Two new species of Paraphlomis (Lamiales, Lamiaceae) from limestone karsts in Guangdong Province, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
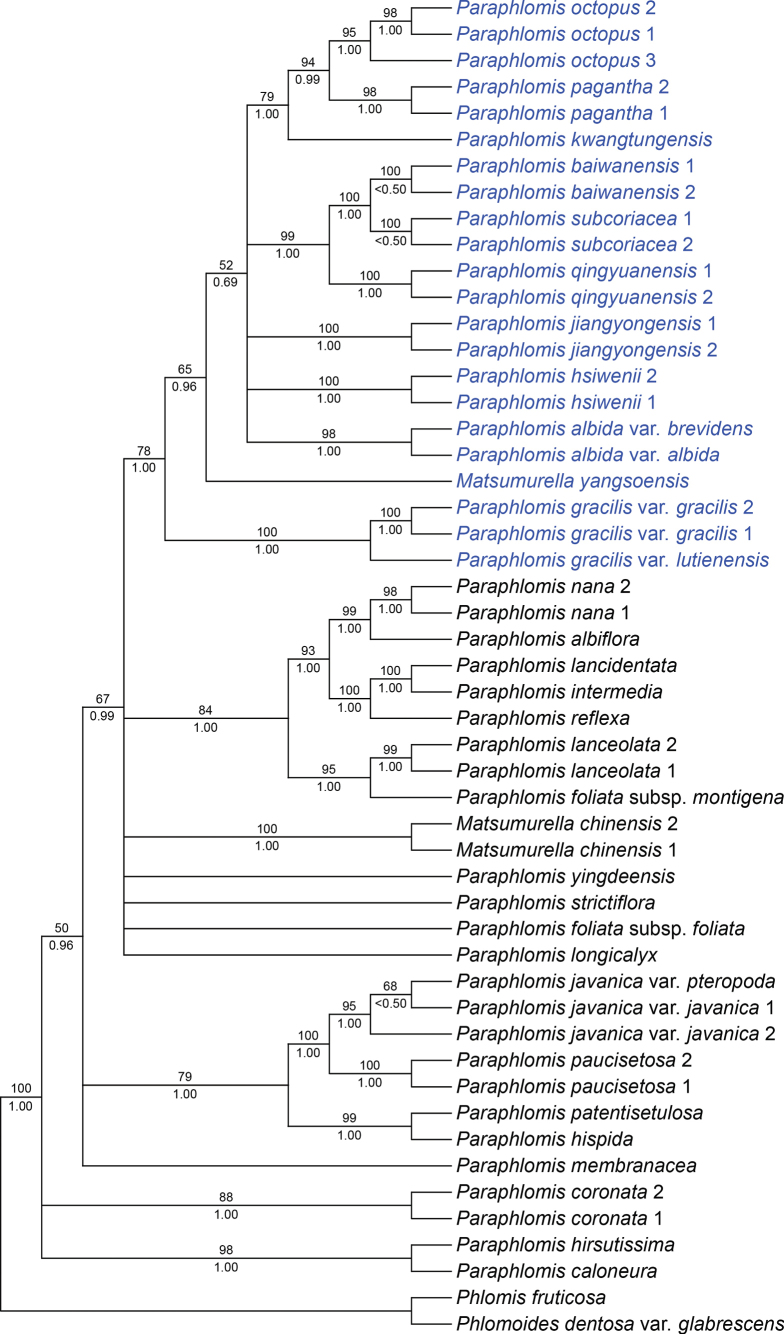 Figure 1
Figure 1 Figure 2
Figure 2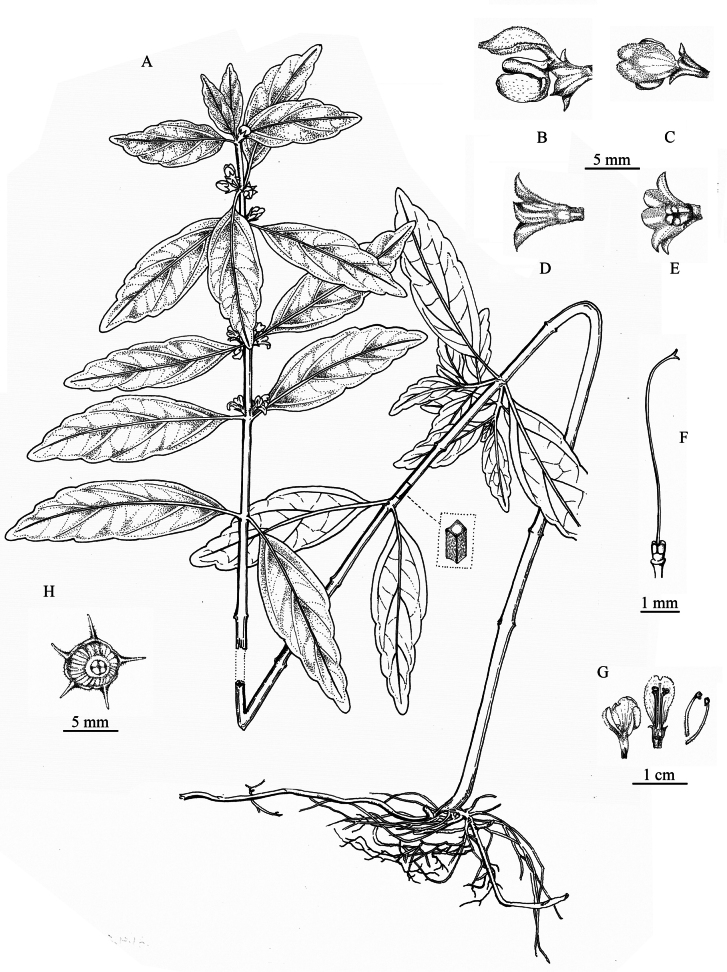 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5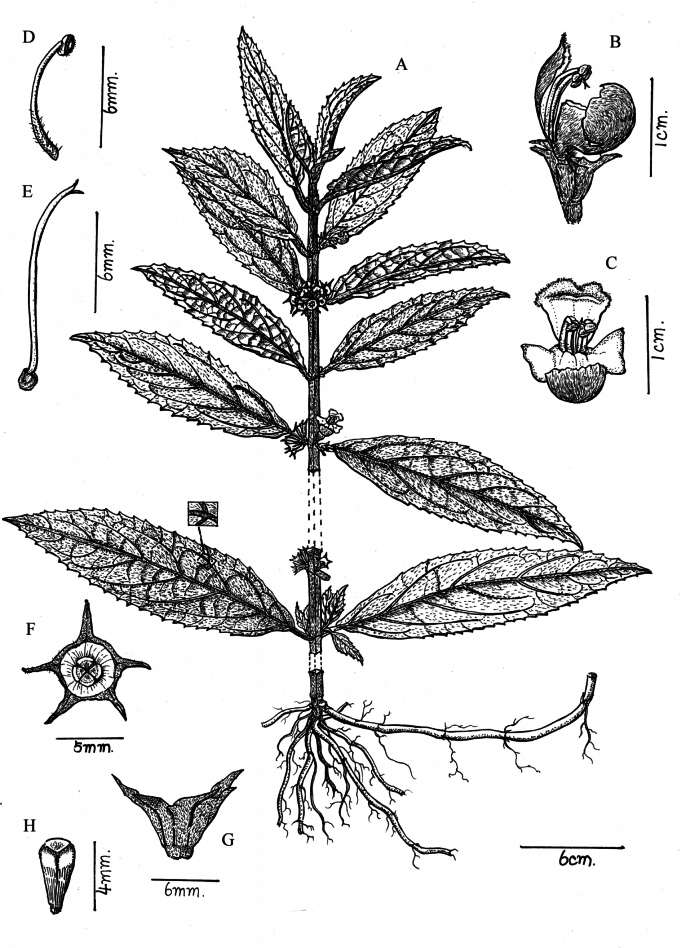 Figure 6
Figure 6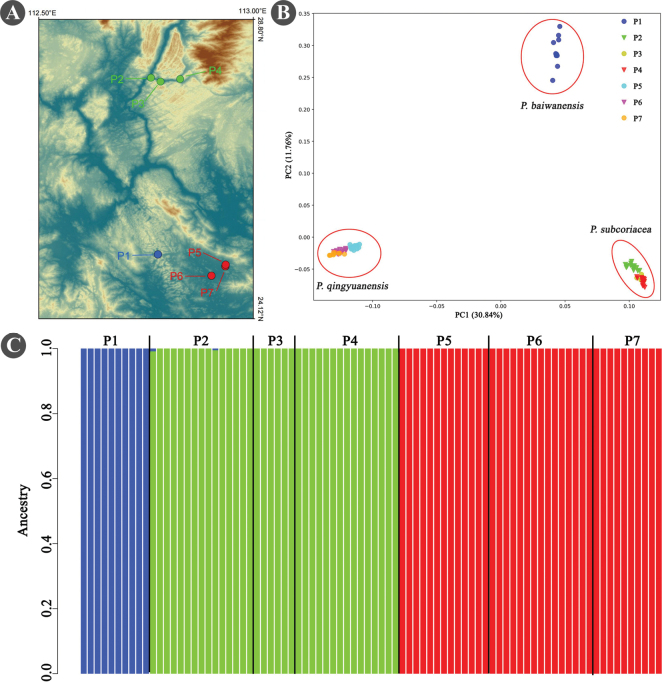 Figure 7
Figure 7| Characters |
|
|
|
|
|---|---|---|---|---|
|
| 50–90 cm, slender, 1.4–3.1 mm in diam | 50–80 cm tall, erect, stout, 3.6–5.3 mm in diam | ca. 60 cm, ca. 3.6 mm in diam | ca. 40 cm, ca. 2.5 mm in diam |
|
| much branched | unbranched or 2–3 branched | unbranched or 2–3 branched | unbranched |
|
| 0.4–1.6 cm | 0.6–1.3 cm | 1.0–1.3 cm | 0.3–0.6 cm |
|
| 2.7–9.5 cm long, 1.2–2.5 cm wide | 12.5–18.7 cm long, 2.6–4.8 cm wide | 7–15 cm long, 1.5–3.2 cm wide | 5–8 cm long, 2.4–3.4 cm wide |
|
| cuneate, not decurrent | cuneate or abruptly obtuse, not decurrent | attenuate or abruptly obtuse, not decurrent | obtuse to rounded, not decurrent |
|
| papery | leathery | thin leathery | thin leathery |
|
| 2–4 (-5) pairs | 5–7 (-8) pairs | 5–6 pairs | 4–5 pairs |
|
