Interesting Cytokine Profile Caused by Clinical Strains of Pseudomonas aeruginosa MDR Carrying the exoU Gene
Nallely S. Badillo-Larios, Edgar Alejandro Turrubiartes-Martínez, Esther Layseca-Espinosa, Roberto González-Amaro, Luis Fernando Pérez-González, Perla Niño-Moreno

TL;DR
This study explores how certain drug-resistant Pseudomonas aeruginosa strains affect immune responses, focusing on cytokine profiles and virulence genes like exoU.
Contribution
The paper identifies a unique cytokine profile linked to Pseudomonas aeruginosa MDR strains carrying the exoU gene.
Findings
Strains with the exoU gene showed a distinct cytokine profile with reduced anti-inflammatory and proinflammatory cytokines.
Type III secretion system genes were present in over half of the cases, indicating their role in virulence.
Cytokine expression varied significantly based on resistance and toxicity categories of the strains.
Abstract
Pseudomonas aeruginosa is an opportunistic pathogen in HAIs with two facets: the most studied is the high rate of antimicrobial resistance, and the less explored is the long list of virulence factors it possesses. This study aimed to characterize the virulence genes carried by strains as well as the profile of cytokines related to inflammation, according to the resistance profile presented. This study aims to identify the virulence factors associated with MDR strains, particularly those resistant to carbapenems, and assess whether there is a cytokine profile that correlates with these characteristics. As methodology species were identified by classical microbiological techniques and confirmed by molecular biology, resistance levels were determined by the minimum inhibitory concentration and identification of MDR strains. Virulence factor genotyping was performed using PCR. In addition,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
