A new species of holothuroid from the Labrador Sea (eastern Canada): Pseudothyonelabradorensis sp. nov. (Echinodermata, Holothuroidea, Dendrochirotida, Sclerodactylidae)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
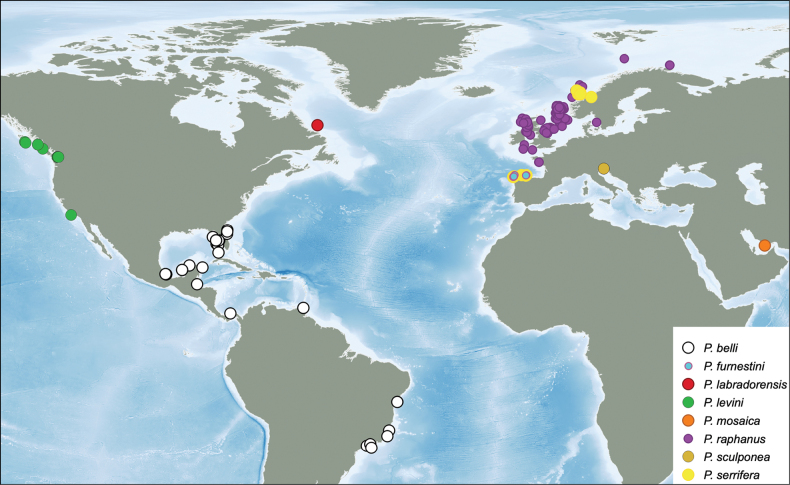 Figure 1
Figure 1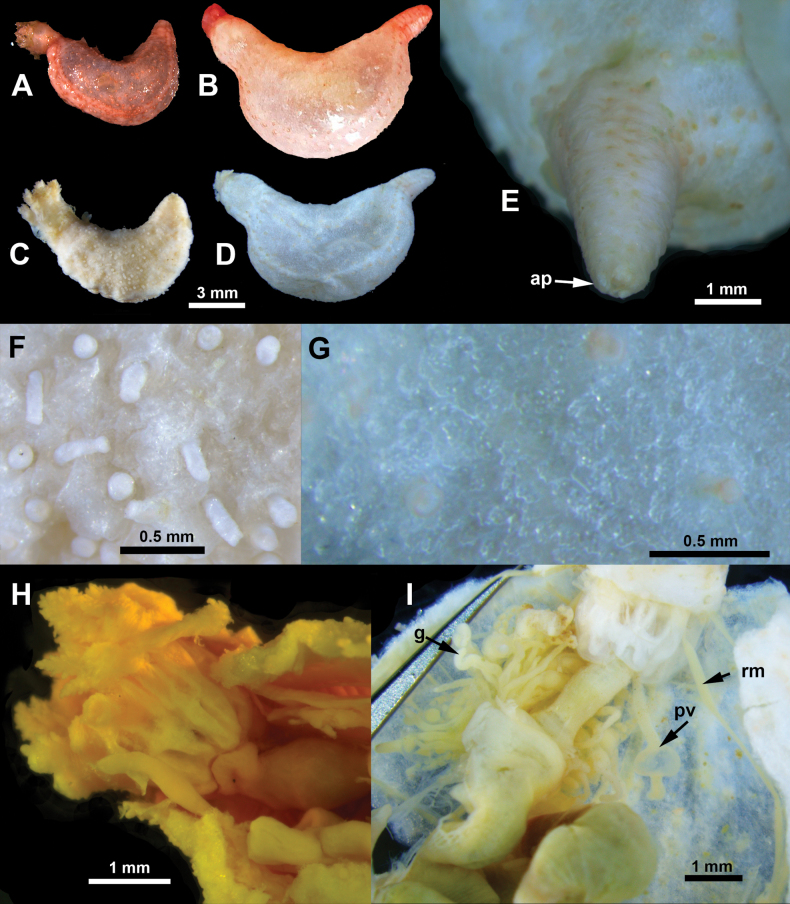 Figure 2
Figure 2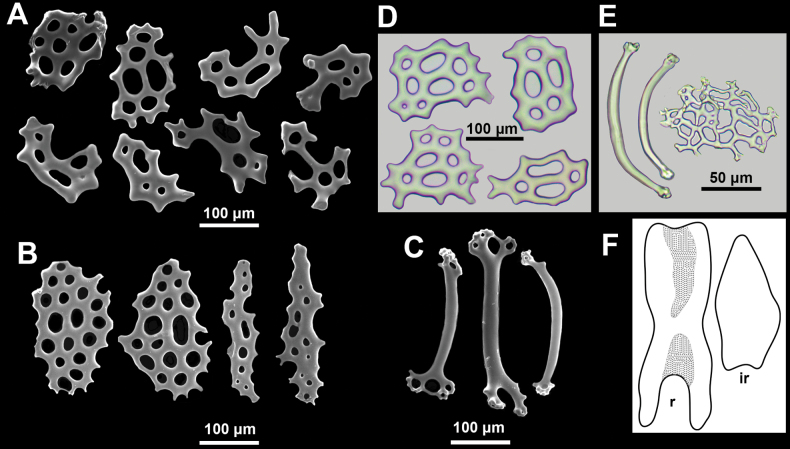 Figure 3
Figure 3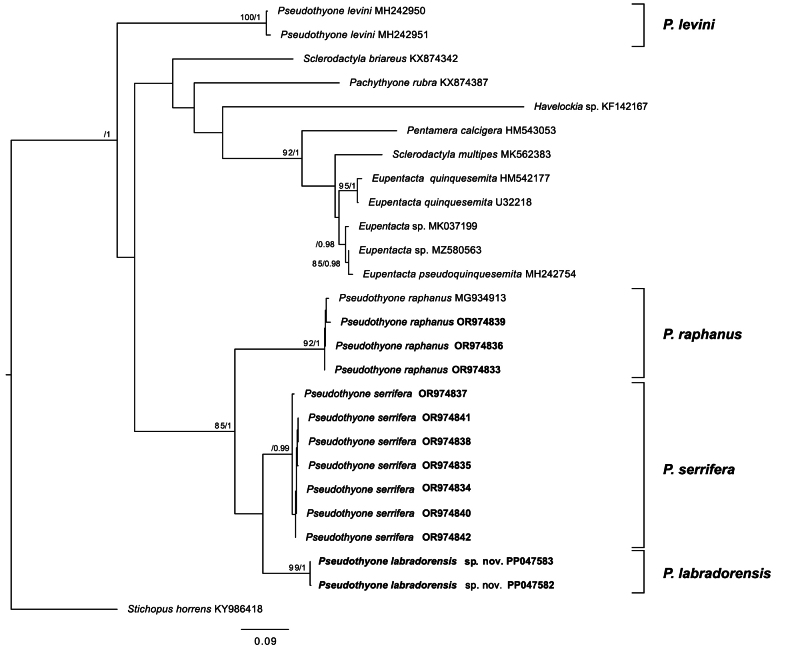 Figure 4
Figure 4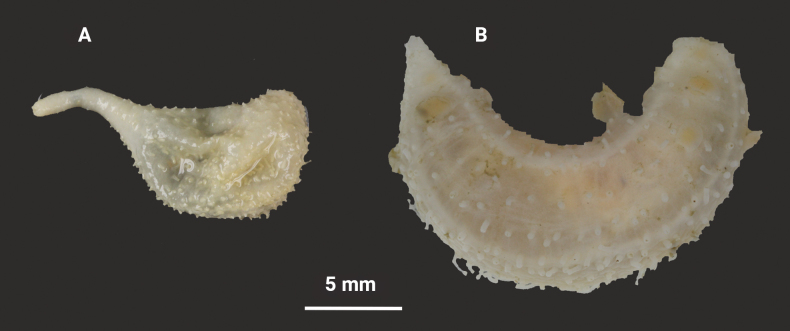 Figure 5
Figure 5| Identification | Voucher | GenBank Acc. | BOLD Process ID | Catalog Number | Storing Institution* | Collection Date | Region | Latitude | Longitude | Depth (m) |
|---|---|---|---|---|---|---|---|---|---|---|
| AMLAB-02 |
|
|
| 30/08/2020 | coast of Labrador (eastern Canada) | 56.500 | -58.084 | 740–969 | ||
| PT1210 |
|
|
| 30/08/2020 | coast of Labrador (eastern Canada) | 56.500 | -58.084 | 740–969 | ||
|
| Sletvik2016_87 |
|
|
| 25/10/2016 | Norway, Trondheimsfjorden | 63.67830 | 9.79612 | 122 | |
|
| ZMBN_120538 |
|
|
| 08/06/2014 | Norway, Halsnøyfjorden | 59.75777 | 5.49778 | 60 | |
|
| ZMBN_120547 |
|
|
| 02/05/2006 | Norway, Kobbaleia, Bergen area | 60.314 | 5.156 | 22–42 | |
|
| Sletvik2016_111 |
|
|
| 25/10/2016 | Norway, Trondheimsleia | 63.657 | 9.634 | 260–278 | |
|
| Sletvik2016_88 |
|
|
| 24/10/2016 | Norway, Trondheimsfjorden | 63.586 | 9.846 | 280–102 | |
|
| Sletvik2016_89 |
|
|
| 24/10/2016 | Norway, Trondheimsfjorden | 63.586 | 9.846 | 280–102 | |
|
| Sletvik2016_90 |
|
|
| 24/10/2016 | Norway, Trondheimsfjorden | 63.586 | 9.846 | 280–102 | |
|
| Sletvik2016_91 |
|
|
| 25/10/2016 | Norway, Trondheimsleia | 63.657 | 9.634 | 260–278 | |
|
| Sletvik2016_92 |
