Molecular and morphological evidence supports the resurrection of Chrysospleniumguangxiense H.G.Ye & Gui C.Zhang (Saxifragaceae)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
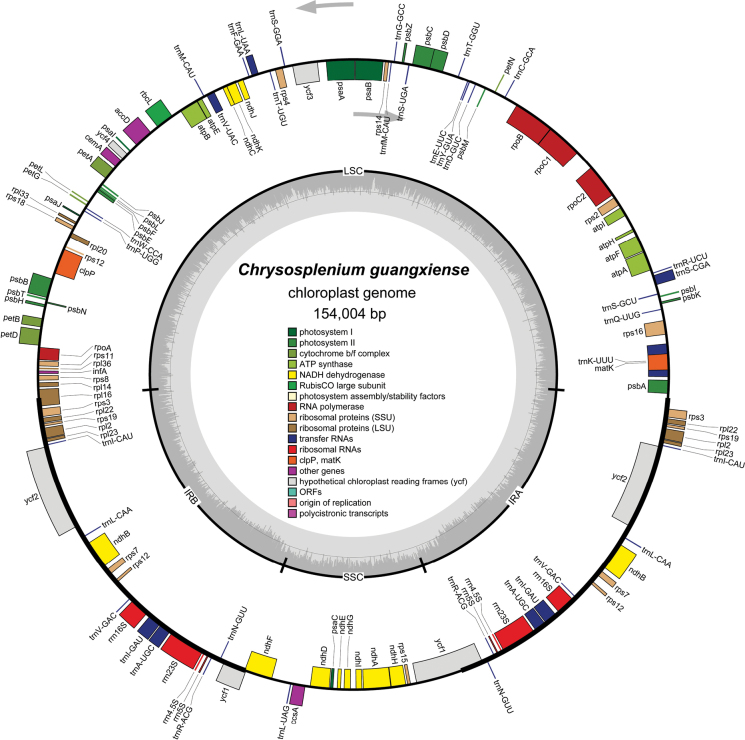 Figure 1
Figure 1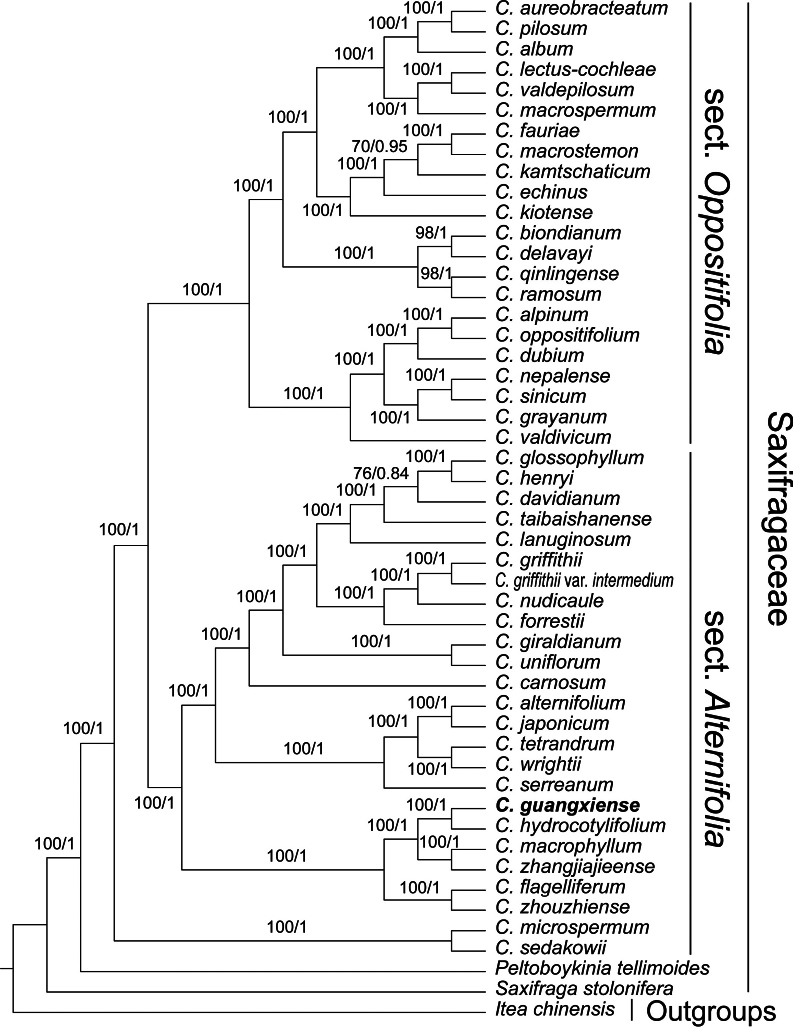 Figure 2
Figure 2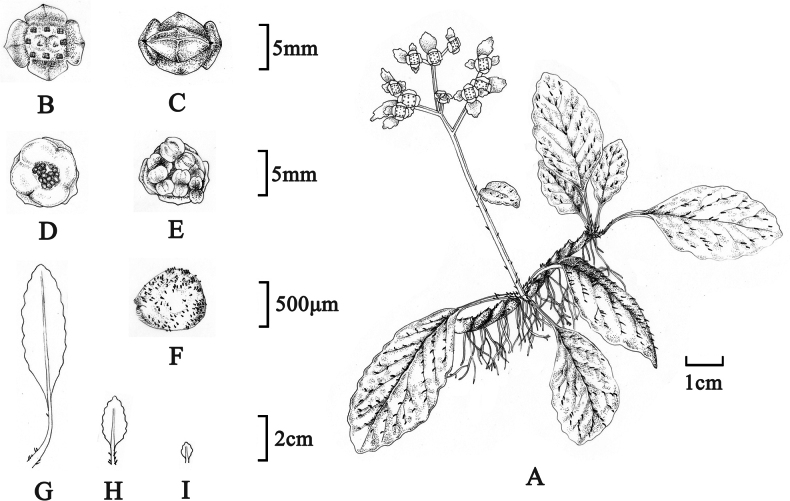 Figure 3
Figure 3 Figure 4
Figure 4| Species | Location | Voucher specimens | Herbarium | Plastid GenBank number | nrDNA GenBank number |
|---|---|---|---|---|---|
| Nikkou-shi, Japan | HSN09815 | HSN |
|
| |
| Gangwon Province, South Korea | KYI-2009032 | ( |
|
| |
| Shanxi, China | HZ2017050107362 | HSN |
|
| |
| Sichuan, China | HSN013113 | HSN |
|
| |
| Sichuan, China | HSN06442 | HSN |
|
| |
| Sangzhi, Hunan, China | SZ2016080907105 | HSN |
|
| |
| Georgia | P03_WF11 | ( | – |
| |
| Nikkou-shi, Japan | HSN09817 | HSN |
|
| |
| Nikkou-shi, Japan | HSN09823 | HSN |
|
| |
| Nikkou-shi, Japan | HSN09816 | HSN |
|
| |
| Nikkou-shi, Japan | HSN7797 | HSN |
|
| |
| Sichuan, China | JZ2018042507981 | HSN |
