Identification of epithelial ciliated cells as the non‐endothelial source of mouse endocan
Yoann Stordeur, Sophie Salomé Desnoulez, Patricia de Nadai, Joanne Balsamelli, Saliha Ait Yahia, Anne Tsicopoulos, Philippe Lassalle, Alexandre Gaudet

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
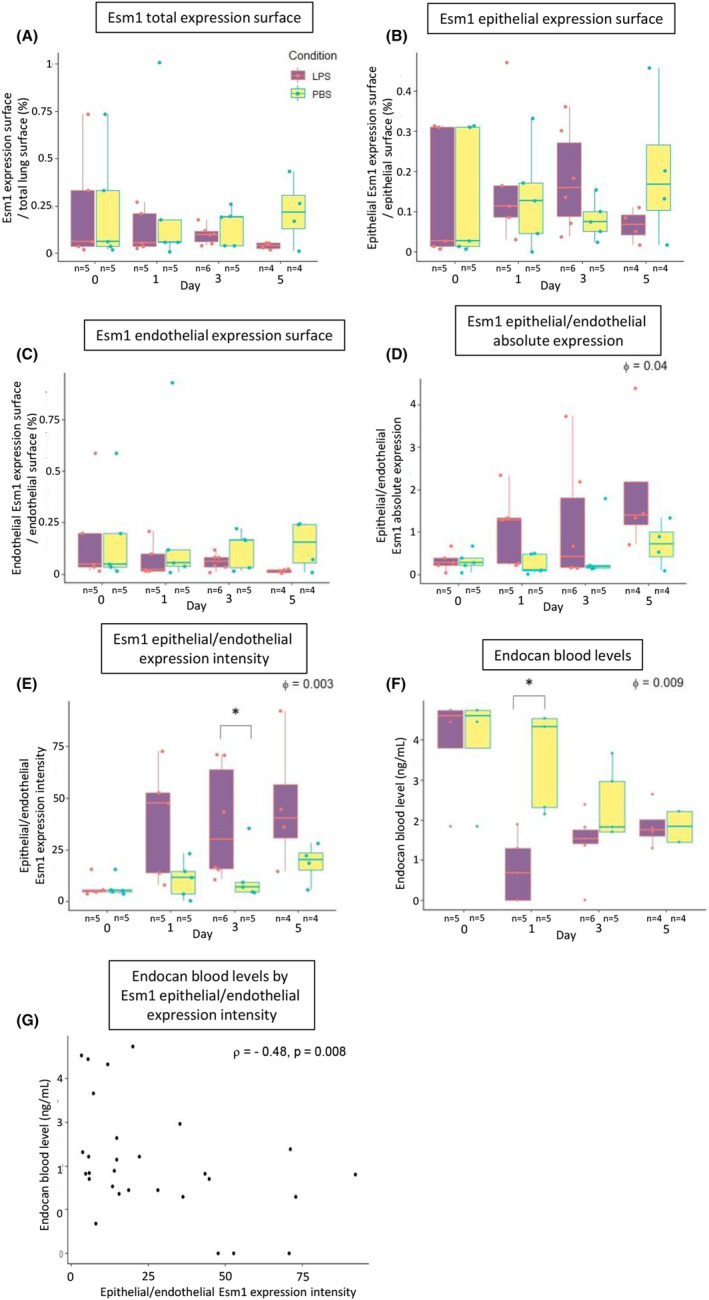 Figure 1
Figure 1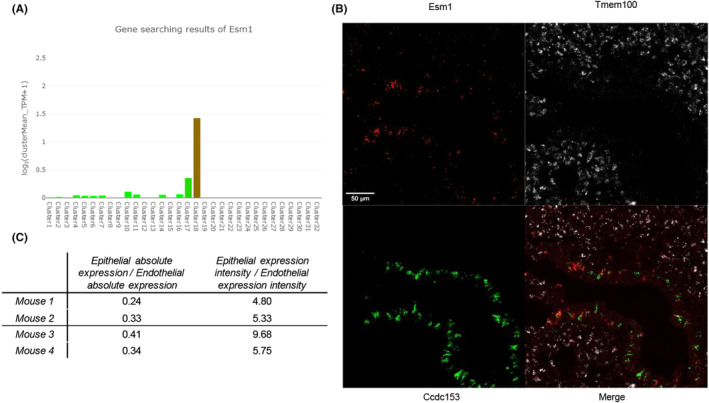 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInflammation biomarkers and pathways · Respiratory Support and Mechanisms · Neonatal Respiratory Health Research
Endocan is a circulating proteoglycan mostly secreted by pulmonary endothelial cells in human.1 Because of its immunomodulatory properties resulting from the inhibition of leukocyte diapedesis, endocan has been proposed as a candidate molecule for the treatment of acute respiratory distress syndrome (ARDS).2, 3 In vivo studies investigating the biological effects of endocan mostly rely on mouse models.4 However, major differences have been reported between the physiological properties of mouse and human endocan.4 One critical difference relies in the cellular populations responsible for the pulmonary expression of endocan. Moreover, it is still unknown whether measurement of blood endocan routinely performed for the daily clinical practice is a good surrogate for lung expression variations occurring in acute lung inflammation.
