Clustered Regularly Interspaced Short Palindromic Repeat-1 (CRISPR-1) Locus as a Tool for Tracing the Zoonotic History of Salmonella enterica Strains
Maan Neamah, Evan Mahdi, Muhammed Sameir, Safin Hussein, Abdulmalik Saber

TL;DR
This study uses CRISPR sequences to trace the zoonotic transmission of Salmonella strains between humans and camels.
Contribution
The study demonstrates the utility of CRISPR-1 locus IGS diversity for high-resolution genotyping and zoonotic tracking of Salmonella strains.
Findings
Five CRISPR-1 types were identified in human isolates and three in camel isolates.
Shared IGSs suggest zoonotic or reverse-zoonotic transmission between humans and camels.
CRISPR-1 profiling reclassified camel isolates as S. enterica serovar Enteritidis with higher accuracy than traditional methods.
Abstract
Background Salmonella enterica is a significant foodborne pathogen that causes considerable illness and death in humans and animals. The clustered regularly interspaced short palindromic repeat (CRISPR)-CRISPR-associated protein (Cas) system in bacteria acts as an adaptive immune defense against invasive genetic elements by incorporating short intergenic spacers (IGSs) into CRISPR loci. These loci serve as molecular records of past interactions with phages and plasmids, providing insights into the transmission and evolution of bacterial strains across different hosts. Aim This study aimed to investigate the diversity of IGSs in the CRISPR-1 locus of S. enterica isolates from humans and camels. The objective was to assess the potential of IGSs to distinguish strains, track sources, and understand patterns of zoonotic transmission. Materials and methods Genomic DNA was extracted from…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
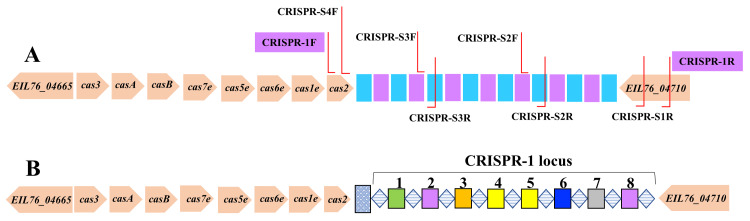 Figure 1
Figure 1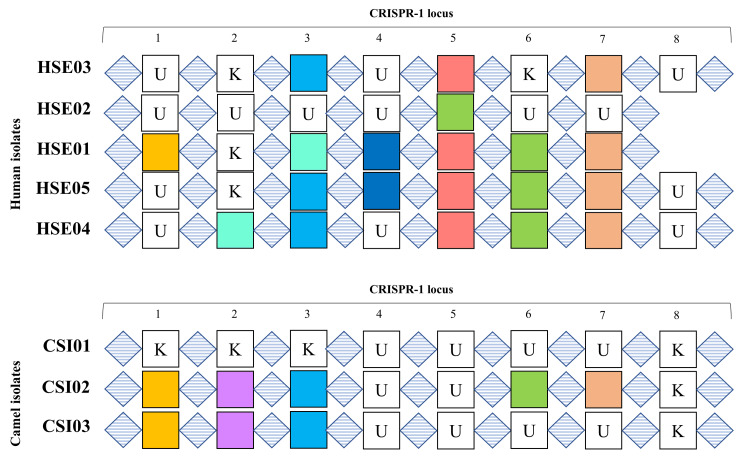 Figure 2
Figure 2| Set | Total number of IGSs | Number of known IGSs | Number of UIGSs |
| HSE01 | 7 | 0 | 7 |
| HSE02 | 7 | 1 | 6 |
| HSE03 | 8 | 5 | 3 |
| HSE04 | 8 | 5 | 3 |
| HSE05 | 8 | 6 | 2 |
| CSI03 | 8 | 4 | 4 |
| CSI03 | 8 | 6 | 2 |
| CSI03 | 8 | 4 | 4 |
|
| |||||
| HSE01 | HSE02 | HSE03 | HSE04 | HSE05 | Identity |
| IGS3 | IGS3 | 100% | |||
| IGS3 | IGS2 | 97% | |||
| IGS4 | IGS4 | 100% | |||
| IGS5 | IGS5 | IGS5 | IGS5 | 100% | |
| IGS6 | IGS5 | IGS6 | IGS6 | 100% | |
| IGS7 | IGS7 | IGS7 | IGS7 | 100% | |
|
| |||
| CSI01 | CSI02 | CSI03 | Identity |
| IGS1 | IGS1 | 100% | |
| IGS2 | IGS2 | 100% | |
| IGS3 | IGS3 | 100% | |
|
|
| Identity | ||||||
| HSE01 | HSE02 | HSE03 | HSE04 | HSE05 | CSI01 | CSI02 | CSI03 | |
| IGS1 | IGS1 | IGS1 | 100% | |||||
| IGS3 | IGS3 | IGS3 | IGS3 | IGS3 | 100% | |||
| IGS6 | IGS5 | IGS6 | IGS6 | 100% | ||||
| IGS7 | IGS7 | IGS7 | IGS7 | IGS7 | 100% | |||
|
| ||||
| HSE01 | HSE02 | HSE03 | HSE04 | HSE05 |
| UIGS1 | UIGS1 | |||
| UIGS2 | UIGS2 | |||
| UIGS6 | UIGS3 | UIGS3 | UIGS2 | |
|
| ||
| CSI01 | CSI02 | CSI03 |
| UIGS1 | UIGS1 | |
| UIGS2 | UIGS2 | |
| UIGS3 | UIGS3 | |
| Set | Source | Identification system | |
| rrnH | CRISPR-1 locus | ||
| HSE01 | Human |