| subtruncate, extended into wings from veins | subtruncate, extended into wings from veins | subtruncate, extended into wings from veins | triangular, conspicuously extended into wings from veins |
|
| upper lip yellow, lower lip red with purple spots | white | white or purple-white | unknown |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and Fungal Species Descriptions · Phytochemistry and Biological Activities · Plant Diversity and Evolution
Introduction
Paraphlomis (Prain) Prain is a member of the tribe Paraphlomideae Bendiksby (Lamiaceae, Lamioideae) (Bendiksby et al. 2011; Li et al. 2016; Zhao et al. 2021). Most species of Paraphlomis are distributed in southern China, with several species occurring in the Himalayas, Korea and Southeast Asia (Li and Hedge 1994; Wu and Li 1997; Ko et al. 2014; Chen et al. 2021). Previous molecular phylogenetic studies of Paraphlomis revealed that the genus was not monophyletic, because species of Matsumurella were recovered within it (Chen et al. 2021; Chen et al. 2022b; Guo et al. 2023). In general, Paraphlomis is characterized by its herbaceous habit, actinomorphic calyx with five lobes less than half as long as the tube, corolla 2-lipped (1/3) with hairy upper lip but hardly bearded along the margin, included stamens and an apically truncate ovary (Wu and Li 1977; Bendiksby et al. 2011; Ko et al. 2014; Chen et al. 2021).
