Conversion of polyploid and alloploid Saccharomyces sensu stricto strains to leu2 mutants by genome DNA editing
Kazuya Kiyokawa, Tetsushi Sakuma, Kazuki Moriguchi, Minetaka Sugiyama, Takeshi Akao, Takashi Yamamoto, Katsunori Suzuki

TL;DR
Researchers developed a CRISPR method to convert various yeast strains into leu2 mutants, enabling the use of existing plasmid resources for genetic studies.
Contribution
A modified CRISPR-Cas9 plasmid was created to generate leu2 mutants in diverse Saccharomyces sensu stricto strains.
Findings
CRISPR-Cas9 plasmids successfully targeted the LEU2 gene in multiple yeast species.
Leu2 mutants were generated in natural, industrial, and allopolyploid strains.
Most plasmids and mutants will be deposited in repositories for future use.
Abstract
A large number of recombinant plasmids for the yeast Saccharomyces cerevisiae have been constructed and accumulated over the past four decades. It is desirable to apply the recombinant plasmid resources to Saccharomyces sensu stricto species group, which contains an increasing number of natural isolate and industrial strains. The application to the group encounters a difficulty. Natural isolates and industrial strains are exclusively prototrophic and polyploid, whereas direct application of most conventional plasmid resources imposes a prerequisite in host yeast strains of an auxotrophic mutation (i.e., leu2) that is rescued by a selection gene (e.g., LEU2) on the recombinant plasmids. To solve the difficulty, we aimed to generate leu2 mutants from yeast strains belonging to the yeast Saccharomyces sensu stricto species group by DNA editing. First, we modified an all-in-one type…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
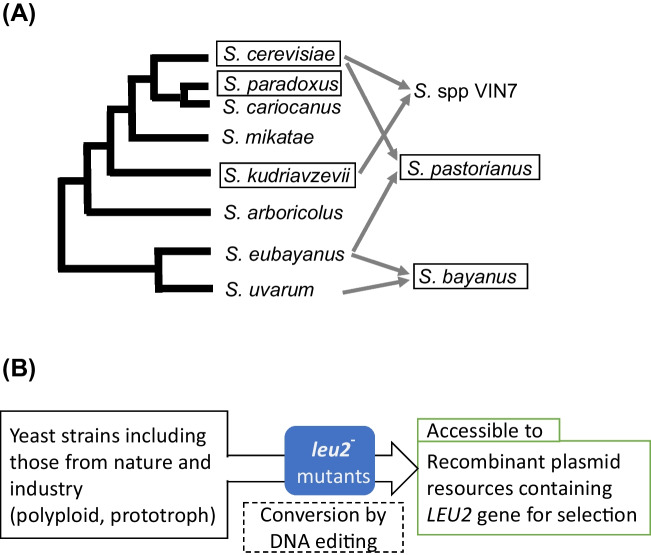 Figure 1
Figure 1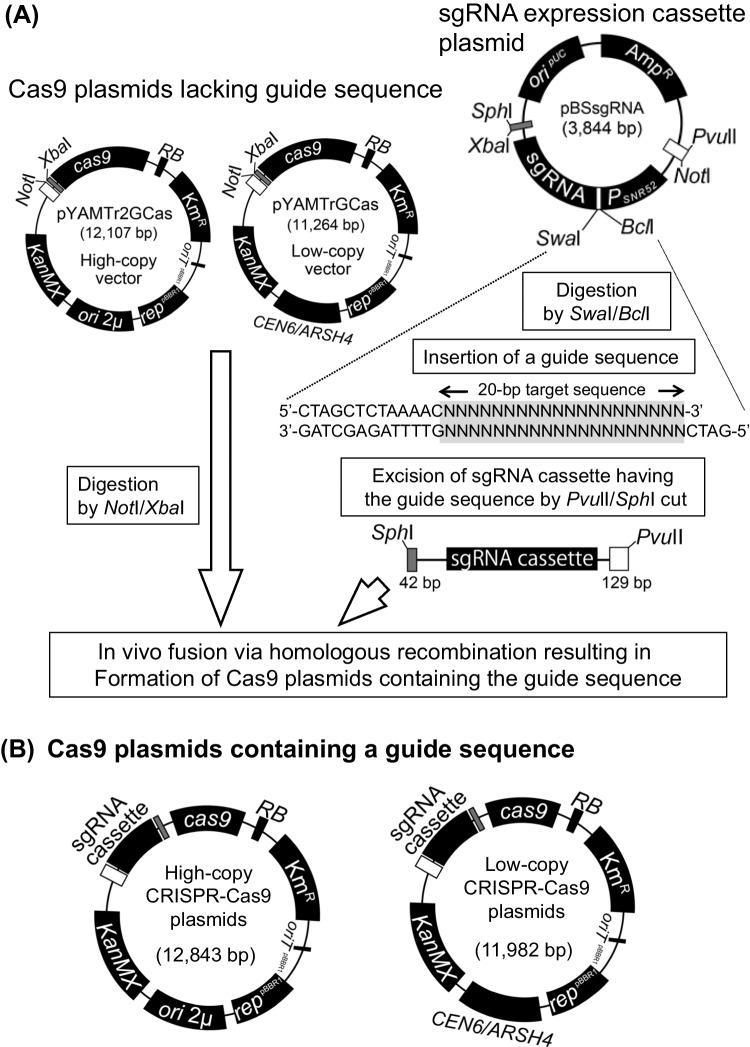 Figure 2
Figure 2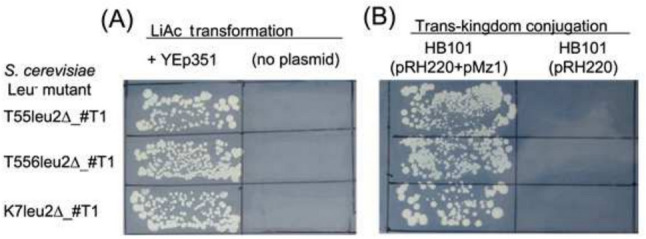 Figure 3
Figure 3 Figure 4
Figure 4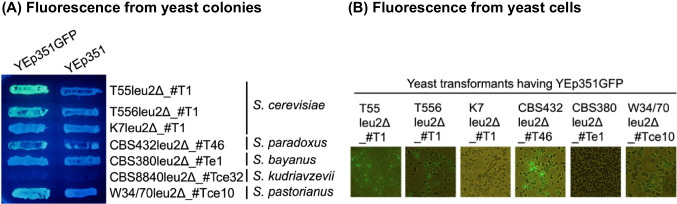 Figure 5
Figure 5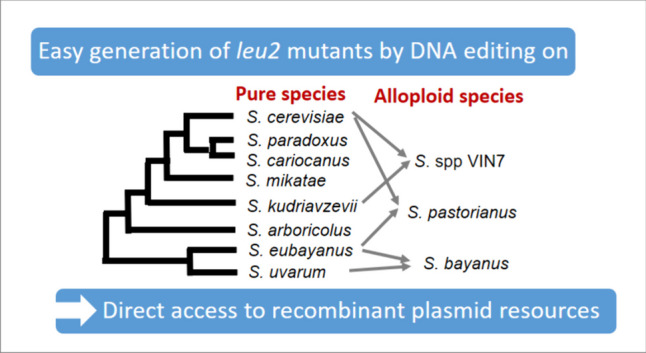 Figure 6
Figure 6- —Ministry of Education, Sports and Culture, Japan
- —Takaki Syunsuke Foundation for Science and Technology for Bread
- —http://dx.doi.org/10.13039/501100001691Japan Society for the Promotion of Science
- —http://dx.doi.org/10.13039/501100009035Program on Open Innovation Platform with Enterprises, Research Institute and Academia
- —Hiroshima University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal and yeast genetics research · CRISPR and Genetic Engineering · Fermentation and Sensory Analysis
Introduction
The yeast Saccharomyces sensu stricto complex group comprises eight “pure” species (e.g., S. cerevisiae, S. paradoxus, S. kudriavzevii, S. eubayanus) and interspecific hybrid species such as S. bayanus and S. pastorianus (Borneman and Pretorius 2015), as schematically shown in Fig. 1A. A large number of strains belonging to the yeast group play important roles in the production of foods and beverages in addition to other biotechnological applications (Replansky et al. 2008; Sicard and Legras 2011; Steensels et al. 2014). This number is increasing owing to the employment of new isolates from nature (Hisatomi and Toyomura 2021; Minnaar and den Haan 2023). The species in the group indicate intimate relationships with each other, that is, haploid cells can mate with the opposite mating-type cells belonging to the other species in the group, and the resulting hybrid cells are viable (Toyomura and Hisatomi 2021). However, the yeast group exhibits a large variation in their genomic sequences between species. Synonymous codon similarity % of S. mikatae, S. kudriavzevii, and S. bayanus to S. cerevisiae is as low as 55% on average (Cliften et al. 2003), which could ensure the durability of the yeast group in nature and generation of valiant strains for industry.Fig. 1. Simple way to convert Saccharomyces sensu stricto strains accessible to conventional recombinant plasmid resources. A Phylogenetic relationships between Saccharomyces sensu stricto group species. A phylogram drawn by Borneman and Pretorius (2015) was adopted with simplification. “Pure” species names are located at the center, whereas alloploid species names are shown on the right side. Allow marks show from which species the ancestor(s) of an alloploid species obtained genomes. Rectangles highlight the species employed for DNA editing experiments in this study. B Aims of this study