|
|
| HSE02 | Human |
|
|
| HSE03 | Human |
|
|
| HSE04 | Human |
|
|
| HSE05 | Human |
|
|
| CSI01 | Camel |
|
|
| CSI02 | Camel |
|
|
| CSI03 | Camel |
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · CRISPR and Genetic Engineering · Vibrio bacteria research studies
Introduction
Serovars of Salmonella enterica are significant pathogens in the global foodborne context, causing a considerable number of cases of gastroenteritis and resulting in thousands of deaths annually [1]. This bacterium has developed various defense mechanisms against bacteriophages, which can infect and lyse bacterial cells. As a result, bacteria have evolved an array of mechanisms to resist viruses. Among the most complex defense systems is the clustered regularly interspaced short palindromic repeat (CRISPR)-CRISPR-associated protein (Cas) system, which functions as an adaptive immune system, providing sequence-specific protection against invasive genetic elements such as phages and plasmids [2,3]. This defense mechanism is intricately connected to the pathogen's capacity to survive and proliferate in diverse hosts, underscoring the importance of comprehending its transmission dynamics.
The mechanisms and epidemiology of zoonotic and reverse-zoonotic transmission of S. enterica are crucial for understanding how this pathogen spreads among different hosts and environments. Zoonotic transmission, which occurs when pathogens are transferred from animals to humans, has been extensively studied for S. enterica. This transmission usually occurs when people ingest contaminated food, come into direct contact with infected animals, or are exposed to animal waste in the environment [4]. Serovars like *Salmonella *Enteritidis and *Salmonella *Typhimurium are commonly found in poultry and livestock and can cause outbreaks of foodborne illnesses in humans [5]. Additionally, these bacteria can carry antibiotic resistance genes, making treatment and control efforts more challenging. Recent studies emphasize the need for comprehensive surveillance and the implementation of biosecurity measures in animal farming to reduce the risk of zoonotic transmission to humans [6,7].
Reverse-zoonotic transmission, also known as anthroponosis, involves the transfer of pathogens from humans to animals. While not as well-documented as zoonotic transmission, this occurrence poses significant threats to animal well-being and has the potential to impact biodiversity. There have been reports of reverse-zoonotic events involving S. enterica, where strains originating from humans have been transmitted to animals, establishing new reservoirs of infection. Notably, instances of S. enterica isolates sourced from humans have been detected in animals, suggesting a complex interplay and bidirectional exchange of pathogens [8,9]. In these transmission pathways, the CRISPR-1 locus within S. enterica plays a vital role as a genetic repository of past interactions with phages and plasmids. Consequently, it facilitates the tracking of the pathogen's transmission history and evolutionary trajectory.
