Physical presence of chemical synapses is necessary for turning behavior of anterograde synaptic vesicles at the branch point of PLM neurons in C. elegans
Amruta Vasudevan, Sandhya P. Koushika

TL;DR
This study shows that the physical presence of synapses is needed for synaptic vesicles to turn at branch points in C. elegans neurons.
Contribution
The study reveals that synaptic connections are essential for branch-specific transport of synaptic vesicles in neurons.
Findings
Synaptic vesicle transport at branch points depends on the presence of synapses.
Removing synaptic connections reduces branch-specific transport in PLM neurons.
This finding highlights the role of synapses in regulating cargo distribution in neurons.
Abstract
Neurons exhibit complex branched axonal morphologies in both vertebrate and invertebrate systems, and show heterogeneity in the distribution of synaptic cargo across multiple synapses. It is possible that differences in transport across multiple branches contribute to the heterogeneity in cargo distribution across multiple synapses. However, the regulation of transport at axonal branch points is not well understood. We demonstrate that branch-specific transport of synaptic vesicles is dependent on the existence of a connection between the branch and synapses. The loss of this connection causes an immediate decrease in branch-specific transport of synaptic vesicles in the PLM neuron of C. elegans .
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
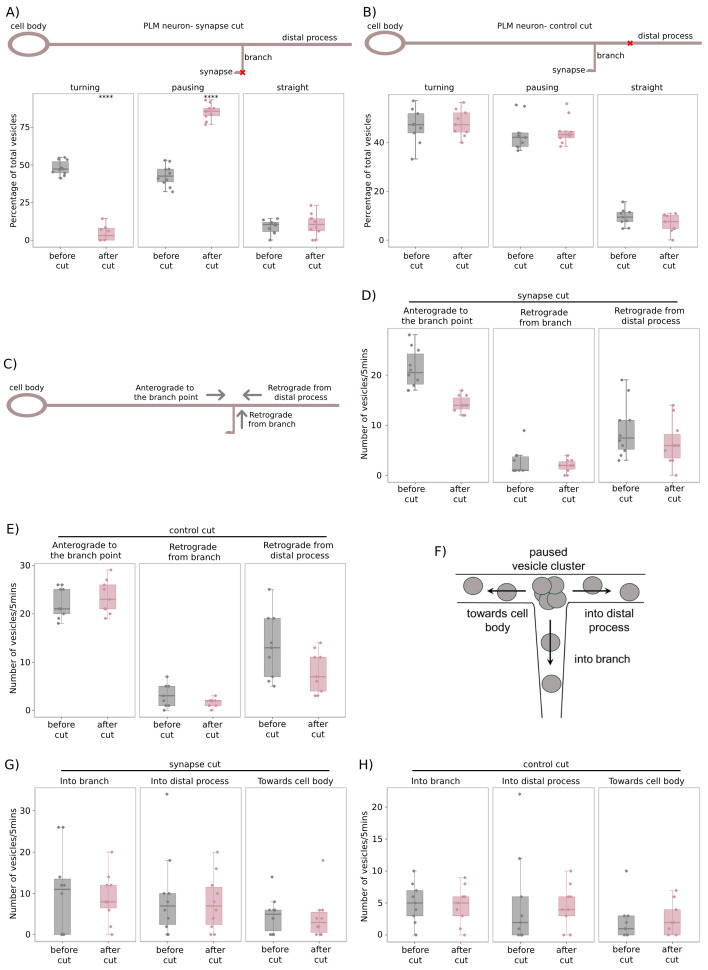 Figure 1
Figure 1|
S. No. |
Strain number |
Genotype |
Reference |
|
1 |
|
|
Bounoutas et al., 2009 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLipid Membrane Structure and Behavior · Cellular transport and secretion · Photoreceptor and optogenetics research
Description
Chemical synapses are known to play a role in remodeling of axonal branches (Alsina et al., 2001, Javaherian & Cline, 2005, Panzer et al., 2006, Ruthazer et al., 2006) . Presynaptic sites that carry a greater number of clustered synaptic vesicles, or display synaptic activity, are shown to stabilize the associated axonal branches (Ruthazer et al., 2006) . Since chemical synapses regulate the formation and structural integrity of axonal branches, they may also affect the transport of cargo along these branches. The regulation of vesicular transport and distribution by en passant chemical synapses is a field of active study, and several studies suggest that presynaptic boutons capture moving synaptic vesicles through various AZ proteins and signaling kinases, which likely oppose the mobility of synaptic vesicles locally (Wu et al., 2013, Edwards et al., 2015, Edwards et al., 2018, Lipton, Maeder, and Shen, 2018, Morrison et al., 2018) . In contrast to en passant synapses, little is known about the regulation of branch-specific transport of