Since the reliable yeast transformation method was developed (Beggs 1978; Hinnen et al. 1978), every gene of the whole S. cerevisiae genome was cloned into various Escherichia coli-yeast shuttle vectors and integrative vectors for laboratory strains. For instance, Makanae et al. (2013) developed a series of overexpression plasmids, each of which carries a different protein-coding gene from S. cerevisiae. Various recombinant plasmid resources have been deposited in biological resource centers, and the plasmids are distributed on demand. Therefore, it is desirable to apply these plasmid resources to industrial and natural yeast strains for molecular breeding.
Most recombinant plasmids prepared for S. cerevisiae contain one of several nutrient synthesis genes (e.g., LEU2, URA3, and TRP1) as the transformant selection marker gene. The corresponding auxotrophic laboratory mutant strains (e.g., leu2, ura3, and trp1) are chosen as the host strain (Akada 2002; Gnugge and Rudolf 2017; Sikorski and Hieter 1989). However, industrial and natural isolate strains generally lack any auxotrophic mutation (prototrophic yeasts) and are mostly diploid and allopolyploid. Therefore, it is difficult to develop auxotrophic mutants suitable for transformation with the conventional recombinant plasmids in such strains. It is possible to add an antibiotic-resistance gene for yeast to the plasmids, but the modification of many substrate plasmids is laborious and time-consuming.
Genome editing of host cell chromosomes is an attractive solution method (Jinek et al. 2012; Raschmanova et al. 2018; Sakuma and Woltjen 2014; Yang and Blenner 2020) to overcome the difficulties in industrial and natural isolate yeast strains. As a convenient and accurate genome editing methodology, the CRISPR-Cas9 system is popular now. A short target sequence (20 nucleotides) in a single-guide RNA (sgRNA) is sufficient for the CRISPR-Cas9 system to generate a double-strand break (DSB) at the target sequence followed by a protospacer adjacent motif (PAM) (Jinek et al. 2012; Ran et al. 2013). In S. cerevisiae strains, the application of CRISPR-Cas9 plasmids containing an sgRNA gene results in the formation of mutations at the target locus. Furthermore, a template DNA that covers the target sequence but has some mutations improves the mutant ratio in DNA editing experiments (DiCarlo et al. 2013; Laughery et al. 2015; Raschmanova et al. 2018). Zhang et al. (2014) reported a Cas9 plasmid harboring a neurothricin-resistance gene and a series of sgRNA-expressing plasmids to target URA3, TRP1, LEU2, and HIS3, respectively. Successive transformation of an industrial S. cerevisiae strain with the Cas9 plasmid and then with one of the sgRNA-expressing plasmids resulted in formation of auxotrophic mutants depending on the guide RNA sequence. Laughery et al. (2015) constructed Cas9 plasmids (pML104 and pML107) containing an sgRNA-expression cassette gene as well as the selection marker gene LEU2 and URA3, respectively. Plasmids simultaneously containing Cas9 and sgRNA genes are categorized as all-in-one type, which is convenient because a single transformation produces mutants.
We aimed to broaden the applicability of the plasmid resources for Saccharomyces sensu stricto complex group. In this study, we focused on LEU2 gene disruption for several reasons. First, LEU2 has been frequently employed as the selection marker gene in yeast plasmid vectors in conventional plasmid resources. Second, among the auxotrophic mutations apt to the conventional yeast vectors, leu2 mutants are the most difficult to achieve in polyploid strains by conventional methods because there is no counter-selective characteristic against chemicals, whereas ura3 mutants are resistant to 5-fluoroorotic acid and trp1 mutants endure the killing action of 5-fluoroanthranilic acid (Akada 2002; Gnugge and Rudolf 2017; Kitamoto et al. 1990). Here, we show all-in-one type CRISPR-Cas9 plasmids with LEU2-gene-targeting guide sequences for application as widely as possible to Saccharomyces sensu stricto species. Dissemination of the Cas9 plasmids to yeast strains belonging to five species in the group produced leu2 mutants at practically desirable frequencies.