As currently circumscribed, a total of 37 species and seven varieties are recognised within Paraphlomis (Li and Hedge 1994; Chen et al. 2022a, 2022b; Yuan et al. 2022; Guo et al. 2023; Yan et al. 2023). China is the distribution center of Paraphlomis, with 23 species documented in the Flora of China (Li and Hedge 1994). In recent years, a number of new species and infraspecies of Paraphlomis were reported in China, including P.javanicavar.pteropoda D. Fang & K.J. Yan and P.javanicavar.angustifoliaf.albinervia D. Fang & K.J. Yan (Yan and Fang 2009); P.breviflora B.Y. Ding, Y.L. Xu & Z.H. Chen (Ding et al. 2019); P.kuankuoshuiensis R.B. Zhang, D. Tan & C.B. Ma (Zhang et al. 2020); P.jiangyongensis X.L. Yu & A. Liu and P.coronata (Vaniot) Y.P. Chen & C.L. Xiang (Chen et al. 2021); P.nana Y.P. Chen, C. Xiong & C.L. Xiang (Chen et al. 2022c); P.longicalyx Y.P. Chen & C.L. Xiang (Chen et al. 2022a); P.hsiwenii Y.P.Chen & XiongLi (Chen et al. 2022b); P.strictiflora J.C.Yuan, B.Chen & C.L.Xiang (Yuan et al. 2022); P.jinggangshanensis Boufford, W.B. Liao & W.Y. Zhao (Zhao et al. 2022); P.yingdeensis W.Y.Zhao, Y.Q.Li & Q.Fan (Guo et al. 2023), P.caloneura K.J.Yan, Y.P.Chen & Y.Feng Huang (Yan et al. 2023).
