PpGATA21 Enhances the Expression of PpGA2ox7 to Regulate the Mechanism of Cerasus humilis Rootstock-Mediated Dwarf in Peach Trees
Xiuzhen Li, Ruxin Wang, Yuman Wang, Xueqiang Li, Qiaofang Shi, Yihe Yu

TL;DR
Cerasus humilis rootstocks cause dwarfing in peach trees by reducing gibberellin levels through the activation of PpGA2ox7 by PpGATA21.
Contribution
Identifies PpGATA21 as a transcription factor that activates PpGA2ox7, linking GA metabolism to dwarfing in peach trees.
Findings
PpGA2ox7 expression increases in Cerasus humilis grafted plants, correlating with reduced plant height and internode length.
PpGATA21 binds to and activates the PpGA2ox7 promoter, leading to decreased gibberellin levels.
Grafting onto Cerasus humilis significantly reduces GA levels and promotes dwarfing in peach trees.
Abstract
Dwarfing rootstocks enhance planting density, lower tree height, and reduce both labor in peach production. Cerasus humilis is distinguished by its dwarf stature, rapid growth, and robust fruiting capabilities, presenting substantial potential for further development. In this study, Ruipan 4 was used as the scion and grafted onto Amygdalus persica and Cerasus humilis, respectively. The results indicate that compared to grafting combination R/M (Ruipan 4/Amygdalus persica), grafting combination R/O (Ruipan 4/Cerasus humilis) plants show a significant reduction in height and a significant increase in flower buds. RNA-seq indicates that genes related to gibberellin (GA) and auxin metabolism are involved in the dwarfing process of scions mediated by C. humilis. The expression levels of the GA metabolism-related gene PpGA2ox7 significantly increased in R/O and are strongly correlated with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
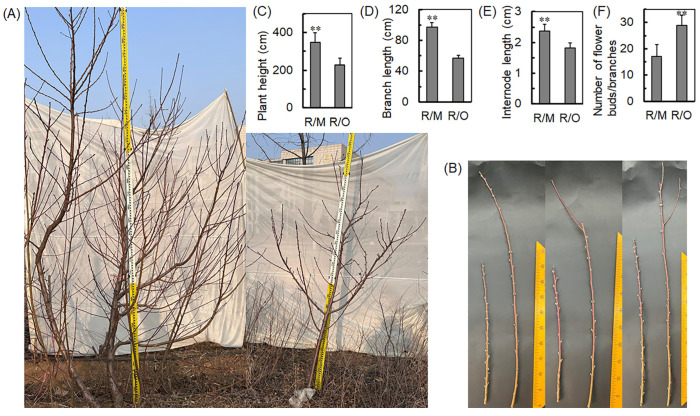 Figure 1
Figure 1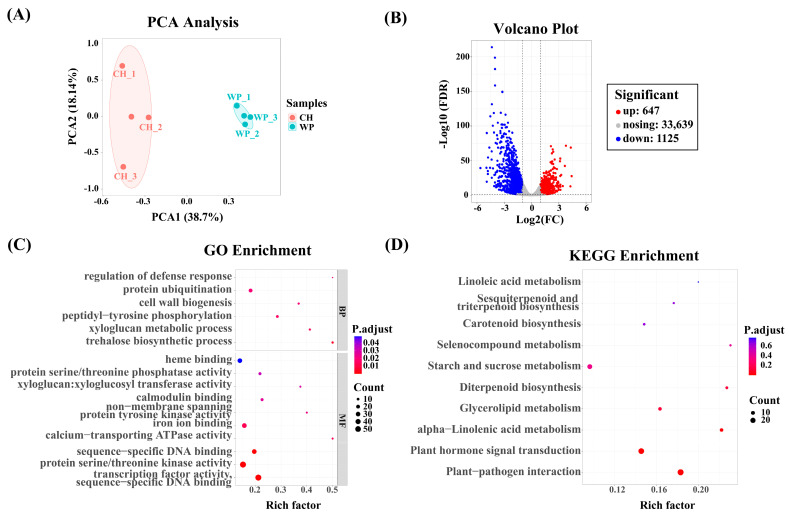 Figure 2
Figure 2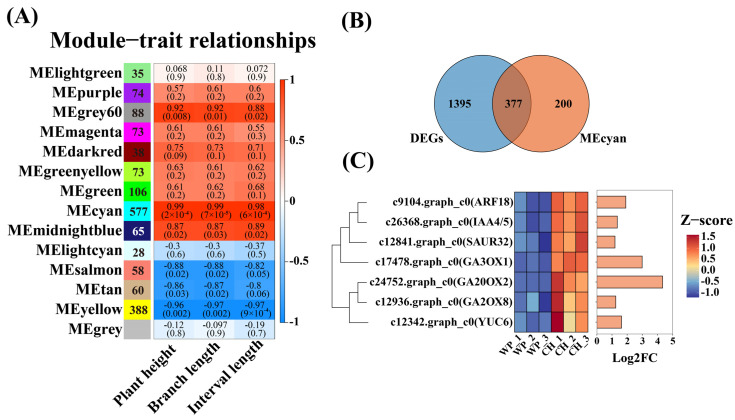 Figure 3
Figure 3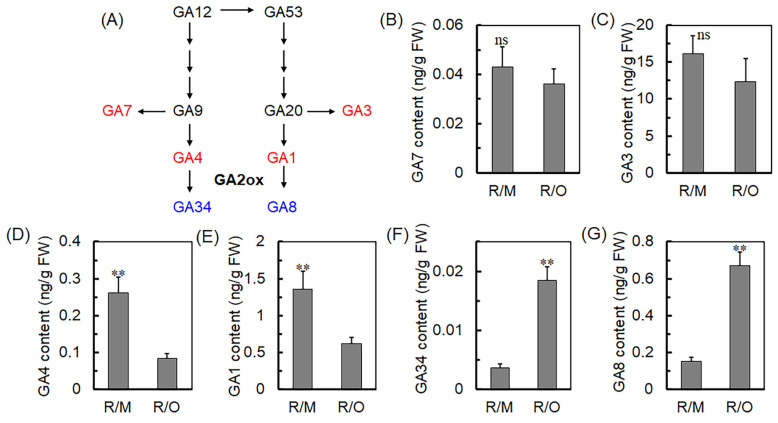 Figure 4
Figure 4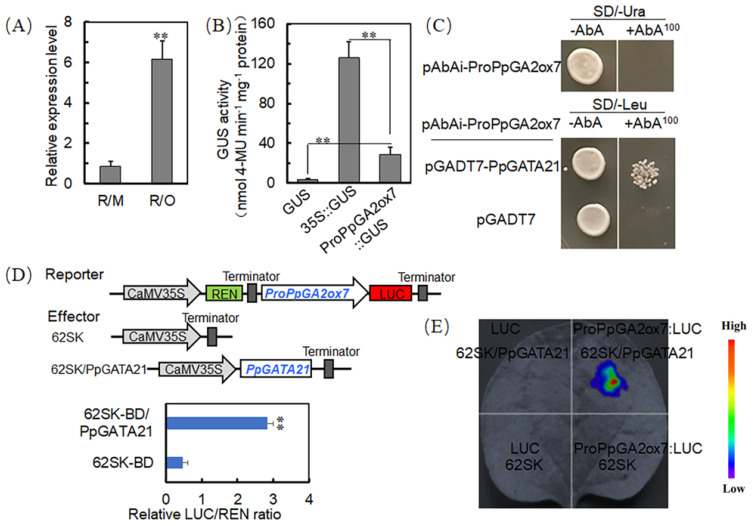 Figure 5
Figure 5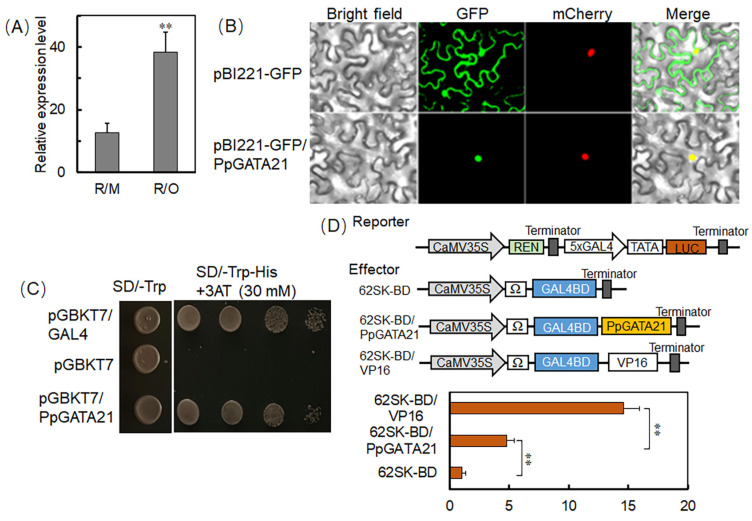 Figure 6
Figure 6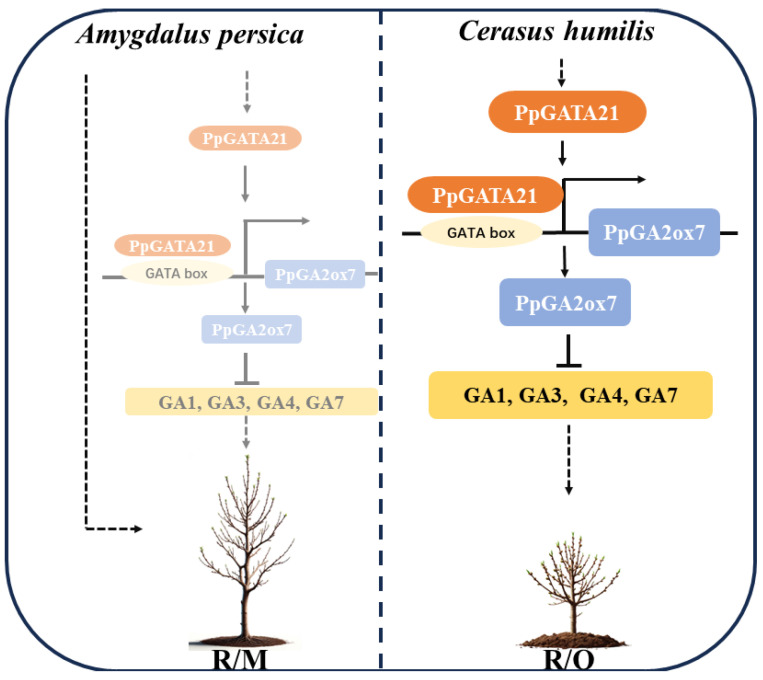 Figure 7
Figure 7- —Henan Province Science and Technology Key Project