|
|
| 25/10/2016 | Norway, Trondheimsleia | 63.657 | 9.634 | 260–278 | |
|
| Sletvik2016_86 |
|
|
| 26/10/2016 | Norway, Trondheimsleia | 63.594 | 9.508 | 56–45 |
| Identification | Current family attribution | Voucher | GenBank Acc. |
|---|---|---|---|
|
|
| BIOUG<CAN>: |
|
|
|
| no data |
|
|
|
| BMBM-0776 |
|
|
| ECHINO_001_091 |
| |
|
| USNM:IZ:1503386 |
| |
|
| isolate CZ1 |
| |
|
| NMV_F151829 |
| |
|
|
| SIO:BIC:E6676 |
|
|
|
| BIOUG<CAN>:HLC-30032 |
|
|
|
| BFHL-1914 |
|
|
|
| BMBM-1095 |
|
|
|
| Echin 6852V |
|
|
|
| SIO:BIC:E6814 |
|
|
|
| isolate SHP3 |
|
| 1 | Tube feet present on tapered posterior part of body |
|
| – | Tube feet absent or few on tapered posterior part of body |
|
| 2 | Body-wall ossicles surrounding tube feet with elongated non-perforated prolongations; terminal plate in tube feet star-shaped |
|
| – | Body-wall ossicles lack prolongations; terminal plate of irregular shape or absent |
|
| 3 | Body-wall ossicles smooth plates and knobbed buttons |
|
| – | Body-wall ossicles smooth plates only |
|
| 4 | Tube feet ossicle terminal plate only |
|
| – | Tube feet with supporting rods and terminal plate |
|
| 5 | Body-wall plate perforations with serrated margins |
|
| – | Body-wall plate perforations smooth |
|
- —Canadian Network for Research and Innovation in Machining Technology, Natural Sciences and Engineering Research Council of Canada 501100002790 http://doi.org/10.13039/501100002790
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEchinoderm biology and ecology · Marine and coastal plant biology · Marine Biology and Ecology Research
Introduction