Understanding both zoonotic and reverse-zoonotic transmission mechanisms is crucial for developing effective control strategies. Enhanced genoserotyping techniques, such as CRISPR-based genotyping, offer higher resolution when tracking transmission routes compared to traditional methods. These techniques can identify specific intergenic spacers (IGSs) that act as molecular markers for different host interactions, providing valuable insights into the spread and persistence of the pathogen across human and animal populations. This knowledge is essential for formulating policies and implementing interventions aimed at controlling the spread of Salmonella and safeguarding public health. At the core of these genoserotyping techniques lies the CRISPR-Cas system.
The CRISPR-Cas system operates by integrating short IGS sequences into the bacterial genome's CRISPR loci. These IGSs come from exogenous genetic elements and are inserted at regular intervals among repetitive sequences, acting as molecular records. This mechanism of information storage allows the bacterial cell to identify and degrade future encounters with the same or similar genetic material [10]. The CRISPR loci themselves serve as a unique repository of past interactions between bacteria and invasive genetic components. This valuable information helps us understand bacterial population dynamics and their adaptive responses to environmental conditions over time [11].
Understanding the diversity and evolution of CRISPR loci in S. enterica is important for several reasons. Firstly, it can advance genotyping and source-tracking methods, which are crucial for epidemiological investigations and identifying potential contamination sources [12]. Additionally, it can provide valuable insights into the transmission of Salmonella strains among humans, animals, and other reservoirs, improving our understanding of pathogen transmission dynamics.
CRISPR loci are an ideal tool for studying zoonotic or reverse-zoonotic transmission because bacteria can acquire genetic material from viruses or other mobile genetic elements found in various host environments. This enables them to effectively capture their previous interactions. For instance, Salmonella strains can acquire and incorporate intergenic sequences from phages or plasmids that are specific to human, animal, and environmental reservoirs [13]. As a result, their CRISPR loci retain a genetic signature that can reveal their transmission patterns and evolutionary origins.
The primary goal of this study was to discover new arrays within Salmonella CRISPR loci and evaluate their efficacy in differentiating strains and tracing their sources. We aimed to develop a distinctive genoserotyping system using CRISPR profiles that could potentially enhance or outperform current methods. Furthermore, we examined the zoonotic relationship between Salmonella strains in humans and camels by comparing their CRISPR IGS content.
Materials and methods
Isolates of Salmonella
The Department of Veterinary Microbiology, Faculty of Veterinary Medicine, University of Kufa in Iraq has provided 20 bacterial isolates for this study. These isolates include two subspecies of S. enterica: 10 isolates of S. enterica subsp. *enterica *serovar Enteritidis obtained from humans and 10 isolates of S. enterica subsp. *enterica *serovar Indiana obtained from camels. The isolates were incubated overnight at 37°C in LB medium.
Genomic DNA extraction
The sediments from fresh *Salmonella *isolate cultures in LB broth media were obtained by centrifugation at 14,000 rpm for two minutes. The bacterial genomic DNA was extracted using a commercial DNA extraction kit (G-spin Genomic DNA Extraction Kit for Bacteria, 17121, Intron, Korea) following the manufacturer's instructions. The quality and quantity of the extracted DNA were assessed using a Quantus™ fluorometer (Promega, USA).