synaptic cargo by terminal synapses, which are located at the end of axonal branches. Several studies show that increased synaptic/growth cone activity in axonal branches is strongly correlated with increased synaptic cargo transport along those branches (Aletta & Goldberg, 1984, Schacher, 1985, Goldberg & Ambron, 1986, Goldberg & Schacher, 1987, Schacher et al., 1999, Ruthel & Hollenbeck, 2003, Lyles, Zhao, & Martin, 2006, Tymanskyj, Curran, and Ma, 2022) . However, the mechanism of this synapse-mediated regulation of branch-specific transport in vivo is not understood at all. It is unclear whether synapses regulate branch-specific cargo transport simply by stabilization of the branch or through additional mechanisms.
It has been proposed that terminal synapses send feedback signals to the neuronal cell body in order to maintain transport of synaptic cargo in the associated branch, and that the strength of this signal likely scales with the synaptic strength of the associated branch (Aletta & Goldberg, 1982, Aletta & Goldberg, 1984) . A recent study demonstrated that, following synaptogenesis, an unidentified retrograde signal from the postsynaptic neuron can trigger large-scale transcriptional and translational changes in the presynaptic neuron, which in turn increases the flux of bidirectional mitochondrial transport in the presynaptic neuron (Badal et al., 2019) . However, the source and nature of these retrograde signals is unknown. Several recent studies examine the ability of synapses to capture transiting vesicles locally (Wong et.al.,2012, Bulgari et.al.,2014). Known instances of retrograde signaling occur over shorter distances at individual synaptic boutons (Zhao & Nonet, 2000, Doi & Iwasaki, 2002, Chang et al., 2011, Hsieh et al., 2014) , and it is rare to find studies which discuss the existence of long-range synaptic signals that act from a distance to direct vesicles towards themselves. We have previously shown that branch-specific transport of synaptic vesicles is regulated by the levels of the anterograde motor UNC-104 /Kinesin-3 (Vasudevan et al., 2024) in the PLM neuron of C. elegans . It is possible that long-range retrograde signaling from synapses, if it exists, regulates branch-specific transport by acting on the motor-cargo complex and/or the cytoskeleton. Chemical synapses in the C. elegans PLM neuron play a role in stabilizing the synaptic branch, similar to studies conducted in vertebrate neurons as discussed earlier. It has been observed that removing the connection between the synaptic branch and the chemical synapse causes the synaptic branch to retract into the main process over a timescale of 24hrs (Wu et al., 2007) . Since the PLM synapse is necessary for maintenance of the synaptic branch, we investigated whether it regulates the branch-specific transport of synaptic vesicle precursors (pre-SVs).