During a botanical expedition of the limestone area in Qingyuan city, Guangdong Province in 2023, we discovered two unknown species of Paraphlomis. The calyx teeth of the two unknown species extended into wings from veins, which are consistent with the characteristics of Paraphlomisser.Subcoriaceae C.Y. Wu et H.W. Li (Li 1965; Wu and Li 1977). However, their morphological characters differ from the two known two species of the series, P.subcoriacea C. Y. Wu ex H. W. Li and P.brevifolia C. Y. Wu & H. W. Li. Thus, we suspected both of them were undescribed species. After careful field observations, morphological comparisons with other species of Paraphlomis, and molecular phylogenetic studies, we confirmed that they were new species and named them as P.qingyuanensis W.Y. Zhao, R.M. Wu & Q. Fan and P.baiwanensis W.Y. Zhao, Y.P. Chen & Q. Fan.
Materials and methods
Morphological study
The flowering and fruiting plants of the two new species were examined in the field from August to December in 2023 and compared with herbarium specimens deposited in IBSC, KUN, GCMI and SYS (herbarium acronyms follow Thiers 2023). The two putative new species were most similar to Paraphlomissubcoriacea and P.brevifolia. We have carried out several field work trips to the collection site of the type specimens of these two species from December 2023 to April 2024. Unfortunately, we did not find P.brevifolia in the field due to a lack of detailed collection site information (Li 1965). Therefore, the morphological features contrasting with those of P.brevifolia were based on its type specimens (IBSC0005124). All morphological characteristics were measured using dissecting microscopes.
Phylogenetic analyses
The nuclear ribosomal internal and external transcribed spacers (ITS and ETS) were used for reconstructing the phylogeny of the suspected new species and related taxa based on previous study (Chen et al. 2021; Zhao et al. 2022). Most sequences were downloaded from GenBank, except for the two nuclear ribosomal DNA (nrDNA) sequences of the two new species and Paraphlomissubcoriacea, which were newly sequenced in the present study. Genomic DNA of the suspected new species was extracted from silica-gel-dried leaves using the modified 2× CTAB procedure of Doyle and Doyle (1987). The ITS and ETS sequences were amplified with primer pairs 17SE/26SE (Sun et al. 1994) and ETSB/IGS (Beardsley and Olmstead 2002), respectively, with PCR amplification and sequencing following Chen et al. (2016). A total of 49 accessions representing 30 species and four varieties/subspecies of Paraphlomis and two species of Matsumurella were sampled in the phylogenetic study. Phlomoidesbracteosa (Royle ex Benth.) Kamelin & Makhm. and Phlomisfruticosa Sieber ex C. Presl were selected as the outgroups. The GenBank accession numbers are listed in Appendix 1.
Nucleotide sequences were aligned using MAFFT 7 (Katoh and Standley 2013). After removing aligned columns with more than 70% missing data using Phyx (Brown et al. 2017), the two nrDNA regions were concatenated for phylogenetic reconstruction. The phylogenetic relationships were assessed using the Bayesian inference (BI) and maximum likelihood (ML) methods, and both were implemented on the online server Cyberinfrastructure for Phylogenetic Research Science (CIPRES) Gateway (http://www.phylo.org/; Miller et al. 2010). The BI analysis was carried out using MrBayes 3.2.7a (Ronquist et al. 2012) and the ML analysis using RAxML-HPC2 (Stamatakis 2014), with detailed settings following that of Chen et al. (2021). The resulting BI tree with posterior probabilities (PP) and best-scoring ML tree with bootstrap support (BS) values were visualized and annotated using TreeGraph 2 (Stover and Müller 2010).
Population genetic structure analyses
To investigate the phylogenetic relationship between the two newly discovered species and their close relatives, we further conducted an analysis of their population genetic structure. A total of 84 individuals were sampled from seven populations of the two putative Paraphlomis new species and its close relative P.subcoriacea (Appendix 2). The fresh leaves were dried and stored with silica gel, and then sent to JieRui BioScience Co. Ltd. (Guangzhou, China) for DNA extraction, ddRAD-seq library preparation, and Illumina sequencing. The produced raw data was processed with the software Stacks 2.55 (Catchen et al. 2013). The procedure “process_radtags” was used to demultiplex RAD tags, “denovo_map.pl” to process all the fastq files, and “populations” to filter the data by setting “--min-maf 0.05 --max-obs-het 0.7 -R 0.8 –write-random-snp –vcf”. The produced vcf file was transformed to ped file using bed file using software vcftools 0.1.16 (Danecek et al. 2011), and to bed file using the software Plink v1.90 (Chang et al. 2015). The produced bed file was used to perform Bayesian cluster analysis with the software ADMIXTURE v1.3.0 (Alexander et al. 2009), in which the number of groups (K) was set from 1–6 and the optimal K was determined by the minimum value of cross-validation error (CV). Principal coordinate analysis (PCA) was performed with Plink, and python script were used to draw the scatter diagram.