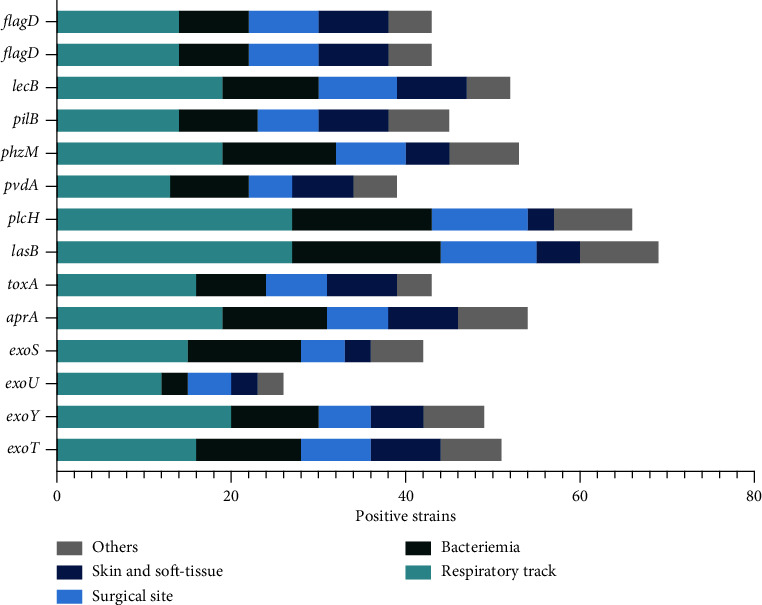 Figure 1
Figure 1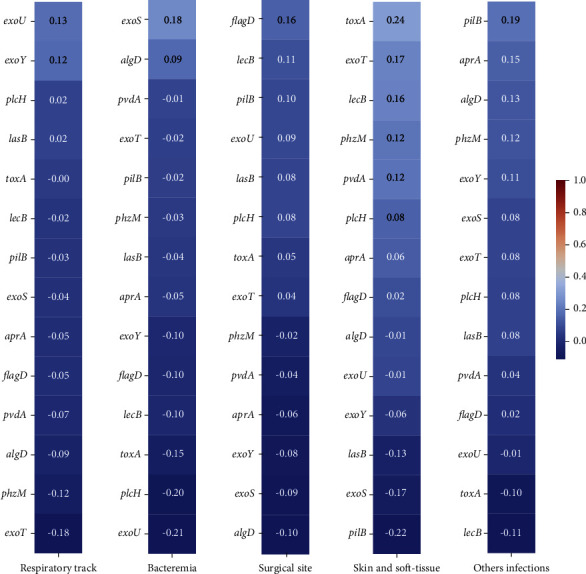 Figure 2
Figure 2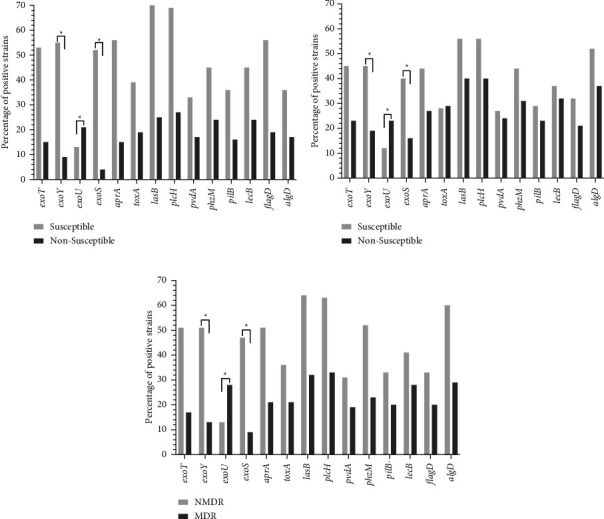 Figure 3
Figure 3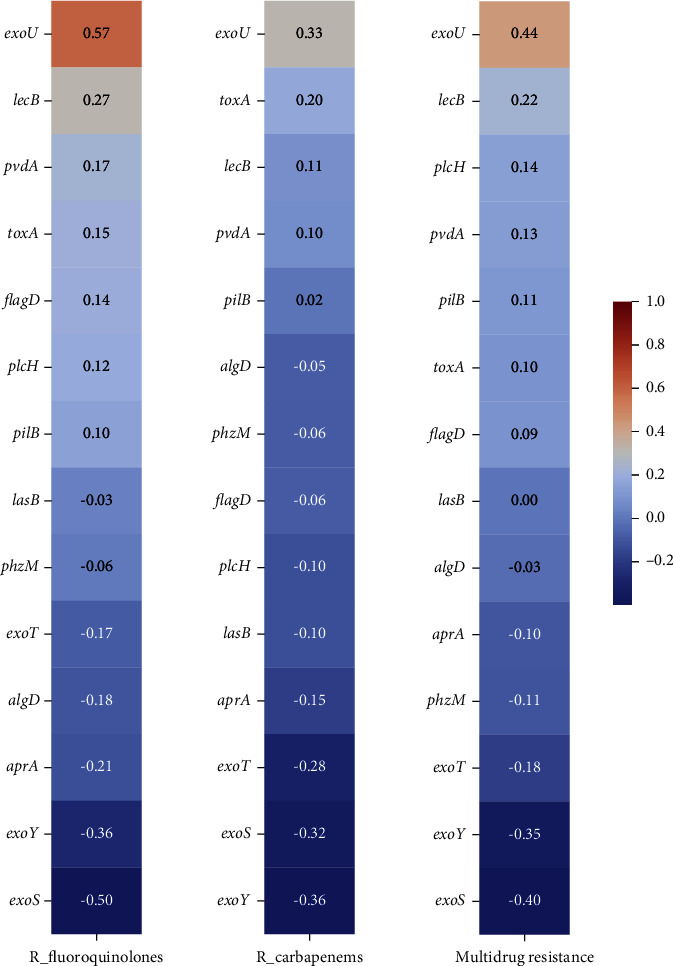 Figure 4
Figure 4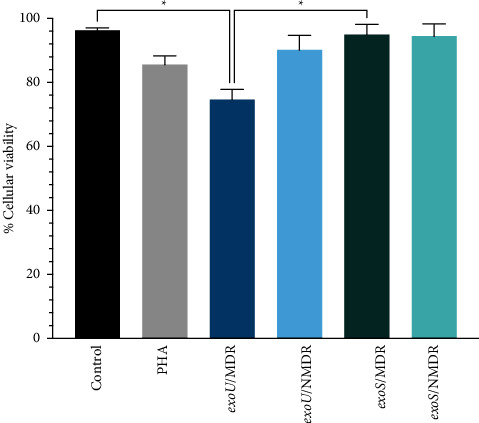 Figure 5
Figure 5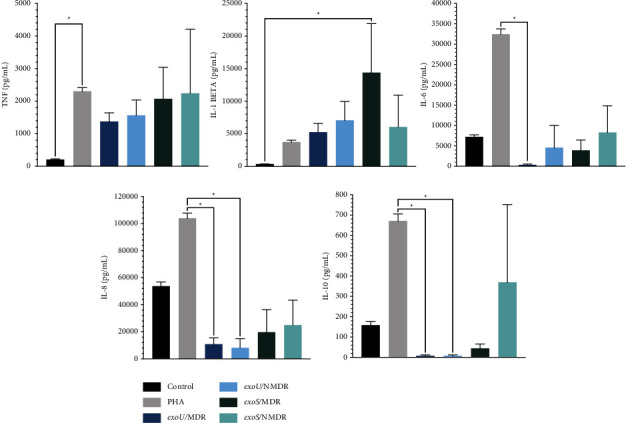 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Bacterial biofilms and quorum sensing · Bacteriophages and microbial interactions
1. Introduction
Pseudomonas aeruginosa is a Gram-negative bacterium and one of the most important pathogens in healthcare-associated infections (HAIs), causing 32,600 infections and 2,700 deaths in the United States in 2017 [1]. Classified as opportunistic pathogens involved in acute and chronic infections, the main reason for this versatility is the large list of virulence determinants. It possesses both intrinsic and acquired resistance to antibiotics and high metabolic flexibility because of its ability to use various carbon sources or electron acceptors [2]. Owing to these mechanisms, P. aeruginosa is responsible for several HCAIs, such as pneumonia, surgical infections, bacteremia, and urinary tract infections [3]. The challenge of treating an infection caused by P. aeruginosa is why the World Health Organization (WHO) has included it in the “critical” category in its priority list of pathogens that urgently require research and development of new antibiotics [4].