The genus Pseudothyone was established by Panning (1949) for Thyoneraphanus Düben & Koren, 1846 within the new subfamily Sclerodactylinae. Apart from the type species, Panning assigned five other species to this genus: P.belli (Ludwig 1886), P.mosaica (Koehler & Vaney, 1910), P.poucheti (Barrois, 1882), P.argillacea (Sluiter, 1910), P.buccalis (Stimpson, 1855) and P.trachyplaca (Clark, 1924). The latter two species were later assigned to other genera, whereas P.poucheti was synonymized with P.raphanus according to Théel (1886), and P.argillacea with P.belli according to Deichmann (1954).
Pseudothyone currently includes seven species characterized by a wide range of morphological characters. The original diagnosis by Panning (1949) included the following: ten tentacles, undivided pieces of calcareous ring and its radial pieces with two fork-shaped processes of medium length undivided or consisting of few large pieces, and body-wall ossicles composed of only plates. Later investigations of this genus added more variations to taxonomic characters (Lambert and Oliver 2001; Martins 2019): tentacles ten equal-sized (in P.levini Lambert & Oliver, 2001) or eight bigger and two ventral smaller in all other species; body-wall ossicles smooth plates and knobbed buttons (P.belli), or smooth plates only (all other species); tube feet ossicles tables and end plates (P.belli and P.mosaica), rods and end plates (P.sculponea Cherbonnier, 1958, P.serrifera (Östergren, 1898) and P.levini), or end plates only (P.raphanus and P.furnestini Cherbonnier, 1969); radial pieces of calcareous ring with short posterior processes (P.levini), or of medium length (all other species), with posterior processes undivided (P.raphanus), or divided into several pieces (P.belli, P.mosaica, P.sculponea).