CRISPR-1 locus amplification and sequencing
The PCR was able to copy the CRISPR-1 locus region with a forward primer (CRSPR-1F) (5′-CGGAGTCGGAAACGTAGTAATG-3′) and a reverse primer (CRSPR-1R 5′-GAATGAGTGACGCTGAGAAGAA-3′). Primers were designed using Primer3. The PCR reactions were carried out using an Optimus 96G thermal cycler (QLS, UK) under the following conditions: initial denaturation at 94°C for five minutes, followed by 35 cycles of denaturation at 94°C for 30 seconds, annealing at 56°C for 30 seconds, extension at 72°C for four minutes, and a final extension step at 72°C for 10 minutes. Agarose gel electrophoresis was used to separate the amplified PCR products. The gel used was 1.5%, and the DNA dye used was RedSafe (Intron, Korea). The electrophoresis conditions were set at V: 90 for a duration of 42 minutes. A DNA ladder was included as a marker. Subsequently, the amplified PCR bands were concentrated using the Wizard® SV Gel and PCR Clean-Up System (A9281, Promega, USA), followed by sequencing.
Sanger DNA sequencing (Macrogen, Korea) was performed using the following sets of primers: CRSPR-S1R (5′-CGGAGTCGGAAACGTAGTAATG-3′), CRSPR-S2F (5′-GCCACCCTCGGCTTTAAT-3′), CRSPR-S2R (5′-GAGTAACGTGCGCTGGAA-3′), CRSPR-S3F (5′-CTGTTGCATTAGATTCGTGTTCC-3′), CRSPR-S3R (5′-CCTGTACGCCTCAGGTTTATC-3′), and CRSPR-S4F (5′-CGTATTCCGGTAGATBTDGATGG-3′). The gene map and primer positions are shown in Figure 1A. The semi-conserved CRISPR-1 locus in S. enterica subsp. *enterica *serovar Typhimurium ATCC14028 (accession number: CP034230) has been extensively studied and serves as a prominent model due to its clearly defined gene map (Figure 1B).
Structural gene map of the CRISPR-1 locus. The CRISPR locus is flanked by the EIL76_04665 and EIL76_04710 genes. To differentiate between amplification primers and sequencing primers, two lavender boxes indicate the positions of the amplification primers (A). The DRs of the CRISPR-1 locus in Salmonella strains are depicted by diamond boxes with light-blue horizontal lines, while the IGSs are represented by colored squares (B).CRISPR: clustered regularly interspaced short palindromic repeats; DRs: direct repeats; IGSs: intergenic spacers
Results
Organization and structure of the CRISPR-1 locus
Out of a total of 20 samples of S. enterica, the CRISPR-1 region of 17 (90%) was successfully amplified by PCR. Of these, 10 isolates were from humans, and seven samples were from camels. The PCR amplification showed distinct sizes of 700-800 bp PCR bands for the CRISPR-1 region in the positive samples. The CRISPR-1 region can be divided into two sub-regions: direct repeats (DRs) and IGSs of variable sequences (Figure 1B) [14]. The results revealed a wide range of CRISPR-1 region content among the isolates, regardless of whether they were from similar or different hosts. However, because the number of DRs and IGSs in the CRISPR-1 locus varies among or within species, this structure remains semi-conserved across all S. enterica strains.
Analysis of CRISPR-1 sequences
The Sanger sequencing method was used to determine the sequence of the CRISPR-1 locus in a total of 17 positive samples. The findings revealed that the sequences of the human-isolated *S. enterica *can be classified into five primary types (HSE01, HSE02, HSE03, HSE04, and HSE05), while the camel-isolated strains can be grouped into three main sequences (CSI01, CSI02, and CSI03). The arrangement and structure of the CRISPR-1 sequence in Salmonella are derived from a human isolate. The sequences of the CRISPR-1 region in human-isolated strains displayed two distinct patterns. Pattern-HA showed eight DRs and seven IGSs (HSE01 and HSE02), whereas Pattern-HB consisted of nine DRs and eight IGSs (HSE03, HSE04, and HSE05) (Figure 2).