Materials and methods
Microbial strains, plasmids, and culture conditions
Table 1 lists bacterial and yeast strains and plasmids used in this study. Table 1. Bacterial and yeast strains and plasmids used in this study(A) Microbial strainsStrainRelevant genotype and/or characteristicsReference or sourceSaccharomyces cerevisiaeT55Laboratory strain, MATa prototrophTohoyama et al. 1979 (BY29297/NBRP)^1^T55leu2Δleu2(547–548, 551) mutant derived from T55This study (BY29298/NBRP)^1^T556Laboratory strain, MATa/MATa, prototrophCross between T55 and T56, Tohoyama et al. 1979 (BY29299/NBRP)^1^T556leu2Δleu2(547–548, 551) mutant derived from T556This study (BY29300/NBRP)^1^Kyokai No. 7 (K7)Rice wine (Sake)-brewing yeast, prototrophRIB1003/NRIB^2^K7leu2Δleu2(547–548, 551) mutant derived from Kyokai No. 7This study (NRIB)^3^IS289-1CMATa leu2-1 met8-1 aro1B ade8 cdc9Lab stockHD119-3MATa trp5 leu1 cyh2Lab stockS. paradoxusCBS432^T^ (IFO10609^T^)Type strain, an isolate from Quercus spp., prototrophBY20589/NBRP^1^CBS432leu2Δleu2(547–548, 551) mutant derived from CBS432^T^This study (BY29301/NBRP)^1^S. kudriavzeviiCBS8840^T^ (IFO1802^T^)Type strain, an isolate from partially decayed leaf in Japan, prototrophBY20109/NBRP^1^CBS8840leu2Δleu2(245–248) mutant derived from CBS8840^T^This study (BY29302/NBRP)^1^S. bayanusCBS380^T^ (IFO1127^T^)Type strain, an isolate from turbid beer, prototrophBY21388/NBRP^1^CBS380leu2Δleu2(547–548, 551) mutant derived from CBS380^T^This study (BY29303/NBRP)^1^S. pastorianusW34/70Lager beer-brewing yeast, prototrophHefebank Weihenstephan GmbH, GermanyW34/70leu2Δleu2(245–248) mutant derived from W34/70This study (NRIB)^3^Escherichia coliJM110dam dcm supE44 hsdR17 thi leu rpsL1 lacY galK galT ara tonA thr tsx**Δ(lac-proAB)/F'[traD36, proAB^+^, lacI^q^, lacZΔM15]Yanisch-Perron et al. 1985HB101mcrB mrr hsdS20(rB- mB-) recA13 leuB6 ara-14 proA2 lacY1 galK2 xyl-5 mtl-1 glnV44 λ^−^, Sm^R^Boyer and Roulland-Dussoix 1969HB101 RedETmcrB mrr hsdS20(rB- mB-) recA13 leuB6 ara-14 proA2 lacY1 galK2 xyl-5 mtl-1 glnV44 λ^−^ (repA^pSC101^* ori*^pSC101^ araC PBAD::gbarecA), Sm^R^ Ap^R^Lab stock(B) PlasmidsPlasmidRelevant genotype and/or characteristicsReference or sourceSource plasmids to prepare new Cas9 plasmids, a mini plasmid to insert a guide sequence, and a template DNApML104ori^pUC^ ScURA3 ori_2μ PTDH3::SpCas9 PSNR52::sgRNA cassette, Ap^R^Laughery et al. 2015 (#67638/Addgene)^3^pYAMTr2Grep^pBBR1^ RB oriT^pBBR1^ lacZα KanMX ori_2μ, Km^R^Kiyokawa et al. 2023 (BYP10212 /NBRP)^2^pYAMTrGrep^pBBR1^ RB oriT^pBBR1^ lacZα KanMX ARSH4 CEN6, Km^R^Kiyokawa et al. 2023 (BYP9801 /NBRP)^2^pYAMTrAurep^pBBR1^ RB oriT^pBBR1^ lacZα AurMX ARSH4 CEN6, Km^R^Kiyokawa et al. 2023 (BYP9799 /NBRP)^2^pBluescript KS ( +)ori^pUC^* lacZα*, Ap^R^Short et al. 1988Cas9 plasmids and a plasmid to insert guide sequencepYAMTr2GCasrep^pBBR1^ RB oriT^pBBR1^ lacZα KanMX ori_2μ PTDH3::SpCas9, Km^R^This study (BYP10221/NBRP)^1^pYAMTrGCasrep^pBBR1^ RB oriT^pBBR1^ lacZα KanMX ARSH4 CEN6 PTDH3::SpCas9, Km^R^This study (BYP10222/NBRP)^1^pBSsgRNAori^pUC^* P_SNR52_::sgRNA cassette, Ap^R^This study (BYP10223/NBRP)^1^Mini plasmids harboring a guide sequencepBSsgScLEU2ori*^pUC^* P_SNR52_::sgRNA cassette with ScLEU2 guide sequence, Ap^R^This studypBSsgSeLEU2ori*^pUC^* P_SNR52_::sgRNA cassette with SeLEU2 guide sequence, Ap^R^This studypBSsgSc-SeLEU2ori*^pUC^* P_SNR52_::sgRNA cassette with Sc-SeLEU2 guide sequence, Ap^R^This studyCas9 plasmids harboring a guide sequencepYAMTr2GCsgScLEU2pYAMTr2GC having the guide sequence ScLEU2, Km^R^This study (BYP10224/NBRP)^1^pYAMTrGCsgScLEU2pYAMTr2GC having the guide sequence ScLEU2, Km^R^This study (BYP10225/NBRP)^1^pYAMTr2GCsgSeLEU2pYAMTr2GC having the guide sequence SeLEU2, Km^R^This study (BYP10226/NBRP)^1^pYAMTr2GCsgSc-SeLEU2pYAMTr2GC having the guide sequence Sc-SeLEU2, Km^R^This study (BYP10227/NBRP)^1^Bacterial-yeast shuttle plasmids having ScLEU2 to select yeast transformants by Leu^−^ complementationYEp351ori*^pUC^ ScLEU2 ori_2μ, Ap^R^Hill et al. 1986YEp351GFPYEp351 having PGAL1::GFP, Ap^R^This studypFA6a-kanMX6-PGAL1-GFPori^pUC^ KanMx PGAL1::GFP, Ap^R^Longtine et al. 1998Plasmids for trans-kingdom conjugationpMz1ori^incQ^ ScLEU2 ori_2μ mob^incQ^ oriT^incQ^, Ap^R^ Gm^R^Mizuta et al. 2012pRH220ori^pUC^ oriT^RK2^, tra^RK2^ Cm^R^Mizuta et al. 2012^1^Serial number in National Bioresource Project (NBRP) budding yeast, Hiroshima, Japan^2^Serial number at National Research Institute of Brewing (NRIB), Hiroshima, Japan^3^Deposition communicates with NRIBDeposition communicates with NRIB and Addgene (Watertown, Massachusetts, USA)
Yeast strains were cultured in YPD medium (1% yeast extract, 2% polypeptone, 2% glucose) at 28 °C. E. coli strains were grown in LB medium (1% Bacto tryptone, 0.5% NaCl, 0.5% Bacto yeast extract) at 37 °C.
Yeast strains were cultured also on synthetic media. Synthetic dextrose (SD) medium consists of 2% glucose and 0.67% Bacto yeast nitrogen base without amino acids. Solid media for yeast were prepared by adding 2% agar. Synthetic galactose solid medium is the same with the solid SD medium but contains galactose in place of glucose. To verify auxotrophic mutant phenotypes in recombinant yeasts, colonies were streaked onto solid SD medium and solid SD medium supplemented with 30 μg/ml leucine.