We have previously shown that anterograde pre-SVs either turn into the synaptic branch, pause at the branch point, or go straight into the distal process of the PLM neuron (Vasudevan et al., 2024) . In this study, we used a laser ablation paradigm as described in (Figs. 1A & 1B) to investigate the role of the PLM chemical synapses in regulating branch-specific transport of pre-SVs. To visualize transport of pre-SVs, we used transgenic animals expressing GFP:: RAB-3 specifically in the touch receptor neurons (TRNs). Although this transgene overexpresses RAB-3 in TRNs, we did not observe any defects in synaptic vesicle transport due to the overexpression. The PLM synaptic branch was ablated distally, leaving most of the branch process intact while severing its connection to the cluster of chemical synapses ( Fig. 1A ). This was termed as a ‘synapse cut'. At the PLM branch point, transport of GFP::RAB-3-labelled pre-SVs was examined for 5 minutes prior to the ablation, and for 5 minutes immediately after the ‘synapse cut'. Prior to the ablation, ~40-50% of anterogradely moving pre-SVs turned into the synaptic branch, ~40% paused at the branch point, and the remaining ~10% went straight into the distal process ( Fig. 1A, Extended Figure 1B), consistent with our prior observations (Vasudevan et al., 2024) . Immediately following the ‘synapse cut', the proportion of anterograde pre-SVs turning into the synaptic branch reduced significantly to ~5-10%, a majority (~80%) paused at the branch point, while the proportion going straight into the distal process was unchanged (~10%) ( Fig. 1A, Extended Figure 1B). To ensure that the changes observed in anterograde pre-SV transport at the PLM branch point were not artefacts of laser ablation, a ‘control cut' was performed ( Fig. 1B ), where the PLM distal process was ablated at approximately the same distance from the branch point as the average length of the PLM synaptic branch (~30µm). The proportion of anterograde pre-SVs that turn into the synaptic branch, pause at the branch point, and go straight into the distal process were not significantly different from the uncut controls ( Fig. 1B, Extended Figure 1C).
In both the ‘synapse cut' and ‘control cut', we tested whether vesicular transport was altered in the ablated process by examining the number of pre-SVs transported in both the anterograde and retrograde directions along the synaptic branch and distal process (Figs. 1C & 1F). Anterograde transport in the synaptic branch comprises pre-SVs that i) turn into the branch, or ii) mobilize from the paused vesicle cluster at the branch point into the branch ( Fig. 1F ), while pre-SVs moving from the branch back to the branch point contribute to retrograde transport in the branch ( Fig. 1C ). Similarly, anterograde pre-SVs i) going straight across the branch point or ii) mobilizing from the paused vesicle cluster into the distal process contribute to anterograde transport in the distal process ( Fig. 1F ), while retrograde transport comprises pre-SVs moving from the distal process back to the branch point ( Fig. 1C ). We found that the flux of retrograde pre-SVs originating from the synaptic branch and the distal process did not differ significantly in both ablation paradigms (Figs. 1D & 1E, Movies 1,2,3,&4). Similarly, the proportion of pre-SVs mobilizing from the paused vesicle cluster to the synaptic branch and distal process did not significantly change in both ablation paradigms (Figs. 1G & 1H, Movies 1,2,3,&4). It is important to examine how the microtubule distribution is perturbed immediately after the ablation paradigms, however our observation that vesicular transport in the ablated process is not significantly altered suggests that the microtubules are not perturbed at these timescales. In summary, the loss of PLM synapses has an immediate and specific effect on the turning behavior of anterograde pre-SVs, suggesting that chemical synapses likely send out continuous signals that regulate vesicle entry into the synaptic branch. These signals are likely sensed by the axonal transport machinery of pre-SVs and responded to almost immediately (~1min), as anterogradely moving pre-SVs lose their preference for turning into the synaptic branch minutes after the connection of the synaptic branch to the chemical synapses is severed.
Methods
Maintenance of C. elegans strains
C. elegans strains were grown at 20°C on Agar plates made with Nematode Growth Medium and seeded with E. coli OP50 (Brenner, 1974). Wormbase ( http://www.wormbase.org ) was used as a reference for information on phenotypes of different strains (Sternberg et al., 2024) . Animals belonging to the young adult stage were picked for imaging and laser ablation from uncrowded, uncontaminated plates.