The alignment of CRISPR-1 loci across all sets of Salmonella isolates.Diamond boxes with light-blue horizontal lines depict the DRs, while colored squares represent the IGSs. Shared IGSs are indicated by squares of the same color, whereas white squares represent unshared IGSs among different sets. Any white square labeled with the letter "K" signifies that the corresponding IGS is associated with a known sequence in the NCBI database. Conversely, white squares labeled with the letter "U" indicate that the corresponding IGS does not have a match to any known sequence in the NCBI database.CRISPR: clustered regularly interspaced short palindromic repeats; HSE: human Salmonella Enteritidis; CSI: camel Salmonella Indiana; NCBI: National Center for Biotechnology Information; DRs: direct repeats; IGSs: intergenic spacers
The IGSs of all isolates were examined to identify potential similarities and matches with previously sequenced Salmonella genomes in the National Center for Biotechnology Information (NCBI) database. Consequently, seven IGSs were discovered in HSE01 of Pattern-HA that were exactly the same as the CRISPR-1 IGS sequences already known. In contrast, HSE02 only had one IGS (IGS5) that matched a known Salmonella-CRISPR-1 sequence. Several single nucleotide polymorphisms (SNPs) were observed in the remaining IGSs at that locus. These SNPs do not appear to correspond to any known IGSs in the NCBI database. As a result, these spacers were labeled as unknown IGSs (UIGSs). The number of known IGSs and UIGSs is summarized in Table 1. Similar findings were also reported in isolates of Pattern-HB.
Following the analyses conducted on *Salmonella *isolates from humans, we also examined the CRISPR-1 region of Salmonella obtained from camels. This examination involved assessing the abundance of IGSs and comparing them to the established Salmonella CRISPR-1 sequences documented in the NCBI database. Although different from isolates sourced from humans, the three sets of CRISPR-1 loci in camel-isolated strains consistently consisted of nine DRs and eight IGSs. However, there was variability in the similarity of the three sets to the established Salmonella CRISPR-1 sequences.
Sharing IGSs is common among isolates from the same host
We analyzed IGSs and DRs in various isolates from the same host. Human-isolated strains showed a high frequency of IGS sharing. However, the arrangement or positioning of the shared IGSs appeared to be unusual. Specifically, IGS3 of the CRISPR-1 locus in set HSE01 was 97% similar to IGS2 of set HSE04. Additionally, IGS5 in set HSE02 was found to be identical to IGS6 in sets HSE01, HSE04, and HSE05 (Table 2; Figure 2). On the other hand, the CRISPR-1 locus in sets CSI02 and CSI03 of camel isolates shared the first three IGSs (IGS1, IGS2, and IGS3) (Table 3; Figure 2).
Sharing IGSs is common among isolates from different hosts
The IGSs were also analyzed to determine if there were any common IGSs among isolates from different hosts. The results showed that the CRISPR-1 locus in human isolates had specific IGSs that were also found in isolates obtained from camels (Table 4; Figure 2).
Isolated individuals from the same host also share unknown IGSs
Finally, we conducted an investigation to determine whether there were any UIGSs that were shared between isolates from humans and camels. However, our findings showed that UIGSs are specific to the host and are only shared among individuals of the same host species. Specifically, HSE03 and HSE05 shared UIGS1, while HSE03 and HSE04 shared UIGS2 and UIGS3 (Table 5). It is worth noting, though, that the isolates from camels exhibited a different pattern. The sets of CSI01 and CSI03 shared three distinct UIGSs but did not share any UIGSs with CSI02 (Table 6).
Analysis of CRISPR-1 provides different identifications from the rrnH region
The analysis of the CRISPR-1 locus contents in human and camel S. enterica isolates yielded different results. For isolates from humans, the identified content matched the CRISPR-1 loci, confirming their classification as S. enterica subsp. *enterica *serovar Enteritidis. This classification was further confirmed through 16S rRNA sequencing and the conventional O and H serotyping systems. In contrast, analysis of the rrnH region sequence using the same DNA sequencing technique revealed that the isolates from camels were identified as S. enterica subsp. *enterica *serovar Indiana. Surprisingly, the CRISPR-1 content in camel isolates showed a closer resemblance to that of S. enterica subsp. *enterica *serovar Enteritidis rather than serovar Indiana (with an identity of 89% for CSI01, 93% for CSI02, and 89% for CSI03), as documented in Table 7.