Results and discussion
The combined nuclear data set was 1211 bp (773 bp for ITS, 438 bp for ETS) in length, including 347 variable sites (165 for ITS, 182 for ETS) and 200 parsimony-informative characters (87 for ITS, 113 for ETS). The resulting phylogenetic tree of Paraphlomis in this study was similar to that in previous studies (Chen et al. 2021; Zhao et al. 2022). The two accessions of P.baiwanensis grouped together (Fig. 1: BS = 100%/PP < 0.50) and this new species was sister to P.subcoriacea (Fig. 1: BS = 100%/PP = 1.00). The P.baiwanensis-P.subcoriacea clade was further sister to another new species, P.qingyuanensis (Fig. 1: BS = 99%/PP = 1.00). All three species were nested within the previously suggested “Clade III” by Chen et al. (2021) (Fig. 1: BS = 78%/PP = 1.00). Species of “Clade III” (marked in blue in Fig. 1) are characterized by hairy nutlets/ovaries (Chen et al. 2021; 2022b). The nutlets of both new species are sparsely hispid and densely glandular at apex (Figs 2M, 5I, 6H), which confirmed the significance of nutlet morphology for the infrageneric classification of Paraphlomis (Chen et al. 2021).
The best-scoring maximum likelihood tree of Paraphlomis inferred from concatenated nrDNA (ETS and ITS) dataset. Support values ≥ 50% BS or 0.50 PP are displayed above and below the branches, respectively. Multiple accessions of the same species are numbered according to Appendix 1.
ParaphlomisqingyuanensisA habitat B plants C individual D inflorescences with unopened flowers E opening flowers F leaves G stolons and roots H lateral view of flower I upper lip of flower J dissected corolla and stamens K infructescence L frontal and lateral view of calyx M dissected calyx and seeds. (Photographs: A, B by Qin-Dai Xiong; C–G by Wan-Yi Zhao).
The close relationships among the two new species and P.subcoriacea are also supported by morphological evidence. In morphology, the two putative new species are most similar to Paraphlomisbrevifolia C. Y. Wu et H. W. Li and P.subcoriacea C. Y. Wu ex H. W. Li. They share the features such as conspicuously extended calyx teeth, and the fact that they are also growing in a limestone habitat. A comparison of their morphological features is presented in Table 1 and Fig. 4.
The putative new species Paraphlomisqingyuanensis differs from P.brevifolia by its slender and much branched stem (vs. unbranched) (Figs 2C, 4A), leaves without glandular (vs. abaxially golden glandular) (Table 1). Furthermore, the reticulate veins and pilose indumentum on leaves of P.brevifolia are more obvious (Fig. 4B). P.qingyuanensis could also be easily distinguished from P.subcoriacea by its papery leaves (vs. thin leathery), fewer leaf veins, 2–4 pairs (vs. 5–6 pairs), and smaller leaf size (Table 1, Fig. 4A, D).
Paraphlomisbaiwanensis is most similar to P.subcoriacea. They share such features as leaves lanceolate, 5–7 lateral vein pairs, and leaf base shape (Table 1). However, P.baiwanensis could be easily distinguished from the latter by its stout stem (diam. 3.6–5.3 mm vs. ca. 3.6 mm), leathery leaves (vs. thin leathery), and larger leaves size (12.5–18.7 cm × 2.6–4.8 cm vs. 7–15 cm × 1.5–3.2 cm) (Table 1, Fig. 4C, D). Furthermore, the distribution areas of the two species are separated by a distance of approximately 45 km, exhibiting complete non-overlap (Fig. 7A). The results of population genetic analysis also confirmed significant differences in their genetic structure (Fig. 7B).
Bayesian cluster analysis showed lowest CV value as K = 3, each of the three species P.baiwanensis (P1), P.subcoriacea (P2–4), and P.qingyuanensis (P5–7) possesses a unique gene pool, and no gene admixture is observed in any individuals (Fig. 7B). PCA analysis reveals similar results in which the 84 individuals are assigned into three groups and individuals of the same species are clustered into the same group (Fig. 7C). These results showed that individuals of the three species can be separated clearly from each other based on genomic data, strongly supported their species status and no obvious gene flow was observed among the three species.
Taxonomic treatment
Paraphlomis
qingyuanensis
Taxon classificationPlantaeLamialesLamiaceae
W.Y.Zhao, R.M.Wu & Q.Fan sp. nov.
972DB679-0048-5138-81CE-40A3870F8F9B
urn:lsid:ipni.org:names:77344362-1
Type.