Time-lapse image acquisition
Live animals were anesthetized in 3mM Tetramisole in M9 buffer on 5% agarose pads. Imaging was conducted on an Olympus IX83 inverted fluorescence microscope, integrated with the Yokogawa CSU-X1 spinning disc scan head from Perkin Elmer UltraVIEW, and the Hamamatsu ImagEM C9100-13/14 EMCCD Camera, using a 488nm diode-pumped solid state laser. Software used for image acquisition was Volocity 6.0 by Perkin Elmer. Images of the PLM branch point were acquired in the Green channel at a frame rate of 5 frames per second (fps), using a UPLSAPO 100XO, 1.4 N.A. oil immersion objective. Typically, the length of the PLM process imaged spans ~70-80μm, and the length of the PLM synaptic branch imaged spans ~5-20μm depending on the orientation of the immobilized animal. Time-lapse movies were taken for a duration of 10mins.
Laser-based axotomy of the PLM synaptic branch and distal process
Laser ablation experiments were performed using a 355nm Ultraviolet range pulsed nanosecond laser (Minilite Series, Flashlamp pumped, Q-Switched, Nd:YAG) operated in low energy mode at a repetition rate of 10, with a manual trigger. Time-lapse fluorescence images of the PLM branch were acquired at 5fps, using a 100X/1.4 NA oil objective on the Olympus IX83 microscope, integrated with the Yokogawa CSU-X1-A3 spinning disc and Hamamatsu ImagEM C9100-13/14 EMCCD Camera (by Perkin Elmer), using a 488nm diode-pumped solid state laser. 5min movies were acquired prior to, and immediately after the ablation of the PLM synaptic branch/distal process.
Kymograph generation and annotation of pre-SV trajectories
All image panels used for representation and analysis of time lapse movies were generated using FIJI (ImageJ v1.52p). Experimental kymographs were generated using the ‘MultipleKymograph' plugin. Plugins were downloaded from the NIH website using the links http://www.rsbweb.nih.gov/ij/ and http://www.emblheidelberg.de/eamnet/html/bodykymograph.html.
In the kymograph, pre-SVs moving in the retrograde direction (towards the cell body), and anterograde direction (away from the cell body) appear as sloped lines, while stationary cargo appear as vertical lines. A cargo is counted as moving if it has been displaced by at least 3 pixels in successive time frames.
For each movie of the PLM branch point, a ‘branch kymograph' and ‘straight kymograph' are generated as follows:
a) Branch kymograph is generated by tracing a curved segmented line along the neuron from the branch to the pre-branch main process
b) Straight kymograph is generated by tracing a straight segmented line from the distal process to the pre-branch region along the main process
Since the branch and straight kymographs both share the same pre-branch region and only differ from each other beyond the branch point, a comparison of vesicle trajectories in both kymographs helps us identify the branch point. Consequently, a vesicle trajectory that starts in the pre-branch region and crosses the branch point in the branch kymograph is categorized as a ‘turning vesicle'. A vesicle trajectory that starts in the pre-branch region and crosses the branch point in the straight kymograph is categorized as a ‘straight vesicle'. A vesicle trajectory that starts in the pre-branch region and stops at the branch point in both the branch and straight kymographs is categorized as a ‘pausing vesicle'.
Quantification and statistical analysis
Kymographs manually annotated with pre-SV trajectories are used to identify and quantify turning, pausing, straight vesicles, and pre-SVs mobilizing from the branch point using custom ImageJ macros ‘SegmentsBranch.ijm', ‘SegmentsStraight.ijm', ‘AnalysisBranch.ijm', ‘AnalysisStraight.ijm' developed for this analysis. These codes are available at the GitHub repository link https://github.com/amruta2612/Fiji-macros . Wilcoxon rank sum test (non-parametric) with continuity correction was used on data sets that did not pass the test of normality and the distributions across different categories were varied. p-value<0.0001 is marked as ****. All statistical tests were conducted in R.