- —Program for Science & Technology Innovation Talents in Universities of Henan Province
- —Top Young Talents in Central Plains
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Reproductive Biology · Plant Physiology and Cultivation Studies
1. Introduction
Peach (Prunus persica L. Batsch) is a globally important economic fruit. However, peach trees exhibit vigorous growth, produce numerous branches, and grow rapidly in the early stages, leading to high management and labor costs. With the establishment of modern standardized orchards, growers need to enhance standardized cultivation techniques for peach trees and the mechanization of orchard management [1,2]. The utilization of dwarf rootstocks can facilitate the transition of fruit trees from vegetative growth to reproductive growth, leading to earlier flowering and fruiting, enhancing early economic returns, and improving fruit quality [3,4]. Furthermore, the compact size of dwarf rootstocks makes them ideal for high-density planting, simplifying management during production [5]. Therefore, to align with the development trends of modern orchards, selecting dwarfing rootstocks, controlling tree vigor, and adopting low-density cultivation techniques have become primary objectives in peach tree breeding. Grafting is widely employed in horticulture to enhance plant growth characteristics, including improved tolerance, fruit quality, and dwarfing [6,7]. Research has demonstrated that macromolecules transported via vascular tissue between rootstock and scion play a crucial role in graft-induced phenotypic changes [8]. PbWoxT1 of tobacco rootstocks can be transferred to wild-type tobacco scions, influencing flower morphology [9]. Expression of CcFT3 in the phloem tissues of citrus can induce precocious flowering in grafted scions [10]. However, the molecular mechanisms underlying the interactions between rootstocks and scions remain largely unknown.