Most species of Pseudothyone are distributed in the Atlantic Ocean (Fig. 1). Pseudothyonefurnestini, P.raphanus, and P.serrifera occur in the Northeast Atlantic, and the latter two are also known from the Mediterranean. Pseudothyonesculponea is known only from the Mediterranean, and P.belli is from the Western Atlantic (Atlantic US coast, Caribbean and Brazil). Moreover, two species are known outside the Atlantic: P.mosaica from the Persian Gulf, and P.levini from the northeastern Pacific. Bathymetric distribution also differs between the species. The shallowest is P.belli occurring at sublittoral depths from the low-tide mark to 37 m (Pawson et al. 2010). Among shallow-water representatives are also P.levini occurring from the intertidal to 70 m depth and P.sculponea reported from 21 to 41 m (GBIF.org 2023). Pseudothyoneraphanus and P.serrifera occur deeper, at 10–1200 m and 200–1200 m, respectively (Madsen and Hansen 1994; Fernández-Rodríguez et al. 2019). The deepest-dwelling species is P.furnestini reported from 440–1347 m (Fernández-Rodríguez et al. 2019). Pseudothyonemosaica is known from a single record at 97 m.
Distribution of Pseudothyonelabradorensis sp. nov. and other species in the same genus, based on published records and GBIF (2023). Map was prepared using QGIS 3.16.5-Hannover.
Phylogenetic relationships of species within the genus Pseudothyone, as well as the latter’s position within Sclerodactylidae, remain unclear. The taxon was originally described by Panning (1949) as Sclerodactylinae, a subfamily of Cucumariidae. Later, Pawson and Fell (1965) upgraded its status to the family Sclerodactylidae with two subfamilies, Sclerodactylinae and Cladolabinae. Thandar (1989) restricted the diagnosis of Sclerodactylidae and described a new subfamily, Sclerothyoninae. Smirnov (2012) recognized the subfamilies Cladolabinae and Sclerothyoninae as separate families. Available molecular data (Miller et al. 2017) partly support the system of Smirnov, showing no sister relationships between sclerothyonins Afrocucumisafricana (Semper, 1867) and Euthyonidiellahuwi O’Loughlin in O’Loughlin et al., 2014 and sclerodactylins Pachythyonerubra (Clark, 1901) and Sclerodactylabriareus (Lesueur, 1824).
In this study we describe a new species, Pseudothyonelabradorensis sp. nov., from the bathyal depths of the Labrador Sea (Northwest Atlantic Ocean) based on morphological and molecular data. Using molecular data on partial sequences of the mitochondrial gene cytochrome c oxidase subunit I (COI), we examined phylogenetic relationships of P.labradorensis sp. nov. with two Atlantic congeners, P.raphanus (type species of the genus) and P.serrifera, for which we obtained additional genetic data, as well as with the Northeast Pacific species P.levini. COI is commonly used for recovering relationships within the genera of Holothuroidea (O’Loughlin et al. 2014; Li et al. 2018; Ogawa et al. 2022; Ogawa et al. 2023). To test monophyly of Pseudothyone we included available data on the representatives of Sclerodactylidae sensu Smirnov (2012) as well as Pentameracalcigera (Stimpson, 1851), as close relationships of Pentamera to Sclerodactylidae has been shown previously (Arndt et al. 1996; Miller et al. 2017).
Material and methods
Two specimens of Pseudothyonelabradorensis sp. nov. were collected together in the same location (North Atlantic Fisheries Organization NAFO, Zone 2J https://www.marineregions.org/gazetteer.php?p=details&id=23382) using a rock dredge deployed during the ISECOLD scientific expedition aboard the research icebreaker CCGS Amundsen on 30 August 2020 (Table 1). The rock dredge net had a 7 mm mesh. Due to the large volume of mud collected at this station, a fourth of the material was sieved through a 2 mm sieve and the rest was scanned for larger individuals through a 17 mm diameter mesh sieve. The specimens later identified as Pseudothyone were preserved in 100% ethanol for morphological and molecular analysis.
The specimens of P.raphanus and P.serrifera had previously been collected in 2006–2016 with a Van Veen grab, Triangular dredge and Agassiz trawl deployed from the research vessels R/V Gunnerus, R/V Håkon Mosby and R/V Hans Brattström (Table 1). They were preserved and subsequently stored in 96% ethanol.