Plasmid construction
An outline of the plasmid structure is shown in Fig. 2.Fig. 2. Scheme for preparing CRISPR-Cas9 plasmids harboring a 20-bp guide sequence. A Three tool plasmids and processes to construct all-in-one CRISPR-Cas9 plasmids containing a 20-bp guide sequence. The high-copy (ori 2μ) yeast vector pYAMTr2GCas and the low-copy (CEN/ARS) yeast vector pYAMTrGCas lack a guide sequence. The two yeast vectors were linearized by treatment with XbaI and NotI to accept the guide sgRNA expression cassette. The mini-plasmid pBSsgRNA possesses an sgRNA expression cassette lacking a guide sequence. A ds-oligonucleotide DNA containing a 20-bp guide sequence was ligated with BclI/SwaI-digested pBSsgRNA plasmid to place the target guide sequence on the cassette sgRNA expression cassette. The sgRNA expression cassette with the target guide sequence was excised from the guide-sequence-containing mini-plasmids by digestion with PvuII and SphI. Finally, the XbaI/NotI-linearized vector plasmids and the PvuII/SphI fragment harboring the sgRNA expression cassette were introduced into either a yeast strain or an E. coli strain expressing λ Red recombinase. The vector and guide-containing cassette DNAs were fused together in vivo by homologous recombination at a 42-bp overlapping region (left arm) and a 129-bp overlapping region (right arm). B Resultant CRISPR-Cas9 plasmids containing the target guide sequence
To build all-in-one type CRISPR-Cas9 plasmids containing the Cas9 gene and a sgRNA expression cassette with a 20-bp guide sequence, we first constructed two vector plasmids as follows. The Cas9 gene and a part of the target-lacking sgRNA expression cassette were excised from pML104 (Laughery et al. 2015) as a 5.1-kbp EcoRI/XbaI DNA fragment. The 5.1-kbp EcoRI/XbaI DNA fragment was ligated with a high-copy-type yeast Agrobacterium-mediated transformation (AMT) vector pYAMTr2G (Kiyokawa et al. 2023) cleaved by EcoRI and XbaI (7.0-kbp) and a low-copy-type yeast AMT vector pYAMTrG (Kiyokawa et al. 2023) linearized by EcoRI and XbaI (6.1-kbp), respectively. The resulting plasmids were named pYAMTr2GCas and pYAMTrGCas, respectively (Fig. 2A).
Second, a mini plasmid harboring the sgRNA expression cassette lacking a target sequence, which was obtained from pML104, was prepared as follows. The target-lacking sgRNA expression cassette was obtained as a 0.9-kbp PvuII/XhoI DNA fragment from pML104. The 0.9-kbp PvuII/XhoI DNA containing the target-lacking sgRNA expression cassette was ligated with EcoRV/XhoI-cleaved pBluescript KS ( +) plasmid DNA (Short et al. 1988) to form a 3.9-kbp plasmid named pBSsgRNA (Fig. 2A).
Three 20-bp guide sequences were designed as described in “Guide sequence for CRISPR-Cas9-mediated genome editing.”
Each ds-oligonucleotide guide sequence was inserted into the sgRNA expression cassette of the pBSsgRNA plasmid as follows: The pBSsgRNA plasmid DNA was prepared using a dam^−^ dcm^−^ E. coli strain JM110 (Yanisch-Perron et al. 1985). The pBSsgRNA plasmid was digested with SwaI overnight at 25 °C and subsequently treated with BclI for 4 h at 50 °C. Each of the three sets of complementary oligonucleotide DNA (Table 3 (A1)) containing the 20-bp guide sequence (Table 2 (A)) was ligated with the 3.8-kbp SwaI/BclI-digested pBSsgRNA. The resulting three plasmids were named pBSsgScLEU2, pBSsgSeLEU2, and pBSsgSc-SeLEU2, respectively.
Insertion of the full-length sgRNA expression cassette from each of the three pBSsgRNA derivatives into the two Cas9 plasmids was performed by in vivo fusion as follows. The Cas9-containing plasmids YAMTr2GCas and pYAMTrGCas were linearized by treatment with XbaI and NotI. The full-length sgRNA expression cassette with a 20-bp guide sequence was taken as a 0.9-kbp PvuII/SphI DNA fragment from pBSsgScLEU2, pBSsgSeLEU2, and pBSsgSc-SeLEU2, respectively. The linearized 12.1-kbp YAMTr2GCas and the 0.9-kbp PvuII/SphI fragment possessing the sgRNA expression cassette were simultaneously introduced into an E. coli strain HB101 RedET expressing λ phage recombinase genes γ, β, and α. In the E. coli cells, the two DNAs were fused in vivo by homologous recombination at 129-bp and 42-bp overlapping sequences between each end of the two DNAs. The three resulting high-copy-type CRISPR-Cas9 plasmids were named pYAMTr2GCsgScLEU2, pYAMTr2GCsgSeLEU2, and pYAMTr2GCsgSc-SeLEU2, respectively.
Construction of low-copy type all-in-one Cas9 plasmids using pYAMTrGCas was performed as described above, but the in vivo fusion in E. coli was replaced with that in yeast strain T55. The resulting low-copy-type CRISPR-Cas9 plasmids were named pYAMTrGCsgScLEU2, pYAMTrGCsgSeLEU2, and pYAMTrGCsgSc-SeLEU2, respectively.
YEp351_PgalGFP was prepared by ligation between SalI- and BamHI-digested YEp351 and 1.5 kbp fragment from SalI- and BglII-digested pFA6a-kanMX6-PGAL1-GFP.
Guide sequence for CRISPR-Cas9-mediated genome editing
We selected 20-bp guide sequences in the target LEU2 gene as follows: Genome sequences of 50 S. cerevisiae strains were retrieved from the Saccharomyces genome database ((Cherry et al. 2012); https://www.yeastgenome.org). To identify common sequences among 50 S. cerevisiae strains in each target gene, multiple sequence alignment was performed using the MAFFT program version 7.463 (Katoh and Standley 2013). Target sequence candidates with low off-target effects were searched using CRISPRdirect (Naito et al. 2015) and CRISPOR (Concordet and Haeussler 2018) programs. Highly conserved regions in the LEU2 gene were used as the query sequence. We examined the applicability of the target sequence candidates to Saccharomyces sensu stricto species using the nucleotide blast program and chose guide sequences (Table 2) to make the applicability as wide as possible.Table 2. Target sequence in LEU2 gene for CRISPR-Cas9(A) Guide sequences designed Name20-bp Guide sequence + PAMPosition in ORFCas9 sgRNA expression plasmid harboring each guide sequence ScLEU2AAGGACCAAATAGGCAATGGTGG569–547pYAMTrGCScLEU2pYAMTr2GCScLEU2 SeLEU2AAGGACCAGATAGGTAATGGTGG569–547pYAMTr2GCSeLEU2 Sc-SeLEU2TGCTGTGGGTGGTCCTAAATGGG225–247pYAMTr2GCSc-SeLEU2(B) Sequence difference from the guide sequences Species, strainScLEU2 (SeLEU2) + PAMTypeSc-SeLEU2 + PAMGenBank assembly accession* S. cerevisiae* S288cAAGGACCAAATAGGCAATGGTGGScTGCTGTGGGTGGTCCTAAATGGGGCA_000146045.2* S. paradoxus* CBS432^T^AAGGACCAAATAGGCAATGGTGGScTGCTGTGGGTGGTCCTAAATGGGGCA_002079055.1* S. cariocanus* NCYC 2890^ T^(Data absent)(Data absent)(Data absent)* S. mikatae* IFO1815^T^AAGGACCAAATAGGCAATGGTGGScTGCAGTAGGTGGTCCTAAATGGGGCA_000167055.1* S. kudriavzevii* CBS8840^T^(Data absent)(Data absent)GCA_00167075.2* S. arboricolus* H-6^ T^AAGGACCAAATAGGCAATGGTGGScTGCTGTAGGTGGTCCTAAATGGGGCA_000292725.1* S. eubayanus* CBS12357^T^AAGGACCAGATAGGTAATGGTGGSeTGCTGTGGGTGGTCCTAAATGGGGCA_001515405.2* S. uvarum* CBS7001AAGGACCAGATAGGTAATGGTGGSeTGCTGTGGGTGGTCCTAAATGGGGCA_027557585.1* S. pastorianus* CBS1513^T^AAGGACCAAATAGGCAATGGTGGScTGCTGTGGGTGGTCCTAAATGGGGCA_013180785.1 (scaffold1263_cov98)AAGGACCAGATAGGTAATGGTGGSeTGCTGTGGGTGGTCCTAAATGGGGCA_013180785.1 (scaffold184_cov57)* S. bayanus* CBS380^T^AAGGACCAGATAGGTAATGGTGGSeTGCTGTGGGTGGTCCTAAATGGGGCA_013180675.1A bold underlined face letter indicates a nucleotide sequence different from that of S. cerevisiae
DNA transformation in yeast
The lithium acetate (LiAc) yeast transformation method was performed according to a protocol described by Gietz and Schiestl (2007) with minor modifications. Yeast cells (1.9 × 10^7^ colony forming unit (CFU)) were suspended in a solution containing 100 ng of circular plasmid DNA, LiAc, polyethylene glycol (PEG) 4000, and carrier DNA and incubated for 40 min at 42 °C.