Discussion
The study has identified five distinct CRISPR-1 types among human S. enterica isolates and three among camel isolates. The presence of shared IGSs indicates the possibility of zoonotic or reverse-zoonotic transmission events. For several decades, the gold standard method for identifying different strains of Salmonella has been serotyping with specific antisera [15]. However, this method is only available in a limited number of high-quality laboratories and is applicable to a small subset of Salmonella strains [16]. As a result, researchers encounter challenges when trying to identify a wide variety of Salmonella strains that go beyond the well-known ones, such as Typhi, Paratyphi, Typhimurium, and Enteritidis [17].
There are two commonly used protocols for molecular serotyping. The first protocol examines two genes, fliC and fljB, which play a role in flagella production [18]. The second protocol focuses on genes in the rfb locus, which is responsible for the production of O-polysaccharides [19]. From a genetic standpoint, the rfb locus is approximately 8-23 kb in length, making it challenging to amplify using a conventional PCR protocol. Additionally, this system has limitations in accurately identifying the majority of Salmonella O serogroups and lacks sufficient discriminatory power between the identified Salmonella O serogroups [14].
The Salmonella CRISPR-1 locus IGSs provide a historical record that may indicate a potential reverse-zoonotic pathway. Although this study only examined a limited number of complete CRISPR-1 locus sequences, these sequences offered valuable and significant data for tracing the history of the tested isolates. There was a noticeable variation in the number of IGSs among the human-isolated strains, as well as shared IGSs across the five sets. It is uncommon for an isolate to possess CRISPR-1 with two copies of the same IGSs. Notably, the HSE01 set was the only sequence of the CRISPR-1 locus to contain two identical IGSs, specifically IGS1 and IGS2. We observed that HSE01 and HSE02 had a total of seven IGSs, while the other three sets had eight IGSs. This suggests that the isolates from these two sets may have encountered fewer hosts or predators compared to isolates exhibiting the eight-IGSs trait.
Phages are viruses that selectively infect bacterial hosts. Bacteria can evade phages through various mechanisms, such as CRISPR [20]. As a result, when bacteria encounter fewer hosts, they may have reduced exposure to phages and fewer IGSs. On the other hand, encountering a greater number of hosts would have the opposite effect [21]. Based on this hypothesis, it is suggested that sets HSE03, HSE04, and HSE05 have encountered a higher number of hosts compared to HSE01 and HSE02.
There are three identical IGSs shared by HSE01 from humans and CSI02 from camels, indicating a zoonotic origin. However, strains of S. enterica subsp. *enterica *serovars Typhimurium, Gallinarum, Dublin, Ohio, Javiana, and Pullorum share an additional IGS (IGS8) with CSI02. In fact, the majority of these strains are animal strains, particularly serovars Gallinarum and Pullorum, which are two Salmonella strains specific to poultry [22].
The strains in set HSE01 likely originated from humans and were subsequently transmitted to animals. This transmission occurred before acquiring the additional IGS8 through reverse zoonosis. The CRISPR-1 locus, which acts as the bacterial "memory cells," supports this idea. It has been observed that the CSI02 isolate from camels shares IGS7, which is the most common IGS among all sets originating from humans. This finding further strengthens the hypothesis that the isolates followed a reverse-zoonotic pathway. Various types of bacteria, viruses, and parasites, such as methicillin-resistant Staphylococcus aureus, Cryptosporidium parvum, and Ascaris lumbricoides, have been known to cause reverse zoonosis [23].
The rrnH region has limitations in identifying Salmonella serovars. The difference between the rrnH and CRISPR-1 loci profiling raises concerns about the reliability of commonly used molecular identification systems for Salmonella. Extensive research has been conducted on the effectiveness of the rrnH system in identifying Salmonella species and serovars. While the rrnH system can accurately identify Salmonella at the genus level with high sensitivity and specificity, it is important to note that the system alone cannot consistently differentiate between different Salmonella serovars [24].
The differences observed between CRISPR-1 profiling and rrnH identification may be attributed to the divergent evolutionary pressures that shape these genomic loci. It has been suggested that the CRISPR region evolves at a faster pace and captures strain-specific interactions with mobile genetic elements across various hosts [25]. Moreover, the CRISPR-Cas system serves as a key component of bacterial immunity against phages and other foreign DNA, thereby potentially rendering the CRISPR loci more indicative of the adaptive responses of strains to diverse environments and host reservoirs compared to the relatively conserved rrnH region.