China. Guangdong Province: Yingde City, Huanghua Town, near Hegushi, on the limestone valley, 24°13'N, 112°56'E, alt. 135 m, 7 September 2023, Xiong Qin-Dai ZWY-3793 (holotype: SYS00236954! isotypes: KUN, SYS00236955!, SYS00236968!, SYS00236969!).
Diagnosis.
Paraphlomisqingyuanensis is morphologically similar to P.brevifolia and P.subcoriacea, but differs from the latter two species by its slender and much branched stem, papery leaves, inconspicuous reticulate veining, and fewer leaf veins.
Description.
Herbs perennial, 50–90 cm tall; stoloniferous, with sparsely villous. Stems erect or decumbent, slender and much branched, 4-angled, diameter 1.4–3.1 mm, densely retrorse pilose. Leaves opposite; lamina ovate to oblong, papery, 2.7–9.5 cm long, 1.2–2.5 cm wide, apex obtuse, base cuneate, margin crenate-serrate; adaxially green, abaxially light green, sparsely pilose on both sides, more densely on viens; lateral veins 2–4 (-5)-paired; petiole 0.4–1.6 cm long, densely pilose. Verticillasters born in upper leaf axils, cymes 1–4-flowered; bracteoles inconspicuous, ca. 0.5 mm long, early deciduous; pedicels ca. 1 mm long, sparsely pilose. Calyx light green, obconical, 4.5–5 mm long, outside with sparsely pilose and white glandular, glabrous inside, conspicuously 10-veined; teeth 5, inconspicuous so that calyx mouth appears subtruncate, extended into wings from veins as 1.5–2.0 mm long. Corolla 1.0–1.2 cm long; tube ca. 0.3 cm long, ca. 1 mm in diam., straight, inside of throat dark purple, with pubescent annulate; 2-lipped, villous outside, upper lip oblong, erect and galeate, apex undulate or bilobate, ca. 8 mm long, ca. 4 mm wide; lower lip reflexed, ca. 4.5 mm long, 3-lobed, medium lobe largest, suborbicular, ca. 5 mm long, ca. 4.5 mm wide, apex emarginate, white, dotted with purplish-red spots, lateral lobes ovate, ca. 3.5 mm long, ca. 3 mm wide, apex obtuse, yellow. Stamens 4, inserted above middle and upper of corolla tube, straight, included, filaments flat, sparsely puberulent-villous, anther cells 2, ovoid, glabrous. Style filiform, included, glabrous, apex subequally 2-lobed, ca. 1 cm long. Ovary 4-loculed, truncate at apex. Nutlets triquetrous-oblong, ca. 2.5 mm long, apex sparsely hispid and glandular.
Line drawing of ParaphlomisqingyuanensisA plant B lateral view of flower C upper lip of flower D lateral view of calyx E inner view of calyx F style G dissected corolla and stamens H front view of calyx (Drawn by Zhong-Jin Wang).
Distribution, habitat and conservation status.
Currently, only three populations of Paraphlomisqingyuanensis were found in Huanghua Town of Yingde City, Qingyuan City in Guangdong Province (Fig. 7A). These populations were located in the subtropical monsoon climate region, in a large area of karst landform. The distribution area of P.qingyuanensis is extremely fragmented, and it is not within a protected area. Human activity, such as forestry production and tourism, have a negative effect on population regeneration. Thus, P.qingyuanensis is here suggested to be endangered (EN) according to IUCN categories guidelines B2(a, b(iii)) (AOO < 500 km^2^, number of locations <5, and habitat affected by human activities) (IUCN Standards and Petitions Subcommittee 2022).
Morphological comparison of Paraphlomisqingyuanensis, P.brevifolia, P.baiwanensis, and P.subcoriacea. ParaphlomisqingyuanensisA plant of P.qingyuanensisB plant of P.brevifoliaC plant of P.baiwanensisD plant of P.subcoriacea (Photographs by Wan-Yi Zhao; photographs of P.brevifolia from isotype W. Chen 84411, IBSC0005124).
Phenology.
Flowering was observed from August to December, and fruiting from September to December.
Etymology.
The specific epithet “qingyuanensis” is derived from the type locality of the new species, i.e. Qingyaun City in Guangdong Province, China.
Additional specimens examined.