The selection of peach rootstock significantly influences key traits of peach scion varieties and greatly affects the profitability of peach orchards [11,12,13]. Additionally, introducing dwarfing traits can help control the growth potential of cash crops, including fruit trees [14]. Currently, grafting seedlings is the predominant method used in peach tree production, with Amygdalus davidiana (Carr.) Yu serving as a common rootstock for peach trees [1]. Differences in carbon allocation among various rootstocks influence the reduction of stem elongation [15]. Reduced canopy development (stem elongation) alters the light environment by decreasing intra-canopy shade for developing fruits, which in turn enhances fruit quality [16]. Therefore, the optimal selection and use of dwarfing rootstocks for peaches can enhance light availability and the uniformity of light distribution within the canopy, thereby improving fruit quality and reducing both management and labor costs in orchards [17]. Cerasus humilis, also known as the Chinese dwarf cherry or ‘calcium fruit’, is a fruit-bearing shrub native to northern China [18]. It is characterized by its short stature, rapid growth, and strong fruiting ability. Research has demonstrated that C. humilis can upregulate the expression of growth-related genes such as PpYucca5, PpYucca2, and PpYucca6, thereby reducing auxin levels and achieving a dwarfing effect [1]. However, the molecular mechanisms of dwarfing induced by C. humilis rootstock is still poorly understood.
Hormonal regulation is regarded as the mechanism through which rootstocks impact scion viability by modulating chemical signals from root tips [7]. Alterations in cell elongation and division, controlled by various plant hormones including auxins, cytokinins, gibberellins (GAs), abscisic acid, and brassinosteroids, result in a dwarf phenotype [15,19,20]. GAs are essential for plant vascular development, cambial function, and the growth of xylem and phloem tissues [21]. There is significant evidence that metabolism of GA is crucial in causing dwarfing of plant shoots [22,23]. GA 2-oxidases (GA2oxs) are key enzymes in GA metabolism that degrade the active gibberellins GA1 and GA4 into the inactive forms GA8 and GA34. This transformation is a critical process in the regulation of biologically active GA levels in plants [24]. Currently, only a select few gibberellin molecules such as GA1, GA3, GA4, and GA7 exhibit biological activity in plants.