To address these questions, we first analysed online mouse transcriptomic databases (GSE108097—Mouse Cell Atlas)5 to identify candidate cell subsets involved in the pulmonary synthesis of mouse endocan through the expression of its mRNA Esm1.
We then used the RNAScope™ Multiplex Fluorescent Assay v2 (Bio‐Techne, Rennes, France), on formalin fixed paraffin embedded lung sections from 8 to 12 weeks old C57BL/6 male mice to detect the mRNA expression of Esm1, Ccdc153, a marker of epithelial ciliated cells, and Tmem100, a marker of endothelial cells.
Anaesthetised C57BL/6 mice were exposed to a tracheal instillation of either 2 mg/kg of lipopolysaccharide (LPS) diluted in 50 μL of phosphate‐buffered saline (PBS) to trigger a non‐lethal acute lung injury (ALI) model, or 50 μL of PBS without LPS in control mice, which were considered as the sham group, following a previously published model.2 We quantified the pulmonary expression of Esm1 on whole sections at days 1, 3 and 5 following tracheal instillation of LPS using the RNAScope™ Multiplex Fluorescent Assay v2. Ccdc153 and Tmem100 were respectively used as epithelial and endothelial markers to characterize the source of expression of Esm1.
All images were acquired using a Zeiss LSM 880 confocal microscope, and processed using Imaris (Oxford instruments, Abingdon‐on‐Thames, UK). Magnification was set at 63X for the identification of Esm1 pulmonary cellular source and at 40X for the quantification of Esm1 expression in our ALI model.
Endocan blood levels were measured by ELISA (DIYEK M1®, Lunginnov, Lille, France) on serum after two centrifugations at 1500g.
Our analysis of publicly available transcriptomic data indicated that, in mouse, Esm1 pulmonary expression was predominantly found in ciliated epithelial cells (Figure 1A, detailed list of clusters is shown in Table S1). We report the results regarding the identification and quantification by RNAscope™ of Esm1 expression in non‐endothelial cells in Figure 1B,C. In naïve mice, we found an intense Esm1 signal colocalizing with Ccdc153, along with a less intense Esm1 signal colocalizing with Tmem100 (Figure 1B). Quantification of Esm1 in whole lung sections showed that endothelial cells are responsible for a majority of the total amount of expression of Esm1, with an epithelial/endothelial absolute expression ratio found between 0.24 and 0.41. However, the expression intensity, defined as absolute expression divided by surface area for each cell type, was higher for epithelial ciliated cells with an epithelial/endothelial expression intensity ratio found between 4.8 and 9.68 (Figure 1C). In other words, the expression of Esm1 in epithelial areas was 4.8–9.68 times more intense than in endothelial areas.
In our ALI model, the kinetics of Esm1 expression surface showed no significant variation over time in LPS‐challenged mice versus control mice. However, we found a trend for a decrease from days 1 to 5 for the overall tissue (ϕ = 0.14) (Figure 2A–C). The epithelial/endothelial absolute expression and expression intensity ratios significantly increased over time in LPS‐challenged mice vs control mice (ϕ = 0.04 and ϕ = 0.003, respectively), especially at day 3 (p = 0.03) (Figure 2D,E). In other words, following LPS administration, the share of expression of Esm1 taking its source from epithelial cells increased over time. In addition, these results suggest that this increase in the epithelial share of Esm1 expression seemed notably explained by a stronger intensity of Esm1 expression in epithelial areas relatively to endothelial areas. After LPS administration, blood levels of endocan reached their lowest level at day 1, at 0.68 ng/mL (0–1.93), then rose back to levels observed in the control group at day 5 (ϕ = 0.009) (Figure 2F). The kinetics of blood endocan and Esm1 expression in lung had divergent patterns, as pulmonary expression still remained low by day 5. There was a negative correlation between endocan blood levels and the epithelial/endothelial intensity of expression ratio (ρ = −0.48, p = 0.008). As reflected by this result, the highest relative intensity of Esm1 expression in epithelial areas and lowest relative intensity of Esm1 expression in endothelial areas were observed in mice with the lowest levels of blood endocan (Figure 2G).