According to a review in 2021, it is estimated that 7.1%–7.3% of HCAIs are caused by this pathogen, and the most common infection is pneumonia, followed by surgical site infection (SSI); however, its prevalence has increased over the last decade [5]. In addition, the available data on the Mexican situation are limited; in 2022, the RHOVE (Red Hospitalaria de Vigilancia Epidemiológica, in Spanish) found P. aeruginosa to be the second most common cause of HCAIs in the country [6]. The latest information provided by the IMSS (Instituto Mexicano del Seguro Social, in Spanish) in 2016 points to P. aeruginosa as the third cause, just below Escherichia coli and Staphylococcus aureus [7]. Another report pointed out that Mexico has the highest rate of infection caused by this pathogen [8].
As mentioned above, the complexity of the infections caused by P. aeruginosa has two faces: the most studied is the high rate of antimicrobial resistance, especially resistance to carbapenems, with 17%–52% of the cases worldwide [9], and the ultimate concern is multidrug-resistant (MDR) strains, representing 9% of cases in the USA in 2018 [10].
Another issue mentioned above is the ample list of virulence factors possessed by P. aeruginosa, some of which are enzymes, exoenzymes, toxins, and the ability to grow as a planktonic community as well as in biofilms [11]. In this respect, the arsenal includes factors that are associated with the bacterial surface and those secreted, which include exotoxin A, phospholipase, alkaline protease, pyoverdine, elastases, pyocyanin, and the type three secretion system injectosome, which can deliver four different toxins in the cellular host [2, 12].
The contribution of each factor to the deterioration of the cellular host is variable, and according to Moradali, environmental stress induces differential genomic expression, forming persistent, resistant, but less virulent phenotypes in chronic illness [2]. In acute infections, virulence factors, such as flagella and pili, are vital because they are involved in motility and initial attachment. The flagellum provides swimming motility in an aqueous environment, is essential for bacterial chemotaxis, and adheres to epithelial cells by binding its flagellum to the glycolipid asialo GM1, which can also initiate the inflammatory response. Pili is an important adhesin. It is also a decisive factor for motility, but it does so through a movement called twitching. Together, these two factors also create a highly coordinated form of motility called swarming, which is useful for semisolid surfaces [12].
P. aeruginosa produces lectins that bind with specific sugars in the initial attachment of the bacteria, particularly LecB (also named PII-L), which reduces ciliary beating of the airway epithelium and is linked to biofilm formation, serving as a structural protein in the matrix [13].
P. aeruginosa owns the exotoxin A, which is an ADP-ribosyl transferase related to attachment that enters the cell and inhibits host protein synthesis by inactivating eukaryotic elongation factor 2 [14].
Furthermore, this pathogen has several proteolytic enzymes, such as elastase A and B (LasA, LasB), secreted by the type 2 secretor system (T2SS), both capable of degrading host elastin. Specifically, LecB, also named “pseudolysin,” is considered the most abundant protease capable of disrupting epithelial tight junctions [15].
Alkaline protease, also known as “aeruginosin,” secreted by the type 1 secretor system, is a metalloendopeptidase that can damage the epithelium, interfering with fibronectin and laminin [16]. Phospholipase C, a lipolytic enzyme secreted by T2SS, has hemolytic activity capable of destroying the eukaryotic membrane and destabilizing phospholipids and sphingomyelin [17].
Other important factors include the pigments produced by P. aeruginosa, such as pyoverdine, pyochelin, and pyocyanin. The first one is associated with the characteristic fluorescent green of the species and is related to the acquisition of iron, which is highly efficient but requires enormous amounts of energy in its production. In most cases, active pyochelin is produced [18]. The other pigment, pyocyanin, which is responsible for the blue-greenish color of the colonies in culture, is a phenazine and secondary metabolite related to the decline in lung function because of its inflammatory effects [19].