In this study, the mechanism by which C. humilis mediates dwarfing in peach trees through the regulation of GA metabolism was analyzed. This study aims to explain the mechanism by which C. humilis rootstocks influence the dwarfing of peach trees, providing a robust theoretical basis for using these rootstocks to regulate tree height.
2. Result
2.1. C. humilis Significantly Reduces the Height of Scions
Ruipan 4 was used as the scion and grafted onto Amygdalus persica and Cerasus humilis, respectively. The results indicate that rootstocks significantly influence the growth vigor of grafted peach trees (Figure 1). Compared to R/M (Ruipan 4/Amygdalus persica), R/O (Ruipan 4/Cerasus humilis) significantly reduces the growth height of peach trees, evidenced by notable decreases in plant height (Figure 1A,C), branch length (Figure 1B,D), and internode length (Figure 1B,E). Furthermore, compared to R/M, R/O increases the number of flower buds by 62.5% (Figure 1B,F).
2.2. Differentially Expressed Genes (DEGs) between R/O and R/M
To reveal the potential molecular networks involved in the dwarfing of peach trees mediated by C. humilis rootstocks, RNA-seq was conducted on leaves of R/O and R/M. Principal component analysis (PCA) demonstrates that the three biological replicates of R/M and R/O cluster together, indicating that the samples are reliable and consistent (Figure 2A). To understand the potential regulatory mechanisms, the number of DEGs was quantified, and genes with a |log_2_ FC| > 1 were selected for further analysis. The results indicate that there are 1772 differentially expressed genes between R/O and R/M, with 647 genes upregulated and 1125 genes downregulated (Figure 2B).
For further analysis, we identified 1772 DEGs for Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis. The GO enrichment analysis revealed that the DEGs were significantly enriched in molecular functions, including transcription factor activity, sequence-specific DNA binding (GO:0003700), calcium-transporting ATPase activity (GO:0005388), and iron ion binding (GO:0005506) (Figure 2C). The ten most representative KEGG pathways are illustrated in Figure 2D, with the most significant being alpha-linoleic acid metabolism (ko00592), plant hormone signal transduction (ko04075), and plant–pathogen interactions (ko04626).
2.3. Weighted Gene Co-Expression Network Analysis (WGCNA Analysis)
To investigate the gene regulatory networks involved in the dwarfing process of peach trees mediated by C. humilis rootstocks, a comprehensive transcriptome-wide WGCNA was conducted on all expressed genes (15,540 genes), leading to the identification of key genes within the network. Co-expression modules were constructed using the Pearson correlation coefficient of gene expression in all samples, resulting in a total of 14 modules (Figure 3A). Each grid in Figure 3A is color-coded based on statistical significance, with each grid labeled with two numbers: the correlation coefficient and the p-value. The leftmost side of the grid represents each module and the number of genes enriched. The analysis revealed that there were 577 genes enriched in the MEcyan module, which is significantly correlated with plant height, branch length, and internode length, exhibiting a correlation coefficient of up to 0.99 (Figure 3A). As a result, the genes within the MEcyan module are deemed as a priority for future investigation. Further analysis revealed that 377 genes within the MEcyan module are differentially expressed (Figure 3B). Among these 377 genes associated with plant height, branch length, and internode length, studies focused on genes related to auxin (ARF18, IAA4/5, SAUR32, YUC6) and gibberellins (GA3ox1, GA2ox2, GA2ox7) (Figure 3C). Importantly, the transcription levels of GA2ox-related genes significantly increased. Since GA2ox plays a crucial role in the degradation of active GAs, the metabolism of GAs and the expression of GA2ox genes may be linked to the dwarfing mechanism of peach trees mediated by C. humilis.
2.4. PpGA2ox7 Regulates GA Levels in Peach Trees
To further understand the role of GAs in the dwarfing mechanism of peach trees mediated by C. humilis rootstocks, the GA biosynthesis pathway was used as the starting point (Figure 4A). The levels of GA1, GA3, GA4, GA7, GA8, and GA34 in R/O and R/M were measured (Figure 4B–G). The results indicate that the levels of GA1, GA3, GA4, and GA7 in R/O are significantly lower than those in R/M (Figure 4B–E), while the levels of GA8 and GA34 are significantly higher relative to R/M (Figure 4F,G). These findings suggest that C. humilis rootstocks can alter the levels of six GAs in the biosynthesis pathway, potentially through the expression of the GA2ox family. It was observed that PpGA2ox7 were significantly upregulated in C. humilis rootstocks (Figure 3C), which might influence the dwarfing effect of the rootstocks. Consequently, further investigations into PpGA2ox7 were conducted in this study.