Morphological examination, dissection and photographing were performed using a Leica M205C stereomicroscope equipped with a Leica FLEXACAM C1 digital camera. To extract ossicles, small fragments of the body wall, introvert, tube foot and tentacle skin were digested in a domestic bleach water solution followed by several rinses in distilled water. For light microscopy, ossicles were transferred onto a glass slide, dried using a heating stage and mounted in Canada Balsam. For scanning electronic microscopy (SEM), ossicles were dried with 96% ethanol, mounted on a stub and sputter coated with gold. Ossicles were examined and photographed under a light microscope (Olympus BX43) with a ToupCam U3CMOS08500KPA digital camera and SEM examination was performed using a TESCAN Vega 3.
To evaluate phylogenetic relationships of Pseudothyonelabradorensis sp. nov. within the genus, partial sequences of cytochrome c oxidase, subunit I (COI) were obtained from two examined specimens. Additionally, COI data were obtained for three specimens of P.raphanus and seven specimens of P.serrifera (Table 1). Some of these specimens were collected close to their type localities (Bergen area and Trondheimsfjorden, respectively). Also, three sequences of Pseudothyone, publicly available in GenBank, were used in the analysis: P.raphanus (MG934913) and P.levini (MH242951 and MH242950). To test monophyly of Pseudothyone, published sequences of Sclerodactylidae sensu Smirnov (2012) were analysed (Table 2). A COI sequence of Pentameracalcigera (Phyllophoridae) was also included in the analysis as it was recovered in a sister clade to the sclerodactylid Pachythyonerubra (Miller et al. 2017). Stichopushorrens was set as an outgroup. GenBank accession numbers of the sequences used in the analysis are listed in the Table 2.
Molecular work was carried out in two laboratories applying two different protocols. Genetic data on Pseudothyonelabradorensis sp. nov. (voucher AMLAB-02), P.raphanus and P.serrifera were obtained at the Canadian Centre for DNA Barcoding, University of Guelph following protocols by Ivanova et al. (2006), Ratnasingham and Hebert (2007), and deWaard et al. (2008). Data on another specimen of P.labradorensis sp. nov. (voucher PT1210) was generated at IORAS using the following methods: DNA was extracted using QuickExtract^TM^ DNA Extraction Solution (Lucigen) following the manufacturer protocol; PCR amplification was conducted using Encyclo Plus PCR kit (Evrogen, Moscow) according the manufacturer protocol with annealing temperature set at 48 °C; PCR products were purified from agarose gel using HiPure Gel DNA Mini Kit (Magen); the purified samples were sequenced using the Sanger method on Applied Biosystems ABI 3900 (ThermoFisher Scientific) by Evrogen (Moscow, Russia). All PCR amplifications and sequencing were carried out using the LCOech1aF1 (5′-TTTTTTCTACTAAACACAAGGATATTGG-3′; D. Eernisse unpublished) and HCO2198 (5′-TAAACTTCAGGGTGACCAAAAAATCA-3′; Folmer et al. 1994) primers.
Contigs were assembled from forward and reverse chromatograms using the MUSCLE algorithm implemented in Geneious v.10.0.9 and then manually edited. Sequences were aligned in MEGA 7 (Kumar et al. 2016) also using the MUSCLE algorithm, and then checked for stop-codon presence. The final dataset included 840 aligned positions. Phylogenetic analysis was performed using maximum-likelihood (ML) and Bayesian inference (BI) approaches. PartitionFinder 2 (Lanfear et al. 2017) was used for selecting best-fit partitioning schemes and models of nucleotide evolution. The defined models were TRNEF+I+G for positions 1 and 2, and GTR+G for position 3. ML tree search and bootstrapping was conducted in RAxML-NG (Kozlov et al. 2019) using auto MRE option with cutoff=0.03; the analysis converged after 12950 replicates. BI analysis was performed using MrBayes v.3.2 (Ronquist et al. 2012). The analysis was conducted in two runs, four chains (one cold and three heated) with trees and parameters sampled every 500 generations. The traces were analysed in Tracer v.1.7.1, and then 10% of the trees were discarded as burn-in. Run convergence was evaluated by analysing sump output parameters in MrBayes log file and Tracer v.1.7.1. Genetic distances were calculated using Kimura 2-parameter model (K2P; Kimura 1980) implemented in MEGA 7.
Taxonomy
Order Dendrochirotida Grube, 1840
Family Sclerodactylidae Panning, 1949 sensu Smirnov, 2012
Pseudothyone Panning, 1949
Pseudothyone
labradorensis
sp. nov.
Taxon classificationAnimaliaDendrochirotidaSclerodactylidae
9B64AAFE-BD53-5697-8C2A-D3FE095FFB90
https://www.zoobank.org/B4110DB7-0589-4A96-BC32-344F56ABF794
Type material.