Another secretor system that is important to review is the type three secretion system (T3SS), a syringe-like injectosome that can deliver toxins directly into the host cell [20] and has four toxin effectors: ExoY, ExoT, ExoU, and ExoS. The first two were the most abundant and were expressed in more than 89% of the isolates. ExoY is an adenylate cyclase capable of causing cell necrosis, endothelial disruption, and irreversible actin microtubule disassembly [21]. ExoT, the most prevalent of these effectors, has two main functions: acting as an exotoxin with GTPase-activating protein (GAP) and as an adenosine diphosphate-ribosyl transferase (ADPRT), which disrupts epithelial barriers [22]. However, neither of these toxins is thought to be sufficient to establish an infection specifically in the lungs [21, 22].
The two remaining molecules are less common but highly lethal. ExoS is a GAP found in 55%–72% of clinical isolates and is associated with chronic infections, capable of producing cell death and actin cytoskeletal disruption [23]. Finally, ExoU is less frequent (28%–42% of the isolates), it has phospholipase activity with cytotoxic effects that rapidly destroy the cell membranes of mammalian cells, and some studies have found that it is more frequent in isolates with high resistance [24].
This study aimed to identify the virulence factors associated with MDR strains, particularly those resistant to carbapenems, and assess whether there is a cytokine profile that correlates with these characteristics.
2. Methods
2.1. Bacterial Isolates
A total of 75 isolates of Pseudomonas aeruginosa from infections classified as HCAIs were obtained from a tertiary hospital (HCIMP in San Luis Potosi, Mexico) between May 2018 and June 2019 before approval by the Research Committee and the Research Ethics Committee of the HCIMP. Informed consent was obtained from all participants or legal guardians.
Data collected from the medical records of the patients included sex, age, length of hospital stay, and outcome.
2.2. Bacterial Identification and Antimicrobial Susceptibility Testing
Species identification and antimicrobial susceptibility testing were performed using an automated VITEK 2 (BioMérieux SA. F-69280 Marcy l'Etoile, France). Additionally, PCR amplification was performed to confirm species identity using the primers described by Spilker et al., including primers Pa-gs for the genus and Pa-ss for the species (16S rDNA) [25].
The susceptibility profiles were confirmed using broth microdilution. Briefly, antibiotics were serially diluted 2-fold in 50 μL of Mueller-Hinton broth, mixed with 50 μL of bacteria at a density of 10^6^ colony-forming units/mL, and incubated for 18 h at 37°C. Antimicrobial susceptibility testing was performed using the following antibiotics: amikacin, cefepime, gentamicin, piperacillin/tazobactam, aztreonam, ciprofloxacin, and meropenem. The results were interpreted according to guidelines recommended by the Clinical and Laboratory Standards Institute [26]. Additionally, isolates were classified as nonmultidrug-resistant (NMDR) or MDR (defined as acquired nonsusceptibility to at least one agent in ≥3 antimicrobial categories).
2.3. DNA Extraction
Genomic DNA was extracted using a boiling method as described previously, and one loopful of fresh bacteria (grown overnight on Brain-Heart Infusion agar plates) was collected and suspended in 200 µL of sterile DNase/RNase-free water and incubated at 94 °C for 5 min and −70 °C for 5 min. The bacterial suspension was then centrifuged at 13,000 rpm at 4°C for 3 min, and the supernatant was collected and stored at −20°C.
2.4. Virulence Factor Detection
Detection of the following virulence genes was carried out by conventional PCR: type 4 fimbrial biogenesis protein (pilB), alginate (algD), alkaline protease (aprA), elastase (lasB), exoenzymes for the type three secretion system (exoS, exoU, exoT, and exoY), exotoxin A (toxA), hemolytic phospholipase C (plcH), elastase B (lasB), pyoverdine (pvdA), a lectin (lecB), a phenazine (phzM), and flagellin (flag). Primers, temperature melting (Tm), and expected amplicons of genes associated with virulence used in this study are listed in Supplementary Table 1. Amplification was carried out in a 25-μL volume containing 23 μL of PCR Master mix (DreamTaq Green PCR master mix, Thermo Scientific), 1 μL of each primer (forward and reverse), 1 μL of template DNA, and nuclease-free water.