Before selection using G418, the yeast cells were then cultured in liquid YPD medium at 28 °C for 2 h (for experiments using laboratory strains only) or 3.5 h. After the cultivation, yeast cells were collected and spread on solid YPD medium containing 400 μg/ml G418. To enhance the mutant frequency, 50 pmol of a hybridized 90-base oligonucleotide DNA pair (which were named as dsOligo DNAs as listed in Table 3 (B)) was added as the template DNA together with the CRISPR-Cas9 plasmid DNA to the LiAc/PEG 4000/carrier DNA solution.Table 3. List of oligonucleotides used in this study(A) Oligonucleotides for preparation of guide sequences and of template DNAs Oligonucleotide nameSequence (5’-3’)Application(A1) Complementary pairs for preparation of a guide sequenceResultant plasmid ScLEU2sgRNA_FwGATCAAGGACCAAATAGGCAATGGGTTTTAGAGCTAGpBSsgScLEU2 ScLEU2sgRNA_RvCTAGCTCTAAAACCCATTGCCTATTTGGTCCTT SeLEU2sgRNA_FwGATCAAGGACCAGATAGGTAATGGGTTTTAGAGCTAGpBSsgSeLEU2 SeLEU2sgRNA_RvCTAGCTCTAAAACCCATTACCTATCTGGTCCTT Sc-SeLEU2sgRNA_FwGATCTGCTGTGGGTGGTCCTAAATGTTTTAGAGCTAGpBSsgSc-SeLEU2 Sc-SeLEU2sgRNA_RvCTAGCTCTAAAACATTTAGGACCACCCACAGCA(A2) Complementary pairs for preparation of template DNAsResultant template DNA D-ScLEU2_FwAGAATCACAAGAATGGCCGCTTTCATGGCCCTACAACATGAGACTATTGCCTATTTGGTCCTTGGATAAAGCTAATGTTTTGGCCTCTTCdsOligo DNA (ScLEU2) D-ScLEU2_RvGAAGAGGCCAAAACATTAGCTTTATCCAAGGACCAAATAGGCAATAGTCTCATGTTGTAGGGCCATGAAAGCGGCCATTCTTGTGATTCT D-SeLEU2_FwAGAATTACAAGAATGGCTGCGTTCATGGCACTACAACACCAAACTATTACCTATCTGGTCCTTGGATAAAGCCAATGTTTTGGCCTCATCdsOligo DNA (SeLEU2) D-SeLEU2_RvGATGAGGCCAAAACATTGGCTTTATCCAAGGACCAGATAGGTAATAGTTTGGTGTTGTAGTGCCATGAACGCAGCCATTCTTGTAATTCT D-Sc-SeLEU2_FwAGAAGGCTGATGCCGTTTTGTTAGGTGCTGTGGGTGGTCCTAAATTACCGGTAGTGTTAGACCTGAACAAGGTTTACTAAAAATCCGTAAdsOligo DNA (Sc-SeLEU2) D-Sc-SeLEU2_RvTTACGGATTTTTAGTAAACCTTGTTCAGGTCTAACACTACCGGTAATTTAGGACCACCCACAGCACCTAACAAAACGGCATCAGCCTTCT(B) Primers for PCR to prepare sequence template and for sequencing Oligonucleotide nameSequence (5′-3′)PCR and sequence targetYeast species F-PCR_LEU2-UP_FwGTACCGGTAGTGTTAGACScLEU2**S. cerevisiae, S. paradoxus F-PCR_LEU2-DO_RvGTTCGTACAAACCAAATGC Seu_LEU2_Fw2GGTCAAGAAATCACTGAGGSeLEU2**S. bayanus Seu_LEU2_Rv2CCAATGAACCAGGAATACAA SkuLEU2_FwGAAGCAATTAAGGTTCTSc-SeLEU2**S. kudriavzevii SkuLEU2_RvCTCCCACTAACTCTCTAAC F-PCR_LEU2-UP_Fw2^1^GAAGCCATTAAGGTTCTSc-SeLEU2**S. pastorianus Sc_LEU2_Rv2^1^AAGGAACCTGGGATAACG Seu_LEU2_Rv2^1^CCAATGAACCAGGAATACAA^1^F-PCR_LEU2-UP_Fw2 anneals with Sc-LEU2 target sequence and with Se-LEU2 target sequence, while Sc_LEU2_Rv2 and Se_LEU2_Rv2 specifically hybridizes with the targets ScLEU2 and SeLEU2, respectively
Transformation frequency was represented as the number of transformants per μg of plasmid DNA per output viable cell number after LiAc/PEG 4000 treatment.
Trans-kingdom conjugation to yeast
A mobilizable LEU2 plasmid pMz1was transferred from a donor E. coli strain HB101 that harbors pMz1 and a helper plasmid pRH220 as shown by Mizuta et al. (2012). After co-incubation between a leu2 mutant yeast and the donor E. coli cells, the cell mixture was spread onto SD solid medium, which allows selective growth of yeast cells that obtained pMz1.
Nucleotide sequence determination
The DNA sequencing was determined using the Sanger method by Eurofin Co. (Tokyo, Japan). Template DNA sequencing reaction was prepared by PCR amplification using primer sets listed in Table 3 (B). In leu2 mutants in aneuploid strains, each of two leu2 genes was amplified using allele-specific primer sets shown in Table 3 (B) and sequenced respectively.
Statistical analysis
Statistical analysis was performed using the R program version 4.0.2 and its expansion package (https://www.R-project.org/). Each datum in a table represents a set of a mean value ± its standard deviation (SD). Individual methods and experimental replicates for statistical comparisons are presented in each table.
Results
Construction of all-in-one CRISPR-Cas9 plasmids for conversion of prototrophic yeast strains to leu2 mutant strains
Figure 2 depicts a scheme to construct all-in-one CRISPR-Cas9 plasmids. Using a Cas9 expression gene and sgRNA expression gene cassette from pML104 (Laughery et al. 2015), we prepared three tool plasmids, namely, a small plasmid pBSsgRNA and two CRISPR-Cas9 vector plasmids (pYAMTr2GCas and pYAMTrGCas), which lacked the target sequence. A 20-bp guide sequence for Cas9 scission was inserted into the sgRNA expression gene cassette in pBSsgRNA, and the cassette with the guide sequence was then moved to one of the two CRISPR-Cas9 vector plasmids. The vector plasmid pYAMTr2GCas is a high-copy and pYAMTrGCas is a low-copy replication type, because the former contains ori_2μ and the latter harbors CEN/ARS (Sikorski and Hieter 1989). Because half of the two vectors were derived from pYAMTrG and pYAMTr2G (Kiyokawa et al. 2023), respectively, they harbored the G418 resistance gene cassette KanMX to enable transformant selection for prototrophic yeast strains and contained a right border sequence (RB), an overdrive sequence, and the pBBR1 replication gene, which as a whole permit mobilization by the AMT method.