Additionally, our study has revealed that the CRISPR-1 locus of the Indiana serovar contains a greater number of IGSs compared to the Enteritidis serovar. It is important to note that there are no common IGSs between these two serovars. This could potentially explain the discrepancy between the identity of rrnH and the contents of the CRISPR-1 locus, giving the CRISPR-1 locus an advantage over the rrnH system. The prevalence of *Salmonella *Enteritidis, in contrast to Salmonella Indiana strains, is higher among camels bred in the Middle East region [26,27]. Overall, it appears that the CRISPR-1 locus poses a challenge to the rrnH identification system.
This study is limited by its small sample size and potential biases in isolate selection, with a main focus on human and camel sources. This may not fully represent the genetic diversity of Salmonella. Future research should address these limitations by increasing the sample size and including isolates from a broader range of hosts and environments. By incorporating advanced molecular techniques, such as whole genome sequencing, and integrating molecular data with epidemiological and ecological information, a more comprehensive understanding of *Salmonella *transmission and evolution can be obtained. Ultimately, this would enhance control and prevention strategies.
Conclusions
Our findings have revealed the presence of several short, yet variable, IGS sequences that are specific to either human isolates or camel Salmonella isolates. However, we have also discovered that some of these variable sequences are shared between human and camel isolates. This suggests the possibility of zoonotic or reverse-zoonotic origins. Throughout the zoonotic or reverse-zoonotic journey of the Salmonella strains, each host recognizes these short, variable sequences as unwanted predators, predominantly phages. This recognition leads to the immunization of the infected ancestor against these predators. We believe that this discovery will contribute to identifying the accurate origin of Salmonella isolates worldwide, as well as providing a reliable method for serotyping different Salmonella strains.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Multiple food-animal-borne route in transmission of antibiotic-resistant Salmonella Newport to humans Front Microbiol Pan H Paudyal N Li X Fang W Yue M 23920182941065710.3389/fmicb.2018.00023 PMC 5787089 · doi ↗ · pubmed ↗
- 2The CRISPR-Cas immune system: biology, mechanisms and applications Biochimie Rath D Amlinger L Rath A Lundgren M 11912811720152586899910.1016/j.biochi.2015.03.025 · doi ↗ · pubmed ↗
- 3CRISPR-Cas systems: prokaryotes upgrade to adaptive immunity Mol Cell Barrangou R Marraffini LA 2342445420142476688710.1016/j.molcel.2014.03.011PMC 4025954 · doi ↗ · pubmed ↗
- 4Salmonella, food safety and food handling practices Foods Ehuwa O Jaiswal AK Jaiswal S 9071020213391914210.3390/foods 10050907 PMC 8143179 · doi ↗ · pubmed ↗
- 5Nutritional status of iron, vitamin B 12, folate, retinol and anemia in children 1 to 11 years old: results of the Ensanut 2012 Salud Publica Mex Villalpando S Cruz Vde L Shamah-Levy T Rebollar R Contreras-Manzano A 3723845720152654499810.21149/spm.v 57i 5.7616 · doi ↗ · pubmed ↗
- 6Potential increase in Crimean-Congo hemorrhagic fever incidence in Iraq Post Eid-al-Adha, 2023 New Microbes New Infect Hussein S Qurbani K Ahmed SK 1011755420233772750210.1016/j.nmni.2023.101175 PMC 10505661 · doi ↗ · pubmed ↗
- 7Surveillance and response strategies for zoonotic diseases: a comprehensive review Sci One Heal Sharan M Vijay D Yadav JP Bedi JS Dhaka P 10005022023
- 8Do humans spread zoonotic enteric bacteria in Antarctica?Sci Total Environ Cerdà-Cuéllar M MoréE Ayats T 19019665420193044532010.1016/j.scitotenv.2018.10.272 · doi ↗ · pubmed ↗