2.5. Biofilm Production
Biofilm formation was determined using crystal violet staining. Briefly, 200 µL of TSA broth inoculated with the strains at a concentration of 0.5 on the MacFarland scale was dispensed into a 96-well polystyrene microtiter plate and incubated at 37°C for 24 h. After incubation, the contents of each well were removed by gentle tapping and washed four times with 250 µL of phosphate-buffered saline (pH 7.2) to remove free-floating bacteria. Biofilms formed by adherent bacteria were fixed by air drying. Afterward, wells were stained with crystal violet (0.1%) for 10 minutes. The excess stain was washed three times with deionized water, the plates were dried, and the dye bound to the cells was resolubilized with 200 μL of absolute methanol per well. The optical density (OD) of the stained adherent biofilm was measured using an Epoch Microplate Spectrophotometer (Winooski, VT, USA) at a wavelength of 590 nm. The negative control wells contained inoculated sterile broth, and every experiment was performed in triplicate and repeated three times. Biofilm production was interpreted according to Stepanovic et al. using the following criteria: OD > ODc = no biofilm producer, ODc < OD = 2 (ODc) = weak biofilm producer, 2 (ODc) < OD = 4 (ODc) = moderate biofilm producer, 4 (ODc) < OD = strong biofilm producer, and ODc as three standard deviations (SD) above the mean OD of the negative control [27].
2.6. Isolation of Peripheral Blood Mononuclear Cells (PBMCs)
Peripheral blood from healthy donors was collected in heparin (with informed consent), and PBMCs were obtained using a standard gradient protocol (Ficoll-Paque™ Plus, Merck KGaA, Darmstadt, Germany). Briefly, blood was diluted with PBS in a 1 : 1 ratio, and 30 mL of the mixture was layered into 10-mL Ficoll-Paque PLUS in 50-mL tubes. The tubes were centrifugated at 1000 × g for 20 minutes at 20°C without breakage. Buffy coats were collected, pooled, resuspended in PBS, and centrifuged at 400 × g for 20 min without breaking. Cells were washed twice with sterile PBS, and after centrifugation (400 × g, 30 min), a layer of PBMCs was obtained and collected, and cells were counted in a Neubauer chamber. Cell viability was evaluated using the trypan blue exclusion method (Sigma-Aldrich, USA). Cells were used only when viability was >98%. Cells were suspended in complete RPMI containing the RPMI 1640 medium supplemented with 10% fetal bovine serum (previously inactivated with heat at 56°C for 30 minutes) and 2 mM L-glutamine.
2.7. PBMC: Bacterial Coculture
Sixteen strains recovered from respiratory infections were selected and classified into four groups according to their susceptibility pattern (MDR and non-MDR) and the exotoxin genes detected (exoU and exoS). A concentration of 1 × 10^6^ CFU/mL with PBMC at a concentration of 1 × 10^6^ CFU/mL was seeded in 1-mL 24-well plates.
PBMC bacterial cocultures were incubated in complete RPMI as mentioned before at 37°C in 5% CO_2_ for 24 h. PBMCs were used as negative controls, and PBMC with 5 µg/ml of phytohemagglutinin (PHA, Sigma-Aldrich, St. Louis, MO) was used as a positive control for cell reactivity. Following incubation, 50-µL aliquots were recovered for the cell viability assay under each experimental condition. Before centrifugation at 400 × g for 5 minutes, supernatants were collected, sterile filtered (0.22 µm), and stored at −80°C for further assays.
2.8. Determination of Cytokines
The levels of a panel of cytokines (TNF-α, IL-1beta, IL-6, IL-8, and IL-10) present in the coculture supernatants were determined using a Cytometric Bead Array (CBA) Human Inflammatory Cytokines Kit (BD, Becton Dickinson, Franklin Lakes, New Jersey, U.S.A.) according to the manufacturer's instructions.
2.9. Statistical Analysis
Data were analyzed using GraphPad Prism version 9.0.0 (GraphPad Software, San Diego, California, USA, https://www.graphpad.com) and Python 3.12 (https://www.python.org) for Mac. The distribution of virulence genes among MDR and NMDR strains and their presence in carbapenem-resistant and fluoroquinolone-resistant strains were calculated using Fisher's exact test for each gene. The correlation between virulence genes and resistance rates (fluoroquinolones, carbapenems, and multidrug resistance) and the type of infection were calculated using the DataFrame.corr() method in Python. For the presence of virulence genes in biofilm production, the chi-squared test was used, as well the relationship between virulence genes and the infection involved. Differences observed in PBMC assays between groups (MDR/exoU, NMDR/exoU, MDR/exoS, and NMDR/exoS) were analyzed using a nonparametric test of variance (Kruskal–Wallis and Dunn's post-test). Statistical significance was set at p < 0.05.