Reagents
**: **
Extended Data
Description: Raw data for figure 1 . Resource Type: Dataset. DOI: 10.22002/b2jqz-qdw65
Description: pre-SV transport at the branch point of the PLM neuron in jsIs821 young adult animals before ‘synapse cut’. Scale bar=5µm.. Resource Type: Audiovisual. DOI: 10.22002/bh4wj-t6787
Description: pre-SV transport at the branch point of the PLM neuron in jsIs821 young adult animals immediately after ‘synapse cut’. Scale bar=5µm.. Resource Type: Audiovisual. DOI: 10.22002/rz0np-0gy12
Description: pre-SV transport at the branch point of the PLM neuron in jsIs821 young adult animals before ‘control cut’. Scale bar=5µm.. Resource Type: Audiovisual. DOI: 10.22002/a6m6t-3fr28
Description: pre-SV transport at the branch point of the PLM neuron in jsIs821 young adult animals after ‘control cut’. Scale bar=5µm.. Resource Type: Audiovisual. DOI: 10.22002/942sz-7ta97
Description: Supplementary figure 1 showing kymographs of pre -SV transport before and after both ablation paradigms. Resource Type: Image. DOI: 10.22002/kjd96-1ze77
Description: Legend and caption for Extended Figure1. Resource Type: Text. DOI: 10.22002/x9mw9-bds71
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aletta JM Goldberg DJ 19821126 Rapid and precise down regulation of fast axonal transport of transmitter in an identified neuron.Science 21845750036-807591391610.1126/science.61826166182616 · doi ↗ · pubmed ↗
- 2Aletta JM Goldberg DJ 198471 Routing of transmitter and other changes in fast axonal transport after transection of one branch of the bifurcate axon of an identified neuron.J Neurosci 470270-64741800180810.1523/JNEUROSCI.04-07-01800.19846204021 PMC 6564875 · doi ↗ · pubmed ↗
- 3Alsina B Vu T Cohen-Cory S 2001111 Visualizing synapse formation in arborizing optic axons in vivo: dynamics and modulation by BDNF.Nat Neurosci 4111097-62561093110110.1038/nn 73511593233 · doi ↗ · pubmed ↗
- 4Badal KK Akhmedov K Lamoureux P Liu XA Reich A Fallahi-Sichani M Swarnkar S Miller KE Puthanveettil SV 2019115 Synapse Formation Activates a Transcriptional Program for Persistent Enhancement in the Bi-directional Transport of Mitochondria.Cell Rep 263507517.e 310.1016/j.celrep.2018.12.07330650345 PMC 6380353 · doi ↗ · pubmed ↗
- 5Bounoutas A Zheng Q Nonet ML Chalfie M 200983 mec-15 encodes an F-box protein required for touch receptor neuron mechanosensation, synapse formation and development.Genetics 18320016-673160717, 1SI 10.1534/genetics.109.10572619652181 PMC 2766320 · doi ↗ · pubmed ↗
- 6Bulgari D Zhou C Hewes RS Deitcher DL Levitan ES 2014218 Vesicle capture, not delivery, scales up neuropeptide storage in neuroendocrine terminals.Proc Natl Acad Sci U S A 11190027-84243597360110.1073/pnas.132217011124550480 PMC 3948302 · doi ↗ · pubmed ↗
- 7Chang C Hsieh YW Lesch BJ Bargmann CI Chuang CF 2011719 Microtubule-based localization of a synaptic calcium-signaling complex is required for left-right neuronal asymmetry in C. elegans.Development 138160950-19913509351810.1242/dev.06974021771813 PMC 3143565 · doi ↗ · pubmed ↗
- 8Doi M Iwasaki K 2002117 Regulation of retrograde signaling at neuromuscular junctions by the novel C 2 domain protein AEX-1.Neuron 3320896-627324925910.1016/s 0896-6273(01)00587-611804572 · doi ↗ · pubmed ↗