2.5. PpGATA21 Directly Binds and Activates PpGA2ox7 Expression
RT-qPCR results demonstrate that PpGA2ox7 is significantly upregulated in R/O (Figure 5A), suggesting it may respond to the dwarfing effect mediated by C. humilis rootstocks. GUS assay results indicate that, compared to the negative control, the PpGA2ox7 promoter activates the expression of the GUS reporter gene (Figure 5B), demonstrating that this promoter can initiate gene expression.
The PpGA2ox7 promoter was used as bait; yeast one-hybrid (Y1H) screening identified 37 positive colonies, among which three encoded the gene LOC18792628. This gene showed high homology with the Arabidopsis GATA-type transcription factor GATA21, and was therefore named PpGATA21 (XM_020560109, LOC18792628).
To investigate whether PpGATA21 regulates PpGA2ox7, a recombinant plasmid was transformed into Y1H Gold yeast competent cells using a Y1H. Yeast strains co-transformed with the empty pGADT7 and pGADT7-PpGATA21/pABAi-PpGA2ox7 exhibited growth on the SD/-Leu medium, indicating successful transformation. Yeast strains transformed with the empty pGADT7 vector were unable to grow on SD/-Leu plates containing 100 ng/mL AbA, whereas strains transformed with pGADT7-PpGATA21/pABAi-PpGA2ox7 exhibited growth (Figure 5C), the results indicate that PpGATA21 can directly bind to the PpGA2ox7 promoter.
The PpGA2ox7 promoter was cloned into the pGreenII0800-LUC vector, resulting in the reporter plasmid PpGA2ox7-LUC0800. The full-length PpGATA21 was cloned into the 62SK vector, resulting in the effector plasmid PpGATA21-62SK (Figure 5D). The recombinant plasmid was created and then introduced into the A. tumefaciens GV3101 strain along with the pSoup helper plasmid. For the experimental group, the combination (ProPpGA2ox7/LUC + PpGATA21/62SK) was used, with dual empty vector controls (LUC + 62SK) and single empty vector controls (LUC + PpGATA21/62SK and ProPpGA2ox7/LUC + 62SK) serving as comparisons. The ratio of LUC to REN demonstrates that PpGATA21 positively regulates the expression of PpGA2ox7 (Figure 5D,E).
2.6. PpTATA21 Is a GATA-Type Transcriptional Activator
RT-qPCR results demonstrate that, compared to R/M, the expression of PpGATA21 in R/O is significantly increased (Figure 6A), suggesting that PpTATA21 is involved in the dwarfing process of peach trees induced by C. humilis. Subcellular localization revealed that the fluorescence of PpTATA21 overlaps with the nuclear-localized mCherry, indicating that this gene is localized to the nucleus (Figure 6B). Further analysis was conducted to assess the transcriptional activity of PpTATA21, using pGBKT7 as the negative control and pGBKT7/GAL4 as the positive control. Yeast cells containing pGBKT7-PpGATA21 were able to grow on plates containing 3AT, indicating that PpGATA21 possesses transcriptional activation activity (Figure 6C). Further, the LUC/REN ratio of 62SK/BD-PpTATA21 was significantly higher than that of the negative control 62SK/BD, as determined by the dual-luciferase reporter (DLR) (Figure 6D).
3. Discussion
Plums are particularly focused on rootstock breeding as donors for ecological compatibility or resistance [25,26]. Reports suggest that among all Prunus species, Prunus cerasifera shows the highest diversity in both ecological adaptation and morphological traits [27]. Compared to other types of rootstocks, C. humilis is particularly favored for its dwarf stature and vigorous growth. Therefore, exploring and analyzing the dwarfing effect of C. humilis on scions is of significant importance [28]. The findings of this study contribute to a deeper understanding of the working mechanisms of C. humilis and how it mediates the dwarfing of peach scions. In this study, C. humilis significantly reduced the growth height of peach trees and can increase the number of flower buds by 62.4% (Figure 1). The increase in the number of flower buds on scions is attributed to the fact that scions on dwarf rootstocks can cease branch growth earlier and enhance the assimilation and allocation of nutrients toward bud production [3,29]. RNA-seq data identified a total of 1772 DEGs in C. humilis. GO and KEGG analysis highlighted the significance of transcription factor activity and plant hormone signal transduction in the process of C. humilis-mediated peach scion dwarfing (Figure 2). Furthermore, WGCNA revealed 577 genes enriched in the MEcyan module, which were closely associated with scion plant height, branch length, and internode length, with 377 genes showing differential expression (Figure 3). Previous research has demonstrated that weak citrus rootstocks TO and FD exhibit reduced plant height, plant weight, and internode length compared to the strong CC rootstock [30].