Holotype. Canada • 9.5 mm in length; Labrador Sea, 56.500, -58.084, depth 740–969 m (between bottom and recovery); 30 Aug. 2020; Station ISECOLD -0-1000; rock dredge; substratum primarily mud with sparse rocks; IORAS ECH02801, ID AMLAB-02. Paratype. Canada • 14 mm in length, collected at same time and locality as holotype; IORAS ECH02802, ID PT1210. Both holotype and paratype are preserved and stored in 96% ethanol.
Diagnosis.
Body curved, cylindrical, tapered at anterior and posterior ends. Body colour in vivo pinkish. Tentacles 10, two ventral tentacles smaller. Tube feet arranged in several rows along radii, also present in interradii and on tapered posterior part of body. Body-wall ossicles slightly concave plates of irregular shape with smooth margins and perforations. Ossicles of tube feet rod-shaped; terminal plate irregular in shape; rods not numerous, smooth, with enlarged tuberculous ends. Tentacle ossicles rods with enlarged perforated ends. Segments of calcareous ring with undivided posterior projections.
Description.
Body curved towards dorsal side, wider and cylindrical in the middle, anterior end tapered towards terminal mouth, posterior end tapered to a ‘tail’ towards terminal anus, ‘tail’ short, more prominent in paratype and short in holotype (Fig. 2A–D). Body colour pinkish in living specimens (Fig. 2A, B), tentacles, tube feet and ‘tail’ more brightly coloured; colour in ethanol uniformly greyish, tentacles and tube feet whitish or greyish (Fig. 2C, D). Body length 9.5 mm in holotype, 14 mm in paratype. Body-wall skin thin, rough, non-transparent, with dense layer of scale-like ossicles (Fig. 2G). Tentacles (examined in holotype, in paratype they were partly retracted) ten, two ventral tentacles remarkably smaller in size. Tube feet small (Fig. 2F), non-transparent in ethanol, arranged in several rows along radii and also scattered in interradii; tube feet more numerous on mid body, along ventrolateral and mid-ventral radii; on ‘tail’ tube feet smaller and less numerous, arranged in double rows (Fig. 2E). Anal papillae five (Fig. 2E). Pieces of calcareous ring up to 1.6 mm in length, united at most their length, radial segments with undivided posterior projections, with forked grooved anterior projections; interradial segments with grooved anterior projections (Fig. 2H, I). Retractor muscles undivided, not flat, broader anteriorly (Fig. 2I). Polian vesicle single, non-divided (Fig. 2I). Gonad in a tuft (Fig. 2I), gonad tubules in paratype with oocytes of different size.
Pseudothyonelabradorensis sp. nov., external and internal morphology A, C holotype, before preservation (A) and preserved in ethanol (C) B, D paratype, before preservation (B), in ethanol (D) E tapered posterior body part in paratype F tube feet in holotype G body-wall skin with dense layer of ossicles H segments of calcareous ring in holotype I dissected anterior part in paratype. Labels: ap anal papillae, g gonad, pv Polian vesicle, rm retractor muscle.
Body-wall ossicles in a single dense overlapping layer, laying their concave side out; body-wall ossicles small perforated plates, usually do not exceed 200 µm in length, slightly concave, mostly irregular in shape, smooth; perforations with smooth margins, from rounded to oblong in shape, their size and shape can vary even on a same plate (Fig. 3A, D). Introvert with bigger flat plates, plates often narrow and elongated (Fig. 3B). On tube feet supporting rod-shaped ossicles, not numerous, curved in shape, with slightly enlarged perforated ends, length 100–130 µm; terminal plate delicate, irregular in shape, ~70 µm in diameter (Fig. 3E). Tentacle ossicles curved rods, most ossicles ranged 100–480 µm in length, bigger in proximal part of tentacles; smaller rods sometimes enlarged in the middle; bigger rods often with enlarged spatulated and/or bifurcated ends (Fig. 3C).
Ossicles of Pseudothyonelabradorensis sp. nov. A, D body-wall perforated plates B introvert perforated plates C tentacle rods E tube foot rods and terminal plate F right dorsal radial (r) and interradial (ir) segments of calcareous ring. A, B, CSEM images D, E light microscope images F drawing.
Differential diagnosis.
Pseudothyonelabradorensis sp. nov. can be distinguished from other species by a combination of the following characters: pinkish body colour (may disappear after preservation in ethanol); tube feet arranged in double rows on tapered posterior end; body-wall ossicles only perforated plates with smooth perforation margins; introvert ossicles perforated plates; tube foot ossicles rods and terminal plate of irregular shape.
Distribution.