Figure 1A illustrates the phylogenetic relationship of yeast species among Saccharomyces sensu stricto group. As listed in Table 2 (A), we designed three 20-bp guide sequences for DNA editing at LEU2 gene, aiming as broadly as possible among Saccharomyces sensu stricto species strains. In DNA databases, LEU2 gene sequence was available for eight species in the group (Table 2 (B)). The three guide sequences matched well with the sequences in the database and were equipped to the two Cas9 vector plasmids, as listed in Table 2 (A).
In this study, we used seven strains belonging to five yeast species. Most of these strains are natural and industrial strains. As a pilot application with a pedestal purpose, we examined the basic behavior of yeast toward the CRISPR-Cas9 plasmids and template DNAs using two S. cerevisiae laboratory strains.
High yield of leu2 mutant strains from S. cerevisiae laboratory strains
The two all-in-one type plasmids pYAMTrGCsgScLEU2 and pYAMTr2GCsgScLEU2 contain a 20-bp guide sequence for S. cerevisiae LEU2 gene (Table 2 (A)). The former plasmid is of the multi-copy type and the latter is of the low-copy type. Introduction of the two plasmids into the haploid laboratory strain T55 generated prototrophic (Leu^+^) colonies and leucine-auxotrophic (Leu^−^) colonies. As shown in Table 4, the ratio of Leu^−^ colonies was approximately 90%, irrespective of the plasmid copy difference. The diploid laboratory strain T556 produced fewer recombinant colonies than the haploid strain. The ratio of Leu^−^ colonies by the diploid strain was around 50%, which is a half the ratio of the haploid strain. Table 4LEU2 gene disruption in haploid and diploid S. cerevisiae laboratory strainsYeast strainTemplate DNA^1^G418^R^ recombinant colony numberTransformation efficiency (× 10^−4^)Leu^−^ mutant ratio^3^ (%)(A) pYAMTr2GCsgScLEU2 (high copy) HaploidT55dsOligo DNA (ScLEU2)3930 ± 177b^2^1.9 ± 0.6b^2^118/12098ー78 ± 11a0.04 ± 0.01a91/10190 DiploidT556dsOligo DNA (ScLEU2)1840 ± 856b1.2 ± 0.7b116/12097ー10 ± 4a0.005 ± 0.0006a9/2339(B) pYAMTrGCsgScLEU2 (low copy) HaploidT55dsOligo DNA (ScLEU2)301 ± 145b0.2 ± 0.1b155/15898ー16 ± 3a0.01 ± 0.002a50/5394 DiploidT556dsOligo DNA (ScLEU2)178 ± 78b0.1 ± 0.06b119/12694ー7 ± 5a0.004 ± 0.002a12/2060Haploid laboratory strain T55 and diploid laboratory strain T556 were transformed with pYAMTr2GCsgScLEU2 (A) and pYAMTrGCsgScLEU2 (B) using LiAc method. After LiAc/PEG treatment, the yeast cells were cultured in liquid YPD medium at 28 °C for 2 h^1^An annealed pair of oligonucleotide DNAs (dsOligo DNA), which fit the assumed double-strand-break end regions at the target in ScLEU2, was applied as the template DNA together with the all-in-one CRISPR-Cas9 plasmids to the yeast strains. See Table 3 (B)^2^Different letters indicate significant differences (P < 0.05) according to the Tukey–Kramer test (n = 3–4)^3^Ratio of leucine-auxotrophic colonies per G418^R^ resistant colonies
We prepared ds-oligonucleotide DNA (Sc), which is a hybridization of two complementary 90-base sequences that match the target region of ScLEU2 but lacks two nucleotides in the PAM sequence (Table 3 (A2)). Addition of the ds-oligo DNA markedly increased the number of recombinant colonies by more than tenfold, and the Leu^−^ ratio was up to 98% in the haploid strain and approximately 95% in the diploid strain (Table 4).
The mutation(s) that caused the Leu^−^ phenotype were located at the LEU2 locus. First, leucine auxotrophy of the Leu^−^ mutant recombinant colonies was rescued by LiAc transformation with YEp351 (Fig. 3A) and by trans-kingdom conjugation with a donor E. coli HB101 harboring a mobilizable ScLEU2 plasmid pMz1 and a helper plasmid pRH220 (Fig. 3B). Second, crossing haploid Leu^−^ colonies with an opposite mating-type leu2 mutant tester strain IS289-1C failed to complement the Leu^−^ phenotype, while crossing with an opposite mating-type leu1 mutant tester strain HD119-3 complemented the Leu^−^ phenotype (data not shown). Finally, DNA sequencing analysis verified that the mutation is present at the target sequence in LEU2 gene as shown in Fig. 4A. Three haploid and three diploid mutants had a sequence identical to the template ds-oligonucleotide DNA sequence.Fig. 3. Rescue of Leu^−^ phenotype by introduction of ScLEU2 gene. A LiAc transformation with YEp351, which harbors ScLEU2. B Trans-kingdom conjugation using a donor E. coli strain HB101 containing a helper plasmid pRH220 and a ScLEU2-containing mobilizable plasmid pMz1Fig. 4Verification of leu2 mutations by nucleotide sequence analysis. A template 90-bp oligonucleotide DNA sequence was aligned with the corresponding wild-type (WT) and mutant gene sequences. Shade boxes and underlines indicate the 20-bp guide sequence and PAM, respectively. Arrows and filled triangles indicate nucleotide polymorphisms among leu2 mutant strains and between wild-type and template DNA sequences, respectively. A Four mutants derived from S. cerevisiae strain T55 and four mutants from the diploid strain T556. B Three mutants of the sake-producing strain S. cerevisiae Kyokai No. 7 (K7). C Three mutants of S. paradoxus strain CBS432^T^. D Three mutants of S. bayanus strain CBS380^T^. E Two mutants from S. kudriavzevii strain CBS8840^T^. F Three mutants of the beer-producing S. pastorianus strain W34/70
Generation of leu2 mutants from a sake (rice-wine) brewing strain and a S. paradoxus strain
We applied the high-copy plasmid pYAMTr2sgScLEU2 and the template ds-oligonucleotide DNA (Sc) to five yeast strains, which consisted of industrial and natural isolate strains.