Increasing evidence suggests that plant hormones play a role in the dwarfing process of plants [31,32,33,34]. The key gene DkGA2ox1 in persimmon dwarf disease acts as a signaling stimulus affecting the levels of GAs in scions, and overexpression of DkGA2ox1 results in a typical dwarf phenotype [22]. An upregulation of GAox genes has been observed in dwarf lychee varieties, suggesting that GAs play a crucial role in mediating significant differences between dwarf and robust lychee varieties [35]. Consistently, our study found that C. humilis rootstocks alter the synthesis pathway of six types of GAs in peach scions, with significant reductions in the levels of GA1, GA3, GA4, and GA7. The GA2ox family genes are essential in the GA biosynthetic pathway, as illustrated in Figure 4A. The research pinpointed PpGA2ox7, a crucial gene in GA metabolism, as significantly linked to the alterations in plant height, branch length, and internode length influenced by C. humilis rootstock (Figure 3C). This suggests a potential role in inducing dwarfism in the scion through GA metabolism. Similarly, the transcription factor ABI4 directly binds to promoters and activates the transcription of GA2ox7, and mutations in GA2ox7 rescue the dwarf phenotype caused by reduced GA levels in plants overexpressing ABI4 [36]. These results indicate that C. humilis rootstocks induce dwarfing in peach scions by reducing the levels of GA, which is attributed to increased expression of PpGA2ox7.
Transcription factors are essential for the regulation of plant growth, development, stress responses, and hormone signaling [37,38,39]. The expression of hormone-related metabolic genes is continually proven to be regulated by transcription factors [40]. Specifically, the transcription factor JUB1 directly inhibits the biosynthesis genes GA3ox1 and DWARF4 (DWF4), leading to a decrease in levels of GAs and brassinosteroids (BRs). This reduction leads to the development of a characteristic phenotype associated with GA/BR deficiency, including shortened hypocotyls, dwarfism, delayed blossoming, and infertility in male plants [40]. Y1H screening was utilized in this study to identify the transcription factor PpGATA21 of PpGA2ox7. The research revealed that PpGATA21 is a nuclear-localized GATA-type transcription factor, and its expression is upregulated by R/O induction. PpGATA21 is primarily described as an inhibitor within the gibberellin signaling pathway [41]. Transcriptional activation analysis confirmed that PpGATA21 exhibits transcriptional activity (Figure 4). Previous studies have shown that in addition to GA signaling, GATA-type transcription factors also participate in other pathways, including those involving abscisic acid [42] and cytokinins [43]. This study utilized Y1H and DLR techniques to confirm that PpGATA21 plays a positive role in regulating the expression of PpGA2ox7, thereby influencing the involvement of GAs in the dwarfing mechanism of peach scions grafted onto C. humilis rootstocks (Figure 3). Similar to GATA21, the poplar transcription factor PdeGATA3 suppresses GA biosynthesis by inhibiting the expression of PdeGA20ox, leading to dwarfism in poplar trees [44].
Our working model, as shown in Figure 7, portrays the positive regulation of PpGA2ox7 expression by PpGATA21 transcription under the influence of C. humilis rootstocks. On the other hand, C. humilis rootstocks suppress GA biosynthesis by activating the expression of PpGA2ox7 (Figure 1 and Figure 7). Since optimal hormone levels are essential for regular plant growth, the regulation of PpGATA21 by GAs could play a vital role in the accurate management of dwarfing.
4. Materials and Methods
4.1. Fruit Materials and Treatments
Ruipan 4 was used as the scion and grafted onto Amygdalus persica and Cerasus humilis, respectively. The graft combinations were R/O (Ruipan 4/Cerasus humilis (Bge.) Sok) and R/M (Ruipan 4/Amygdalus persica L.), sourced from Luoyang, Henan (34°37′ N, 112°38′ E). In the experimental region, the annual average temperature is 14.2 °C. Traditional field management practices are followed, including the application of organic fertilizer in the fall and urea before bud break in the spring. Additionally, flooding irrigation is utilized both before spring bud break and after autumn leaf fall, with adjustments based on weather conditions. In 2020, one-year-old C. humilis and A. persica rootstocks were grafted with Ruipan 4 scions. The grafted trees experienced growth during the period from 2020 to 2024. Sampling was performed on the current year’s shoots of young trees three years after grafting (May 2023) for RNA-seq, RNA extraction, and hormone content analysis. The study included sampling from four distinct trees and the experiment was conducted three times, totaling 24 trees (R/O and R/M). Each tree was situated 2.0–2.4 m apart.
4.2. Measurement of Plant Growth Parameters
Measurement data for scion growth were recorded, including plant height (cm), branch length (cm), average internode length (cm), and the number of flowers. (1) Plant height and branch length were measured using a tape measure; (2) The number of nodes on the branches above the grafted scion was calculated by counting; (3) Internode length was calculated by dividing the scion length by the total number of nodes; (4) The number of flowers on the scion was determined using the counting method.