Paraphlomisqingyuanensis (paratypes): China. Guangdong Province: Qingyuan City, Yingde City, Huanghua Town, near Hegushi, on the limestone valley, 24°13'N, 112°56'E, alt. 132 m, 6 September 2023, Qin-Dai Xiong QYK-HH-156 (SYS); Yingde city, Huanghua town, Hegushi, 24°13'35.7"N, 112°56'08.97"E, alt. 114 m, 16 December 2023, Qiang Fan 20255 (SYS); Yingde city, Huanghua town, near Hegushi, 24°13'22.71"N, 112°56'05.71"E, alt. 159 m, 26 December 2023, Qiang Fan & Qin-Dai Xiong QYK-HH-1904 (SYS); Yingde city, Huanghua town, Huanghua park, 24°12'03.46"N, 112°54'10.05"E, alt. 205 m, 25 December 2023, Qiang Fan & Qin-Dai Xiong QYK-HH-1882 (SYS).
Paraphlomis
baiwanensis
Taxon classificationPlantaeLamialesLamiaceae
W.Y.Zhao, Y.P.Chen & Q.Fan sp. nov.
6A6801CD-587F-52BB-93A7-284B893883D7
urn:lsid:ipni.org:names:77344363-1
Type.
China. Guangdong Province: Qingyuan city, Qingxin district, Baiwan town, Hecang village, on the limestone cliff, 24°15'09.40"N, 112°46'47.32"E, alt. 310 m, 15 December 2023, Qiang Fan 20251 (holotype: SYS00236952!; isotypes: KUN, SYS00236953!).
Diagnosis.
Paraphlomisbaiwanensis is morphologically similar to P.subcoriacea, but differs from the latter by its stout stem, larger and leathery leaves, and larger flower.
Description.
Herbs perennial, 50–80 cm tall; stoloniferous, with villous indumentum. Stems erect, stout, unbranched or 2–3 branched, 4-angled, diameter 3.6–5.3 mm, with densely retrorse pilose hairs. Leaves opposite; lamina long ovate to lanceolate, leathery, 12.5–18.7 cm long, 2.6–4.8 cm wide, apex acuminate, base cuneate or abruptly obtuse, margin serrulate; adaxially dark green, with densely pilose, abaxially light green with densely brown glandular, sparsely pilose, more densely on veins; lateral veins 5–7 (-8)-paired, raised abaxially and deeply impressed adaxially, anastomosing at leaf margin; petiole 0.6–1.3 cm long, densely pilose. Verticillasters borne in upper leaf axils, cymes (2-) 5–9-flowered; bracteoles inconspicuous, early deciduous; pedicels ca. 1.2–2.0 mm long, densely pilose. Calyx light green, obconical, 5.2–5.7 mm long, glabrous inside, outside with densely retrorse pilose, conspicuously 10-veined; calyx teeth 5, inconspicuous, throat appearing subtruncate, calyx veins extended into wings as 2.5–3.0 mm long. Corolla 1.5–2.1 cm long, white; tube ca. 7 mm long, ca. 2 mm in diam., straight, inside of throat with pubescent annulate; 2-lipped, villous outside, upper lip oblong, galeate, apex undulate, ca. 8–10 mm long, ca. 3.8–5.5 mm wide; lower lip reflexed, ca. 7 mm long, 3-lobed, medium lobe largest, suborbicular, ca. 6.5 mm long, ca. 6 mm wide, apex emarginate, lateral lobes ovate, ca. 4.5 mm long, ca. 3.2 mm wide, apex obtuse. Stamens 4, inserted above middle and upper of corolla tube, straight, included, filaments flat, 7–9 mm long, sparsely puberulent-villous, anther cells 2, ovoid, glabrous. Style filiform, included, glabrous, apex subequally 2-lobed, ca. 1 cm long. Ovary 4-loculed, truncate at apex. Nutlets triquetrous-oblong, ca. 3.8 mm long, apex sparsely hispid.
ParaphlomisbaiwanensisA plant B young branch and leaves C flowering branch D leaves E lateral view of flower F front view of flower G lateral view of calyx H front view of calyx I seeds (Photographs: A, B, D, G, I by Wan-Yi Zhao; C by Qin-Dai Xiong; E-F, H by Qiang Fan).
Distribution, habitat and conservation status.
Paraphlomisbaiwanensis is currently known to occur only in Baiwan town, Guangdong in one population numbering less than one hundred individuals. It was observed to grow on limestone cliffs at altitudes about 300 m. Its known population was located in Qingxin Baiwan Provincial Nature Reserve of Guangdong which is well-protected. More field investigations are needed to determine its wild distribution. Therefore, the Protection level of Paraphlomisbaiwanensis was suggested as Data Deficient (DD) based on the IUCN Red List Criteria (IUCN Standards and Petitions Subcommittee 2022).
Line drawing of ParaphlomisbaiwanensisA plant B lateral view of a flower C frontal view of corolla D stamen E style F front view of calyx G lateral view of calyx tube H seed (Drawn by Rong-En Wu).