Sake-producing S. cerevisiae strain Kyokai No. 7 (K7) is a diploid (Akao et al. 2011). The strain showed a much lower Leu^−^ mutant ratio (19%) than the diploid laboratory strain T556 (100%). Conversely, K7 strain yielded a sevenfold larger number of G418^R^ recombinant colonies than T556 (Table 5). We examined three Leu^−^ mutants of the K7 strain using DNA sequencing analysis at the LEU2 locus. All three K7 mutants exhibited a sequence identical to the template ds-oligonucleotide DNA sequence (Sc) (Fig. 4B). Table 5. Leu^−^ mutant formation induced by CRISPR-Cas9 plasmids in yeasts belonging to five Saccharomyces sensu stricto speciesYeast strainCRISPR Cas9 plasmid having target sequence type^1^ in LEU2 geneTemplate DNA^2^G418^R^ recombinant colony numberTransformation efficiency (× 10^−5^)Leu^−^ mutant ratio (%)S. cerevisiae T556ScLEU2dsOligo DNA (ScLEU2)547 ± 1191.5 ± 0.340/40100SeLEU2dsOligo DNA (SeLEU2)(1.1 ± 0.3) × 10^4^38 ± 190/1200Sc-SeLEU2dsOligo DNA (Sc-SeLEU2)560 ^3^1.9 ^3^6/6 ^3^100 ^3^(No plasmid)(No DNA)Not detected––– Kyokai No. 7ScLEU2dsOligo DNA (ScLEU2)4063 ± 113722 ± 8.823/12019(No plasmid)(No DNA)Not detected − − − S. paradoxus CBS432^T^ScLEU2dsOligo DNA (ScLEU2)132 ± 460.8 ± 0.3112/12093(No plasmid)(No DNA)Not detected–––S. bayanus CBS380^T^ScLEU2dsOligo DNA (ScLEU2)751 ± 576.9 ± 1.10/1680SeLEU2dsOligo DNA (SeLEU2)111 ± 530.9 ± 0.4120/120100Sc-SeLEU2dsOligo DNA (Sc-SeLEU2)537 ^3^5.5 ^3^40/40 ^3^100 ^3^(No plasmid)(No DNA)Not detected–––S. pastorianus W34/70ScLEU2dsOligo DNA (ScLEU2)272 ± 1261.9 ± 0.80/1680SeLEU2dsOligo DNA (SeLEU2)303 ± 2282.9 ± 2.60/1200Sc-SeLEU2dsOligo DNA (Sc-SeLEU2)1106 ± 26211 ± 3.436/22416(No plasmid)(No DNA)Not detected–––S. kudriavzevii CBS8840^T^ScLEU2dsOligo DNA (ScLEU2)69 ± 501.9 ± 1.60/1170SeLEU2dsOligo DNA (SeLEU2)435 ± 33912 ± 110/1200Sc-SeLEU2dsOligo DNA (Sc-SeLEU2)435 ± 17410 ± 7.920/1209(No plasmid)(No DNA)Not detected–––Yeast strains were transformed with a combination of a CRISPR Cas9 plasmid and its corresponding template DNA: pYAMTr2GCsgScLEU2 plasmid and a template ds-oligonucleotide DNA (Sc); pYAMTr2GCsgSeLEU2 and ds-oligonucleotide DNA (Se); pYAMTr2GCsgSc-SeLEU2 and ds-oligonucleotide DNA (Sc-Se)To yield larger number of recombinant colonies from Saccharomyces sensu stricto yeasts, cultivation after LiAc/PEG treatment was extended to 3.5 h (see “Materials and methods”)^1^See Table 2 (A)^2^See Table 3 (B) and footnote #3 in Table 4^3^Data from one time experiment
S. paradoxus CBS432^T^ is a diploid strain. As shown in Fig. 2B, CBS432^T^ strain’s LEU2 gene has 20-bp guide and PAM sequences identical to those of the ScLEU2 gene, whereas there were single nucleotide polymorphisms (SNPs) near the target sequence in S. paradoxus LEU2 compared with ScLEU2 (data not shown). As indicated in Table 5, the CBS432^T^ strain showed a high Leu^−^ mutant ratio (93%). We analyzed the LEU2 sequence in three CBS432^T^ Leu^−^ mutant strains. All three mutants contained one nucleotide substitution in the target sequence and lacked PAM, which appears to reflect an event of replacement by double crossovers with the template ds-oligonucleotide DNA (Sc) (Fig. 4C). In contrast, the three CBS432^T^ leu2 mutant strains showed small differences from the template DNA in sequences surrounding the target sequence, which is likely to show crossover positions near the scission site between the template ds-oligonucleotide DNA and the chromosomal LEU2 gene in the S. paradoxus strain (Fig. 4C).
In contrast to the S cerevisiae and S. paradoxus strains, application of the same plasmid and the template ds-oligonucleotide DNA produced no auxotrophic mutant in S. kudriavzevii CBS8840^T^, S. bayanus CBS380^T^, and S. pastorianus W34/70 (Table 5). This result for S. bayanus and S. pastorianus coincides with the estimation based on their LEU2 gene sequences in the database (Table 2 (B)), whereas no data were available for S. kudriavzevii.
Formation of leu2 mutants from S. bayanus using a guide sequence suitable for S. eubayanus-type LEU2
As shown in Fig. 1A, S*. bayanus* CBS380^T^ is an allodiploid strain whose hybrid genome originated from S. eubayanus and S. uvarum, whereas S. pastorianus W34/70 is an allodiploid strain with an amalgam genome derived from S. eubayanus and S. cerevisiae (Borneman and Pretorius 2015; de la Cerda Garcia-Caro et al. 2022). The all-in-one CRISPR-Cas9 plasmid pYAMTr2GCsgSeLEU2 contains a guide sequence (SeLEU2) suitable for S. eubayanus LEU2 gene as the target.
The S. bayanus strain CBS380^T^ was transformed simultaneously with pYAMTr2GCsgSeLEU2 and a template ds-oligonucleotide DNA (Se), which is similar to the template ds-oligonucleotide DNA (Sc) but suits S. eubayanus LEU2 (Fig. 4D). As indicated in Table 5, the CBS380^T^ strain produced Leu^−^ colonies upon the transformation. Its Leu^−^ ratio was 100%. DNA sequencing analysis verified that three S. bayanus Leu^−^ mutants had a sequence identical to the ds-oligonucleotide DNA (Se) sequence in the target LEU2 locus (Fig. 4D).
Conversely to S. bayanus, and accordingly with the estimation shown in Table 2 (B), S. pastorianus, S. kudriavzevii, and S. cerevisiae strains failed to generate any Leu^−^ mutants by transformation with pYAMTr2GCsgSeLEU2 and its template (Table 5).
Conversion of S. pastorianus and S. kudriavzevii strains to leu2 mutants using a guide sequence common to S. cerevisiae-type and S. eubayanus-type LEU2
The all-in-one CRISPR-Cas9 plasmid pYAMTr2GCsgSc-SeLEU2 contains a 20-bp guide sequence (Sc-Se LEU2 in Table 2), which is highly conserved between Sc-type and Se-type LEU2.
The S. pastorianus strain W34/70 was transformed with a mixture of pYAMTr2GCsgSc-SeLEU2 and its corresponding template ds-oligonucleotide DNA (Sc-Se). The resulting recombinant W34/70 colonies exhibited a Leu^−^ mutant ratio 17% (Table 5). The same mixture was also applied to S. kudriavzevii strain CBS8840^T^as well as to S. cerevisiae T556 and S. bayanus CBS380^T^ strains. This treatment generated Leu^−^ colonies. The Leu^−^ ratio among recombinant colonies of CBS8840^T^ was 7%, whereas that of T556 and CBS380^T^ was 100% (Table 5).