3. Results
3.1. Sample Collection
As shown in Table 1, seventy-five Pseudomonas aeruginosa strains associated with HCAIs were obtained in a year, corresponding to 28 respiratory tract infections, 18 bacteremia, 11 infections from the surgical site, 9 from skin and soft tissue, and 9 from ocular and urogenital infections.
Forty-three patients were males, and thirty-two were female. Most isolates were identified in adults between 36 and 60 years of age (n = 27). The mean length of hospital stays was 42.2 days (range, 3–243 days). Most patients were discharged because of clinical improvement (n = 59), and only five deaths were associated; however, no statistical difference was found.
3.2. Antimicrobial Susceptibility
Among the antibiotics tested (Table 2), different levels of resistance were observed: 24% and 27% were nonsusceptible to amikacin and ciprofloxacin, respectively. Aztreonam showed the highest resistance (48%, n = 36), followed by meropenem (43%, n = 32). The remaining strains presented approximately 30% nonsusceptibility, and 25 strains were classified as multidrug-resistant.
3.3. Virulence Genes in Infections, Biofilm, and Antibiotics
After the PCR analyses, 16 genes were screened. The genes were classified according to the associated infection, biofilm production, susceptibility to fluoroquinolones, carbapenems, and multidrug-resistant status to determine any association or correlation.
Classified by the type of infection, the most prevalent genes were plcH, lasB, and algD, as shown inFigure 1, which were present in at least 90 percent of the strains, followed by exoT, exoY, aprA, phzM, and lecB, which were present in 60 percent of the strains. However, this difference was not statistically significant.
The correlation between the type of infection and the virulence genes shown in Figure 2, with a slight positive correlation between toxA and skin and soft-tissue infection and a negative correlation between pillB and skin and soft-tissue infection as well as between exoU and bacteremia, was detected; nevertheless, any strong positive or negative correlation was found.
As expected, most strains were medium- or high-biofilm producers (Table 3). However, three genes showed significant differences in exoY and toxA, with the majority being the stronger producers with 29 (38.7%) and (37.3%) strains, respectively. The lecB gene was mainly present in moderate producers (27 strains; 36.0%). Interestingly, some trends were observed. The pilB and exoU genes had a higher presence in medium biofilm producers, whereas the rest of the genes were present in strong producers.
Later, it explored how the genes could be distributed among strains with different resistance patterns, and in the case of resistance to carbapenems and quinolones, multidrug resistance was interesting. As shown in Figure 3, statistical differences in two of the genes belonging to SST3, exoS and exoU, were found. Additionally (Figure 4), a negative correlation between exoS and the different resistance patterns was −0.36 and −0.50 complementary, and exoU in the same resistance patterns was positive between 0.33 and 0.57. The last one was relevant because of the association between exotoxin U and strains with nonsusceptibility and MDR and its relationship with cellular destruction.
3.4. Cell Viability after Bacterial Coculture and Cytokine Production
Based on the results of the resistance and frequency of genes, four groups were formed, two bearing the exoU gene (exoU/MDR and exoU/NMDR) and two bearing the exoS gene (exoS/MDR and exoS/NMDR). The first assay involved checking the cellular viability (Figure 5), and significant differences were observed. The exoU/MDR group had higher mortality rates than the control and exoS/NMDR groups.
As shown in Figure 6, some differences were observed in cytokine assays. The presence of the strains did not affect the secretion of TNF; however, IL-6, IL-8, and IL-10 showed lower levels of secretion in the exoU/MDR group than in the positive control group, and exoU/NMDR had lower secretion of IL-8 and IL-10. Interestingly, IL-1beta was higher in the exoS/MDR group than in the control group. These findings provide interesting insights into how the presence of P. aeruginosa affects the in-vitro profile of cytokines.
4. Discussion
P. aeruginosa is an opportunistic pathogen considered a public health problem in every country. Despite the attention being focused on antibiotic resistance, this bacterium is much more complex. It possesses intrinsic resistance, a large genome capable of harboring new resistant genes, and biofilm, a barrier that confers its physical protection. In addition, this pathogen carries an extraordinary repertory of virulence factors, which this study aimed to assess.
It is difficult to scrutinize patient demographics in Mexico because there is little data about how P. aeruginosa appears in HCAIs, and the most recent descriptive analysis was made by González-Olvera and Cols. in 2019. They found a mean of five strains per month compared with six strains per month in this study, comparing the population density of the municipalities of each study, and found that San Luis Potosi has good management of nosocomial infections (NI). In addition, the predominant type of infection was on the respiratory tract (ten percent more), while for González-Olvera, most of the cases were from the urinary tract. [28–30]. Another study focusing on P. aeruginosa was conducted by Elmouaden and Cols. in 2019, with a total of 87 strains classified as NI in two years but not classified according to the type of infection [31]. Aside from these studies to date, no other study has used the same approach as the present study. However, the epidemiological data are concordant, indicating that the highest rates of infection by P. aeruginosa are in the respiratory tract [5, 9].
