Molecular taxonomy of Aphanoascus and description of two new species from soil Stud. Mycol.200247153164
- 4Guarro, J., Gene, J., Stchigel, A. M. & Figueras, M. J. Atlas of Soil Ascomycetes. (CBS-KNAW Fungal Biodiversity Centre, 2012).
- 5Sutton DA Isolation and characterization of a new fungal genus and species, Aphanoascella galapagosensis, from carapace keratitis of a Galapagos tortoise (Chelonoidis nigra microphyes)Med. Mycol.20135111312010.3109/13693786.2012.70176722852752 · doi ↗ · pubmed ↗
- 6Kandemir, H. et al.Phylogenetic and Ecological Reevaluation of the Order Onygenales (2022).
- 7Labuda R Bernreiter A Hochenauer D Kubatova A Kandemir H Schuller C Molecular systematics of Keratinophyton: The inclusion of species formerly referred to Chrysosporium and description of four new species Ima Fungus 202110.1186/s 43008-021-00070-234233753 PMC 8265132 · doi ↗ · pubmed ↗
- 8Li X Zhang ZY Chen WH Liang JD Liang ZQ Han YF Keratinophyton chongqingense sp. nov. and Keratinophyton sichuanense sp. nov., from soil in China Int. J. Syst. Evol. Microbiol.202210.1099/ijsem.0.00546835930466 · doi ↗ · pubmed ↗