Canada, Labrador Sea, depth 740–969 m.
Etymology.
The species is named after its type locality.
Environmental information
A CTD cast and drop camera deployment took place at 494 and 218 m respectively from the rock dredge recovery location. Bottom water temperature at 990 m was 3.7 °C, salinity 34.8 psu. These conditions are associated with Labrador Sea Water, and are in contrast to colder and less saline conditions found on the adjacent continental shelf (Côté et al. 2019). Modelled bottom water velocities (GIOPS daily historical 3D data extracted from https://navigator.oceansdata.ca/public/ for 2022) for this collection location average 0.13 m s^-1^ (range: 0.02–0.23 m s^-1^). Other fauna collected in the rock dredge deployment included soft corals Duvaflorida (Rathke, 1806), sea pens (Pennatuloidea), fragments of the gorgonians Primnoaresedaeformis (Gunnerus, 1763), Paragorgiaarborea (Linnaeus, 1758), Acanellaarbuscula (Johnson, 1862), and the mushroom coral Anthomastus Verrill, 1878. Sponges, bivalves, and a small grenadier were also present, highlighting a diverse range of bottom type requirements. The seafloor imagery further corroborates the dominance of soft sediment in the study area, with sparse gravel and rocks. Based on 42 seafloor images (along a 1.1 km long transect), 98% of the primary sediment was classified as fine sediment, and only 2.4% as gravel.
Molecular data
Both examined specimens of Pseudothyonelabradorensis sp. nov. formed a single, well-supported clade [bootstrap (BS) 99, posterior probability (PP 1), Fig. 4]. The sequences of the holotype and paratype were assigned to the same Barcode Index Number (BIN, AEH8268) and showed little genetic divergence (K2P-distance 0.003). Based on the COI data, the closest taxa to P.labradorensis sp. nov. were P.serrifera (K2P-distance 0.10) and P.raphanus (K2P-distance 0.162). Phylogenetic analysis recovered P.labradorensis sp. nov. in a sister clade with P.serrifera, although this clade was weakly supported (BS 50, PP 0.84). A clade of P.raphanus, P.serrifera and P.labradorensis sp. nov. (raphanus clade) was well-supported in the BI analysis and averagely supported by ML (BS 85, PP 1). Pseudothyonelevini was sister to the clade with all other species of the dataset showing no close relationships with the raphanus clade nor with other examined sclerodactylids. The analysis showed no close relationships of the raphanus clade with examined sclerodactylid representatives.
Phylogenetic relationships inferred using maximum-likelihood (ML) and Bayesian (BI) analyses of the COI dataset (840 bp). Topology corresponds to best-scoring ML tree; node values are bootstrap scores (≥75%) / BI posterior probabilities (≥0.95). Taxa examined in this study are in bold. StichopushorrensKY986418 was set as outgroup.
Discussion
Molecular data supported close relationships of Pseudothyonelabradorensis sp. nov. to the type species of the genus, P.raphanus, and to the Northeast Atlantic species P.serrifera. According to body-wall ossicle morphology, P.labradorensis sp. nov. is most closely related to P.raphanus (Fig. 5A), from which it differs by its pinkish body colour, presence of tube feet on the ‘tail’, absence of fern-like ossicles on the tentacles, and by the presence of rod-shaped ossicles in the tube feet. From the genetically closest species, P.serrifera (Fig. 5B), the new species differs by the pinkish body colour, absence of S-shaped rods on tentacles, and by the smooth margins and perforations of body-wall ossicles. From another Northeast Atlantic species, P.furnestini, the new species differs by body colour and body-wall ossicles. Pseudothyonefurnestini is characterized by whitish colouration and thick body-wall ossicles that are from rounded to oval in shape and often possess solid, unperforated extensions. Also Pseudothyonefurnestini lacks rod-shaped ossicles in the tube feet.
Pseudothyoneraphanus (A) and P.serrifera (B) collected from the type locality areas. AZMBN 120547 BNTNU-VM-72187. Collection data is given in Table 1. Image courtesy of Katrine Kongshavn.
According to molecular data, Pseudothyonelevini was not closely related to the species of the raphanus clade. This species has ten tentacles that are equal in size, whereas other species of Pseudothyone have two ventral tentacles smaller than others. From species of the raphanus clade, it also differs by a less prominent ‘tail’, less perforated body-wall plates and by more robust rod-shaped ossicles on tube feet.