4.3. RNA Extraction and RNA-Seq Analysis
The extraction of total RNA was carried out with the FastPure Cell/Tissue Total RNA Isolation Kit (Vazyme, Nanjing, China), followed by quantification of RNA concentration using a Nanodrop 2000 (Thermo Scientific, Waltham, MA, USA). Sequencing was performed by Beijing Biomarker Technologies Co., Ltd. (Beijing, China). Principal component analysis (PCA) was conducted on all expressed genes using count data. The results of PCA were visualized with the R packages factoextra (1.0.7) and FactoMineR (2.11). Differentially expressed genes (DEGs) were identified using DESeq2 (1.42.1), with a |log_2_ fold change (log_2_FC)| ≥ 1.0 set as the threshold for differential gene screening and an adjusted p-value (padj) < 0.05, resulting in the identification of 1772 differential genes. Weighted Gene Co-expression Network Analysis (WGCNA) was performed using R software (version 4.0.2) to analyze all 15,540 expressed genes, requiring a TPM (Transcripts Per Million) > 1 and the soft threshold was set at 6. We utilized TBtools (2.070) for visualizing WGCNA and generating heat maps [45] and to illustrate the overlapping relationship between all DEGs (1772) and MEcyan in WGCNA. R software was utilized to conduct Gene Ontology (GO) enrichment analysis on 1772 differential genes, identifying significantly enriched GO pathways with corrected p-values < 0.05. A script written in R language was employed to perform Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis on the same set of 1772 differential genes. Multiple testing correction was applied using the false discovery rate method. KEGG pathways with corrected p-values < 0.05 were significantly enriched. The expression of all genes in the transcriptome are provided in Supplementary Table S2.
4.4. RT-qPCR
RNA was reverse transcribed into cDNA using the NovoScript^®^ Plus All-in-one 1st Strand cDNA Synthesis SuperMix (gDNA Purge) (Novoprotein, Suzhou, China). RT-qPCR was conducted in accordance with the instructions provided by the Vazyme ChamQ Universal SYBR qPCR Master Mix (Vazyme, Nanjing, China) and carried out on a CFX96 Real-Time PCR Detection System 574 (Bio-Rad, Hercules, CA, USA). Data analysis was conducted using the 2^−ΔΔCt^ method, with 18S rRNA [46] serving as the internal reference gene. All experiments included three biological replicates. The primers utilized for RT-qPCR are provided in Supplementary Table S1. The melt curve in RT-qPCR experiment is provided in Supplementary Figure S1.
4.5. Quantification of Endogenous Hormones
Plant hormone contents were measured according to methods described in previous studies [47]. Each sample was weighed to 0.2 g and ground into powder using liquid nitrogen. Hormone contents were then measured with a Qtrap 5500 System mass spectrometer (AB Sciex, Shanghai, China), using standards from Sigma Chemical Co. (St. Louis, MO, USA). Each experiment was conducted with three biological replicates.
4.6. Quantitative GUS Assay
The promoter of PpGA2ox7 was inserted into the vector pC0390-35S-GUS (PpGA2ox7::GUS) for the purpose of activating the GUS reporter gene. The GUS empty vector was used as the negative control, and the 35S::GUS vector served as the positive control. Nicotiana benthamiana leaves were immersed in the inoculated solution and placed under vacuum until water-soaked lesions appeared. After treatment, the collected leaf samples were pulverized into powder using liquid nitrogen, and the measurement of GUS activity was conducted with the use of a β-Glucosidase (β-GC) Activity Assay Kit (Boxbio, Beijing, China) for the GUS gene.
4.7. Yeast One-Hybrid (Y1H)
A cDNA library was constructed from mRNA extracted from the leaves of peach trees. Yeast one-hybrid screening was conducted using the Matchmaker Gold Yeast One-Hybrid Library Screening System (Clontech, San Francisco, CA, USA). The full-length sequence of PpGATA21 was cloned into the pGADT7 vector (pGADT7-PpGATA21), and subsequently transformed into bait-reporter cells. An empty pGADT7 vector was used as the control. Screening was conducted on SD/-Leu and supplemented with 100 ng/mL aureobasidin A (SD/-Leu + AbA^100^). The PpGA2ox7 promoter was inserted into the pAbAi vector (pAbAi-ProPpGA2ox7) and transformed into Y1H Gold cells to establish bait-reporter cells. Screening was performed on SD/-Ura medium and supplemented with 100 ng/mL aureobasidin A (SD/-Ura + AbA^100^).