Something remarkable about the strains in this study is the resistance rates detected: 42.7% of the strains in this study were resistant to carbapenems and 33.3% of MDR strains. These rates are part of the increasing global problem; in this respect, many studies have been conducted, but Hojarcada and Cols. resumed this in an excellent review. In some geographical areas, the rates of MDR are between 15% and 30%; in Europe, 13.7% are resistant to at least three antimicrobial groups. In the United States, the situation is not better; MDR pathogens cause 13% of severe HIAs [32]. In Mexico, the data are limited because of the lack of a national network, and some sources suggest that resistance has diminished in the last ten years. The most recent study was conducted by Garza-González in 2019 and did not include information about the state of San Luis Potosi; they found that 27.8% resistance to meropenem was considerably lower than that found in this study [33]. It is worth mentioning that the lack of information about the country's situation has been attributed to problems such as self-prescription, low quality of generic antibiotics, the use of antibiotics in agriculture, and corruption [34].
P. aeruginosa is a pathogen with great versatility in virulence, and this study attempted to find a correlation between virulence genes, type of infection, rates of resistance, and biofilm production. In the first respect, nonstatistical differences were found, which may be because, as some authors point out, genetic differentiation or changes in gene expression occur in the transition from an acute to a chronic infection, which leads to the silencing of genes associated with acute infections when they progress to chronicity or biofilm production [35]. Furthermore, at the time of publication of this manuscript, most studies count virulence genes in all strains rather than counting them according to the type of infection; only one study with a classification similar to this one was found; Fazeli in 2014 found wound infection as the most frequent and exoS as the most prevalent gene; for this project, respiratory tract infections, plcH, and lasB genes are the most common ones, mentioning that discrepancies may be due to geographical differences [36].
Because of the behavior of the data and evidence from other studies, it was considered important to detect genes in different biofilm producers. Statistical differences were observed, and exoY and toxA were prevalent in strong producers, which is not the first report of an association between exoY and biofilm producers. Half of the Azimi strains in 2016 (n = 150) were producers and harbored exoY [37]. Similarly, in 2018, Asadpour found genes encoding exotoxin A in stronger producers, suggesting that biofilm-forming strains are more virulent, potent, and resistant [38]. According to the present study and in agreement with Bogiel T. et al., there may be a potential relationship between virulence factors and biofilm production rates, suggesting that biofilm producers are more virulent [39]. In a completely different scenario, Passos da Silva et al. recently demonstrated that lecB coding for a glycoprotein can bind the exopolysaccharide PSL and stabilize the biofilm matrix, which explains why this gene is present in 69.3% (n = 52) of their strains [13] and 45.3% (n = 49) in this study.
A classic scenario for bacteria is that they are considered simple life forms with a small space in their genome, and some hypothesize that a strain cannot harbor a plethora of virulence factors while containing many resistance-related genes. However, this has changed in recent years; the relationship between antibiotic-resistant genes and virulence factors might follow a Darwinian model, resulting in the emergence of virulent and resistant clones, and precisely, the P. aeruginosa large genome is an excellent candidate to acquire both characteristics [40, 41]. Since 2008, twenty-seven papers have reported a relationship between the exoU gene and resistance to carbapenems, including reports of community and nosocomial infections [42].
The evidence in this study suggests a possible relationship between virulence genes and resistance rates; nevertheless, there are very few studies looking for this relationship, and some have not found significant differences [37]. Although this study revealed differences, it found strains that harbored the exoU gene, were resistant to fluoroquinolones and carbapenems, and were considered MDR. These findings are consistent with the data of Takata et al. and Subedi et al. in 2018, who found that the exoU-positive genotype is more frequent in nonsusceptible fluoroquinolones, carbapenems, and MDR strains [24, 43]. In 2019, Horna found that strains resistant to carbapenems and fluoroquinolones and classified as MDR also harbored the exoU gene [44]. Kainuma et al. found the same phenomenon and suggested that exoU carriers have a greater potential to spread within hospital units, which, if true, represents a major risk [45].