DNA sequencing analyses revealed that all Leu^−^ mutants from W34/70 and CBS8840^T^ strains had a four-nucleotide deletion surrounding PAM, which is likely due to a double-crossover with the template DNA at the target LEU2 locus (Fig. 4E, F). Leu^−^ mutants from S. pastorianus strain W34/70 had some nucleotide polymorphisms, which were probably a consequence of homologous recombination between chromosomal Sc-type and Se-type LEU2 alleles at the target locus (Fig. 4F). One CBS8840^T^ Leu^−^ mutant strain (#Tce39) contained an additional four-nucleotide deletion in the target sequence (Fig. 4E). These results support our assumption that the pYAMTr2GCsgSc-SeLEU2 plasmid, which has the guide sequence Sc-SeLEU2 most common to both Sc-type and Se-type LEU2 genes, is applicable to a broad range of Saccharomyces sensu stricto yeasts to generate leu2 mutant strains.
Easy removal of all-in-one CRISPR-Cas9 plasmid from recombinant leu2 mutants
After DNA editing by plasmid transformation, the plasmid DNA needs to be removed from the resultant mutant transformants for subsequent studies. We analyzed the loss frequency of a resident CRISPR-Cas9 plasmid from recombinant leu2 mutants. First, we cultured leu2 mutant strains induced by pYAMTr2GCsgScLEU2 derived from S. cerevisiae T556 and S. paradoxus CBS432^T^ in liquid YPD medium (without G418) for 24 h and then established colonies on solid YPD medium. Sensitivity to G418 was examined for each colony. All 40 T556- and CBS432^T^-derived colonies were sensitive to G418, indicating that the resident CRISPR-Cas9 plasmid is easily removable.
One leu2 mutant from each of the seven strains was treated as described above. Consequently, G418-sennsitive colonies were obtained easily. PCR analysis further confirmed the absence of the Cas9 plasmid in a G418^S^ leu2 mutant colony of the respective strains. The leu2 mutant strains free of the Cas9 plasmid are available in two public depositories (Table 1 B).
Expression of a foreign gene by transformation of resulting leu2 mutants
The leu2 mutants were transformed with a LEU2 plasmid YEp351GFP, which contains a green fluorescent protein gene that is located just downstream of the promoter of ScGAL1. As shown in Fig. 5, leu2 mutants derived from S. cerevisiae T55 and T556, S. paradoxus CBS432^T^, S. pastorianus W34/70 emitted green fluorescence when transformed with YEp351GFP and grew on the galactose medium, while those with the vacant vector plasmid YEp351 did not indicate the fluorescence. Mutants from S. cerevisiae K7 and S. bayanus CBS380^T^ did not emit the fluorescence. The S. kudriavzevii mutants as well as S. kudriavzevii CBS8840^T^ did not grow on the galactose medium. These results suggest that the S. paradoxus and S. pastorianus strains possess a regulatory mechanism similar with that of the S. cerevisiae galactose regulon, while the S. cerevisiae K7 and S. bayanus CBS380^T^ strains have a different mechanism.Fig. 5. Introduction of a green fluorescent protein gene in yeast leu2 mutants. A Yeast leu2 mutants transformed with YEp351 and its derivative plasmid (YEp351GFP) containing a P_GAL1_::gfp gene were grown on synthetic galactose solid medium. Yeast colonies were irradiated with 365 nm LED light. B The transformed yeast cells harboring YEp351_P_GAL1_GFP were observed under blue light excitation
Applicability prediction of the all-in-one plasmids to other yeast species
In this study, we did not handle strains belonging to five “pure species,” namely, S. cariocanus, S. mikatae, S. arboricolus, S. eubayanus, and S. uvarum. However, it is easy to predict that S. eubayanus and S. uvarum are good substrates for LEU2 gene disruption by the plasmids containing the S. eubayanus type guide (Se), because 100% Leu^−^ ratio was exhibited (Table 5) by the allodiploid S. bayanus strain CBS380^T^, whose genome was derived from S. eubayanus and S. uvarum (Borneman and Pretorius 2015). Furthermore, the DNA database supplied LEU2 gene sequences for S. eubayanus and S. uvarum (Table 2 (B)). The sequence data indicated a perfect match to the guide sequence Se-SeLEU2 (Table 2 (B)). Similarly, the LEU2 sequences of S. mikatae and S. arboricolus suited exactly to the guide sequence SeLEU2. These data forecast the suitability of the Cas9 plasmids for the five remaining species.
Discussion
In this study, we prepared leu2 mutants from seven prototrophic yeast strains belonging to five species of the Saccharomyces sensu stricto group by DNA editing. Leu^−^ mutant ratios varied among the yeast strains. Three diploid strains, namely, one S. cerevisiae laboratory strain T556 and the type strains of S. paradoxus and S. bayanus, achieved an almost 100% Leu^−^ mutant ratio (Table 5). Contrary to the high efficacy of the three diploid strains, S. cerevisiae Kyokai No. 7 (K7), S. kudriavzevii CBS8840^T^, and S. pastorianus W34/70 exhibited lower Leu^−^ ratios, 9–19% (Table 5). Despite the unexpectedly low ratio level in the three strains, it was easy to find the target leu2 mutants because the discrimination between Leu^+^ and Leu^−^ was a facile work for the number of colonies, and all of the Leu^−^ colonies examined by sequence analysis were leu2 mutants (Fig. 4), irrespective of the ratio levels. The guide sequences in the Cas9 plasmids mostly fit the strains (Table 2 (B), Fig. 4). However, the template DNA sequences differed by several nucleotides in some strains (Fig. 4). Therefore, the ratio could be increased by supplying a template DNA that perfectly matches the sequence around the guide sequence for each strain. We discuss an additional scenario due to the presence of the case where the template DNA sequence was not the issue of the template DNA sequence.
Feasible reason for the cases of unexpectedly low Leu− mutant ratio
In the DNA databases, no LEU2 sequence was available for S. kudriavzevii CBS8840^T^, when we started this study. Sequence analysis of CBS8840^T^ strain’s LEU2 revealed one nucleotide difference at 5’ end region of the target 20 bp compared with the common guide sequence for Sc-SeLEU2 (Fig. 4E). The single-nucleotide difference at the site may have caused the low ratio, whereas one nucleotide substitution near the 3’-end of the 20 bp did not matter (Fig. 4A–D). The allodiploid strain S. pastorianus W34/70 harbors Sc-type and Se-type LEU2 genes, similar to the strain CBS1513^T^ (Table 2 (B)). The guide sequence type Sc-SeLEU2 in the Cas9 plasmid was identical to the corresponding regions in the two LEU2 genes in W34/70 (Fig. 4F) and CBS1513^T^ genomes (Table 2 (B)). One possible reason for the low Leu^−^ ratio is that an imaginary higher copy number of the genomic LEU2 gene caused the lower Leu^−^ mutant ratio. In fact, de la Cerda Garcia-Caro et al. (2022) reported that the S. pastorianus W34/70 strain has multiple copies of ScLEU2 and SeLEU2. It is feasible that the higher copy number of the LEU2 gene in the genome caused the lower Leu^−^ mutant ratio.
In this study, we also predicted applicability of the LEU2-targeting plasmids and the template DNAs to other Saccharomyces sensu stricto species, namely, S. cariocanus, S. mikatae, S. arboricolus, S. eubayanus, and S. uvarum.
In conclusion, the series of CRISPR-Cas9 plasmids constructed in this study enabled the generation of leu2 mutant strains from prototrophic strains of Saccharomyces sensu stricto species, including natural isolate, industrial, and allodiploid strains.