4.8. Transcriptional Activation Analysis in Yeast Cells
The CDS sequence of PpGATA21 was inserted into the pGBKT7 plasmid (pGBKT7/PpGATA21), and the plasmid was transformed into Y2H Gold cells. The pGBKT7-GAL4 plasmid served as the positive control, and the empty pGBKT7 vector was used as the negative control. Transformed yeast cells were plated on SD/-Trp and SD/-Trp-His + 3AT (50 mM) selective media.
4.9. Subcellular Localization Analysis
The full-length coding sequence of PpGATA21 (excluding the stop codon) was amplified from leaf cDNA, cloned into the pBI221-GFP vector, and used to create a pBI221-GFP/PpGATA21 fusion construct. Subsequently, the fusion construct was co-transformed with the nuclear marker mCherry into N. benthamiana leaves, and fluorescence signals were then observed using a laser scanning confocal microscope (Olympus, Tokyo, Japan).
4.10. Dual-Luciferase Reporter (DLR)
The promoter sequence of PpGA2ox7 was cloned into the pGreenII0800-LUC vector to generate a reporter construct. The CDS of PpGATA21 was cloned into the pGreen II 62-SK plasmid to produce an effector construct, with pGreenII 0800-LUC and pGreen II 62-SK serving as negative controls. Constructs containing reporter and effector genes were transformed into Agrobacterium tumefaciens strain GV3101 (pSoup, St. Louis, MO, USA), and subsequently injected into N. benthamiana leaves. The Dual-Luciferase Reporter Assay System (Promega, Madison, WI, USA) was used to measure dual-luciferase activity in the injected N. benthamiana leaves.
4.11. Statistical Analysis
Data were statistically analyzed using Microsoft Excel software (2019). Statistical significance between two samples was assessed using Student’s t-test (** p < 0.01).
5. Conclusions
The dwarfing effect of C. humilis on peach scions is pronounced, and it also increases the number of flower buds. This study delves into a less explored area, investigating the impact of grafting at the molecular level. The result indicates that C. humilis enhances the expression of the GA metabolic gene PpGA2ox7 by promoting the expression of PpGATA21, thereby reducing the levels of GA and consequently leading to the dwarfing of peach scions (Figure 7). Through the modulation of GA levels, C. humilis rootstock shows promise as a dwarfing rootstock for peach trees, offering valuable insights for future researchers in selecting suitable rootstocks for breeding programs or establishing modern orchards.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li X. Wang Y. Zhao L. Chen S. Yuan Y. Wei T. Geng J. Effects of Cerasus humilis (Bge). Sok. Rootstock on Peach Growth, Development, and Expression of Growth-Related Genes Horticulturae 2023957610.3390/horticulturae 9050576 · doi ↗
- 2Neri D. Crescenzi S. Massetani F. Manganaris G.A. Giorgi V. Current trends and future perspectives towards sustainable and economically viable peach training systems Sci. Hortic.202230511134810.1016/j.scienta.2022.111348 · doi ↗
- 3Basile B. De Jong T.M. Control of fruit tree vigor induced by dwarfing rootstocks Hortic. Rev.201846399710.1002/9781119521082.ch 2 · doi ↗
- 4Shi J. Dong Z. Song C. Xie B. Zheng X. Song S. Jiao J. Wang M. Bai T. Establishment of an efficient micropropagation system in enhancing rooting efficiency via stem cuttings of apple rootstock M 9T 337Hortic. Sci.202148637210.17221/106/2020-HORTSCI · doi ↗
- 5Habibi F. Liu T. Folta K. Sarkhosh A. Physiological, biochemical, and molecular aspects of grafting in fruit trees Hortic. Res.20229 uhac 03210.1093/hr/uhac 03235184166 PMC 8976691 · doi ↗ · pubmed ↗
- 6Hayat F. Ma C. Iqbal S. Ma Y. Khanum F. Tariq R. Altaf M.A. Khan U. Coulibaly D. Huang X. Comprehensive transcriptome profiling and hormonal signaling reveals important mechanism related to dwarfing effect of rootstocks on scion in Japanese apricot (Prunus mume)Sci. Hortic.202332111226710.1016/j.scienta.2023.112267 · doi ↗
- 7Hayat F. Li J. Liu W. Li C. Song W. Iqbal S. Khan U. Javed H.U. Altaf M.A. Tu P. Influence of citrus rootstocks on scion growth, hormone levels, and metabolites profile of ‘Shatangju’ mandarin (Citrus reticulata Blanco)Horticulturae 2022860810.3390/horticulturae 8070608 · doi ↗
- 8Li W. Chu C. Li H. Zhang H. Sun H. Wang S. Wang Z. Li Y. Foster T.M. López-Girona E. Near-gapless and haplotype-resolved apple genomes provide insights into the genetic basis of rootstock-induced dwarfing Nat. Genet.20245650551610.1038/s 41588-024-01657-238347217 · doi ↗ · pubmed ↗