Our results show that part of Esm1 is intensely produced by the epithelial ciliated cells in mouse lungs, in accordance with the transcriptomic data. These results contrast with those of a study exploring the renal expression of endocan in mice, which identified endothelial cells as the quasi‐exclusive source of this molecule.6
In naïve mice, epithelial absolute expression is lower than the endothelial one, though a down‐regulation of the endothelial fraction of Esm1 expression seems to be observed in ALI until day 5. These variations show some dissimilarities with the apparent increase observed in endocan blood levels from day 1 to 5 following LPS administration, implying a possible influence of extra‐pulmonary sources of expression of endocan.
The present results point out to the fact that we consistently observe trough different studies a major difference between the kinetics of endocan in human and mice. Indeed, in human, the blood levels of endocan are increased under pro‐inflammatory conditions, like sepsis, pneumonia or ARDS.7 On the other hand, in mice, we report in two previous publications from our group a decrease in blood levels of endocan following tracheal LPS‐induced ALI.2, 3 Therefore, the results shown in the present study seem consistent with our previous results in mice, though the discrepancy with the kinetics observed in human remains unexplained.
The type of ALI that was obtained through our model can be questioned as well. Indeed, by injecting directly LPS into the airways is likely to induce a model of primary ALI/ARDS. Therefore, it may not seem surprising that epithelial cells are more activated. There is still a possibility that in other types of secondary ARDS, the expression of endocan could be different and preferentially in the endothelial cells.
These results suggest that mouse models of ALI need refinement to be used for further studies regarding endocan. However, our results are exploratory and further studies to confirm them by other approaches, in vitro or in vivo, seem necessary.
AUTHOR CONTRIBUTIONS
Yoann Stordeur: Conceptualization (supporting); formal analysis (lead); investigation (lead); methodology (supporting); writing – original draft (lead). Sophie Salomé Desnoulez: Formal analysis (supporting); writing – review and editing (supporting). Patricia de Nadai: Investigation (supporting); writing – review and editing (supporting). Joanne Balsamelli: Investigation (supporting). Saliha Ait Yahia: Investigation (supporting). Anne Tsicopoulos: Formal analysis (supporting); writing – review and editing (supporting). Philippe Lassalle: Writing – review and editing (supporting). Alexandre Gaudet: Conceptualization (lead); formal analysis (supporting); methodology (lead); supervision (lead); validation (lead); writing – review and editing (lead).
CONFLICT OF INTEREST STATEMENT
The authors have disclosed that they do not have any conflicts of interest.
Supporting information
Table S1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lassalle P , Molet S , Janin A , et al. ESM‐1 is a novel human endothelial cell‐specific molecule expressed in lung and regulated by cytokines. J Biol Chem. 1996;271(34):20458‐20464.8702785 10.1074/jbc.271.34.20458 · doi ↗ · pubmed ↗
- 2Gaudet A , Portier L , Prin M , et al. Endocan regulates acute lung inflammation through control of leukocyte diapedesis. J Appl Physiol (1985). 2019;127(3):668‐678.31295063 10.1152/japplphysiol.00337.2019 · doi ↗ · pubmed ↗
- 3Hureau M , Portier L , Prin M , et al. Evaluation of endocan as a treatment for acute inflammatory respiratory failure. Cells. 2023;12(2):257.36672192 10.3390/cells 12020257 PMC 9857156 · doi ↗ · pubmed ↗
- 4De Freitas CN , Lassalle P . Highlight on mouse Endocan. Circ Res. 2015;116(8):e 69‐70. doi:10.1161/CIRCRESAHA.115.306353 25858072 · doi ↗ · pubmed ↗
- 5Han X , Wang R , Zhou Y , et al. Mapping the mouse cell atlas by microwell‐Seq. Cell. 2018;172(5):1091‐1107.e 17.29474909 10.1016/j.cell.2018.02.001 · doi ↗ · pubmed ↗
- 6Gaudet A , Zheng X , Kambham N , Bhalla V . Esm‐1 mediates transcriptional polarization associated with diabetic kidney disease. Am J Physiol Renal Physiol. 2024;326:F 1016‐F 1031.38601985 10.1152/ajprenal.00419.2023 PMC 11386982 · doi ↗ · pubmed ↗
- 7De Freitas CN , Gaudet A , Portier L , Tsicopoulos A , Mathieu D , Lassalle P . Endocan, sepsis, pneumonia, and acute respiratory distress syndrome. Crit Care. 2018;22(1):280.30367649 10.1186/s 13054-018-2222-7PMC 6204032 · doi ↗ · pubmed ↗