The final aim was to explore a possible relationship among the genes of T3SS, specifically exoS and exoU, multidrug resistance, and the production of cytokines (TNF, IL-1beta, IL-6, IL-8, and IL-10). The first approach was to determine the effects of these bacteria on cellular viability. As shown in Figure 5, exoU/MDR strains presented higher mortality than the other strains owing to their potent phospholipase activity, which causes necroptosis and cell lysis [46], and the exoS groups showed a higher survival rate. Some evidence points out that exoS may dampen the immune response, provoking cell death by apoptosis, which is slower than necrosis [47]. Information about how P. aeruginosa can affect the pattern of cytokine expression is limited; one of the most accurate studies was made by Christopher H. The Moody group related the effect of a recombinant exoS with the mRNA of cytokines from PBMC. They found a decrease in IL-10 and an increase in TNF, IL-6, IL-8, and IL-1beta [48, 49]. They worked with recombinant proteins. The present study attempted to evaluate whether these results are transferable to clinical strains, which is partial, and observed a significant increase in IL-1beta with a significant difference in the MDR group. A possible explanation is provided by Grandjean et al., who found that ExoS can convert prointerleukin (IL-1beta) into mature IL-1beta [50]. IL-10 (NMDR group) and TNF (MDR, NMDR group) levels also increased, but the difference was not statistically significant, which could be an indirect effect of ExoS through NF-κB [20].
For the exoU group, the first finding was higher levels of IL-1beta, although nonsignificant, than those in the control group. Hardy et al. in 2022 showed that ExoU has a noncytolytic function through the transitory activation of caspase-1 and proIL-1beta. Moreover, interesting findings emerged: IL-6, IL-8, and IL-10 levels decreased, but only in MDR strains. At the time of publication of this manuscript, there were three papers on the relationship between exoU and IL-8; one of them addressed the relationship with IL-6, and the other mentioned the effect of this gene on IL-10. All these studies reported that strains with exoU caused an increase in the cytokines tested. This discrepancy may be due to the articles being published before 2012 and the use of different cell lines. Today, considering the results of Hardy, a second look is taken at the relationship between these exotoxins and their effects on cytokines [51–54]. However, a limitation in this study is that these results are the product of an in-vitro model, and the results should be interpreted with caution due to the complexity of the study and the possibility that there are intracellular cytokines that could cross the membranes of the cells undergoing cell death and affect the cytokine pattern.
In conclusion, the appearance of the exoU virulence gene in MDR strains has increased in recent years, which is concerning because it is an indicator of strains with a more aggressive and difficult-to-treat infectious capacity. More importantly, the fact that these strains present a different cytokine profile than expected represents the possible existence of unknown mechanisms of how these strains interact with host cells, which may represent a benefit for the progression of the infection.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cdc Pseudomonas aeruginosa infection HAI 2021 https://www.cdc.gov/hai/organisms/pseudomonas.html
- 2Moradali M. F. Ghods S. Rehm B. H. A. Pseudomonas aeruginosa lifestyle: a paradigm for adaptation, survival, and persistence Frontiers in Cellular and Infection Microbiology 20177 p. 3910.3389/fcimb.2017.000392-s 2.0-85014708437 PMC 531013228261568 · doi ↗ · pubmed ↗
- 3Azam M. W. Khan A. U. Updates on the pathogenicity status of Pseudomonas aeruginosa Drug Discovery Today 201924135035910.1016/j.drudis.2018.07.0032-s 2.0-8505087886630036575 · doi ↗ · pubmed ↗
- 4Shrivastava S. R. Shrivastava P. S. Ramasamy J. World health organization releases global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics Journal of Medical Society 2018321 p. 7610.4103/jms.jms_25_172-s 2.0-85048729976 · doi ↗
- 5Reynolds D. Kollef M. The epidemiology and pathogenesis and treatment of Pseudomonas aeruginosa infections: an update Drugs 202181182117213110.1007/s 40265-021-01635-634743315 PMC 8572145 · doi ↗ · pubmed ↗
- 6Hospital Epidemiological Surveillance Network General office of epidemiology. Bulletin infections associated with health care (iaas) hospital epidemiological surveillance network (RHOVE) 20222022 www.gob.mx/salud
- 7Arias-Flores R. Rosado-Quiab U. Vargas-Valerio A. Grajales-Muñiz C. Microorganisms responsible for nosocomial infections in the Mexican Institute of social security Revista medica del Instituto Mexicano del Seguro Social 2016541202426820194 · pubmed ↗
- 8Huerta-Gutiérrez R. Braga L. Camacho-Ortiz A. One-day point prevalence of healthcare-associated infections and antimicrobial use in four countries in Latin America International Journal of Infectious Diseases 20198615716610.1016/j.ijid.2019.06.0162-s 2.0-8507021623731229613 · doi ↗ · pubmed ↗