Some other species of Pseudothyone also have remarkable morphological differences. Apart from plates, the body-wall ossicles of P.belli include knobbed buttons and plates with handles, which do not occur in P.raphanus and most other species of the genus. Also P.belli differs by the ossicles from the tentacles, which are plates and rosettes (not rods as in P.raphanus and other species), by introvert ossicles pillared tables and plates, and by tube feet ossicles arched pillared tables. Therefore, P.belli differs remarkably from P.raphanus in most ossicle types. Marked differences in ossicle types are also noted for P.mosaica. This species is characterized by large rounded plates on the body wall and arched pillared tables on its tube feet.
The present results suggest that the taxonomy of Pseudothyone requires further investigation. Particularly, the generic affiliation of P.levini, P.belli and P.mosaica may require additional evaluation. The taxonomic position of the genus also remains unclear. Based on COI data, the raphanus clade (P.raphanus + P.serrifera + P.labradorensis sp. nov.) does not form any well-supported clade with other examined representatives of Sclerodactylidae sensu Smirnov (2012). More data, both morphological and molecular, are needed to analyze the phylogenetic relationships of Pseudothyone.
Scientific collections of fauna in the Labrador Sea are extremely limited so the distribution of this species is yet to be resolved. Pseudothyonelabradorensis sp. nov. is known from a single locality in the Labrador Sea, at a depth between 740–969 m. Metabarcoding surveys of the ISECOLD transects in the Labrador Sea (Côté et al. 2023) did not detect this species but several other holothuroids were detected over benthic habitats at a depth of 1026 m about 300 km to the north (Côté unpubl. data): Chiridotalaevis (O. Fabricius, 1780), Benthogone sp., Enypniastes sp., Psolidae gen. sp., and two unresolved species from the orders Dendrochirotida and Apodida.
Apart from P.labradorensis sp. nov., three more species of Pseudothyone are known from bathyal depths. The type species P.raphanus and another Northeast Atlantic species, P.serrifera, were reported down to 1200 m (Fernández-Rodríguez et al. 2019), with most of their records obtained shallower than 250 m and 600 m, respectively. The deepest species of the genus, P.furnestini, occurred between 440 and 1347 m (ibid.). According to McKenzie (1991), the body colour in P.raphanus can be “sometimes a pale pink”. Combined with personal unpublished observations of pinkish specimens of P.cf.raphanus off south Iceland, it is possible that P.labradorensis sp. nov. may have a wider distribution in the North Atlantic.
Key to the North Atlantic Pseudothyone
**: **
Supplementary Material
XML Treatment for Pseudothyone labradorensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arndt A Marquez C Lambert P Smith MJ (1996) Molecular phylogeny of eastern Pacific sea cucumbers (Echinodermata: Holothuroidea) based on mitochondrial DNA sequence.Molecular Phylogenetics and Evolution 6(3): 425–437. 10.1006/mpev.1996.00918975697 · doi ↗ · pubmed ↗
- 2Barrois T (1882) Catalogue des crustacés Podophtalmaires et des échinodermes recueillis à Concarneau durant les mois d'août septembre 1880. Lille, Imprimerie L Danel, 1–68 pp. 10.5962/bhl.title.53806 · doi ↗
- 3Cherbonnier G (1958) Note sur Pseudothyonesculponea nouvelles espèce d'holothurie dendrochirote de Méditerranée.Vie et Milieu 9: 62–66.
- 4Cherbonnier G (1969) Échinodermes récoltés par la Thalassa au large des côtes d'Espagne et du golfe de Gascogne. Bulletin du Muséum D'Histoire Naturelle, 2 série 41: 343–361.
- 5Clark HL (1901) The holothurians of the Pacific Coast of North America.Zoologischer Anzeiger 24: 162–171.
- 6CôtéD Heggland K Roul S Robertson G Fifield D Wareham V Colbourne E Maillet G Devine B Pilgrim L Pretty C Le Corre N Lawson JW Fuentes-Yaco C Mercier A (2019) Overview of the biophysical and ecological components of the Labrador Sea Frontier Area.DFO Canadian Science Advisory Secretariat Research Document 2018(067): 1–59.
- 7CôtéD Mc Clenaghan B Desforges J Fahner NA Hajibabaei M Chawarski J Roul S Singer G Aubry C Geoffroy M (2023) Comparing e DNA metabarcoding and conventional pelagic netting to inform biodiversity monitoring in deep ocean environments. In: Miller-Saunders K (Ed.) ICES Journal of Marine Science 80: 2545–2562. 10.1093/icesjms/fsad 169 · doi ↗
- 8Deichmann E (1954) The holothurians of the Gulf of Mexico. Bulletin of the U.S.Fish Commission 55: 381–410.