Phenology.
Flowering was observed from June to September, and fruiting from August to December.
Etymology.
The specific epithet “baiwanensis” is derived from the type locality of the new species, i.e. Qingxin Baiwan Provincial Nature Reserve of Guangdong, Qingyuan, China.
Geographic distribution and population genetic structure of Paraphlomisbaiwanensis (P1), P.subcoriacea (P2–4), and P.qingyuanensis (P5–7) based on Admixture analysis A geographic distribution of the seven populations used for analysis B two-dimensional clustering of genetic variation from seven populations obtained via principal component analysis C Populations genetic structure result based on Admixture analysis.
Additional specimens examined.
Paraphlomisbaiwanensis (paratypes): China. Guangdong Province: Qingyuan city, Qingxin district, Baiwan town, Hecang village, on the limestone cliff, 24°15'09.40"N, 112°46'47.32"E, alt. 310 m, 23 August 2023,Yi-Wen Liu QYK-BW-1470 (SYS).
Paraphlomisbrevifolia: China. Guangxi Province: Luocheng City, 1939, Wen Chen 84411 (IBSC0005124, isotype).
Paraphlomissubcoriacea: China. Guangdong Province: Qingyuan City, Yangshan City, Chengjia country, Wuyuan village, Baizhushan, 20 June 1956, Liang Deng 1572 (IBSC0005130, isotype); Yangshan City, Dalang Town, Chakeng village, 7 August 1936, Liang Deng 263 (IBK00059958; IBSC0585118); Yangshan City, Lingbei Town, Xiatianxia, 24°39'48.99"N, 112°45'41.73"E, alt. 295 m, 21 December 2023, Qiang Fan 20257 (SYS); Yangshan City, Lingbei Town, Yanzidong, 24°39'16.92"N, 112°46'59.95"E, alt. 156 m, 27 December 2023, Qiang Fan & Qin-Dai Xiong QYK-LB-1920 (SYS); Yangshan City, Chengjia Town, Baizhuling (collection site of type specimen), 24°39'39.56"N, 112°49'45.69"E, alt. 387 m, 27 December 2023, Qiang Fan & Qin-Dai Xiong QYK-CJ-1911 (SYS).
Supplementary Material
XML Treatment for Paraphlomis qingyuanensis
XML Treatment for Paraphlomis baiwanensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alexander DH Novembre J Lange K (2009) Fast model-based estimation of ancestry in unrelated individuals.Genome Research 19(9): 1655–1664. 10.1101/gr.094052.10919648217 PMC 2752134 · doi ↗ · pubmed ↗
- 2Beardsley PM Olmstead RG (2002) Redefining Phrymaceae: The placement of Mimulus, tribe Mimuleae, and Phryma.American Journal of Botany 89(7): 1093–1102. 10.3732/ajb.89.7.109321665709 · doi ↗ · pubmed ↗
- 3Bendiksby M Thorbek L Scheen AC Lindqvist C Ryding O (2011) An updated phylogeny and classification of Lamiaceae subfamily Lamioideae.Taxon 60(2): 471–484. 10.1002/tax.602015 · doi ↗
- 4Brown JW Walker JF Smith SA (2017) Phyx: Phylogenetic tools for unix.Bioinformatics 33(12): 1886–1888. 10.1093/bioinformatics/btx 06328174903 PMC 5870855 · doi ↗ · pubmed ↗
- 5Catchen J Hohenlohe PA Bassham S Amores A Cresko WA (2013) Stacks: An analysis tool set for population genomics.Molecular Ecology 22(11): 3124–3140. 10.1111/mec.1235423701397 PMC 3936987 · doi ↗ · pubmed ↗
- 6Chang CC Chow CC Tellier LCAM Vattikuti S Purcell SM Lee JJ (2015) Second-generation PLINK: rising to the challenge of larger and richer datasets. Gigascience 4(1): s 13742-015-0047-8. 10.1186/s 13742-015-0047-8PMC 434219325722852 · doi ↗ · pubmed ↗
- 7Chen YP Drew BT Li B Soltis DE Soltis PS Xiang CL (2016) Resolving the phylogenetic position of Ombrocharis (Lamiaceae), with reference to the molecular phylogeny of tribe Elsholtzieae.Taxon 65(1): 123–136. 10.12705/651.8 · doi ↗
- 8Chen YP Liu A Yu XL Xiang CL (2021) A preliminary phylogenetic study of Paraphlomis (Lamiaceae) based on molecular and morphological evidence.Plant Diversity 43(3): 206–215. 10.1016/j.pld.2021.03.00234195505 PMC 8233522 · doi ↗ · pubmed ↗