|
| |
| Sichuan, China | QCS2017102608035 | HSN |
|
| |
| Nikkou-shi, Japan | HSN09810 | HSN |
|
| |
| Shanxi, China | HSN7760 | HSN |
|
| |
| Guangxi, China | HSN13356 | HSN |
|
| |
| Sangzhi, Hunan, China | HSN7505 | HSN |
|
| |
| Hubei, China | HSN09188 | HSN |
|
| |
| Zhejiang, China | HSN7909 | HSN |
|
| |
| Shimane-ken, Japan | DG2019032310004 | HSN |
|
| |
| Nikkou-shi, Japan | HSN09818 | HSN |
|
| |
| Anhui, China | BD2017030507343 | HSN |
|
| |
| Jilin, China | HSN7379 | HSN |
|
| |
| Hubei, China | BD2017030507344 | HSN |
|
| |
| Jilin, China | CBS2016062406656 | HSN |
|
| |
| Nikkou-shi, Japan | HSN09820 | HSN |
|
| |
| Jinfo Mountain, Chongqing, China | HSNTG025 | HSN |
|
| |
| Tengchong, Yunnan, China | GLGH20170607375 | HSN |
|
| |
| Gansu, China | HSN07772 | HSN |
|
| |
| Wales, UK | BGN_RN_W | ( |
|
| |
| Nikkou-shi, Japan | HSN09819 | HSN |
|
| |
| Jilin, China | HSN09819 | HSN |
|
| |
| Sichuan, China | HSN7980 | HSN |
|
| |
| Jilin, China | SJH2017052107372 | HSN |
|
| |
| Irkutsk, Russia | P02_WC8 | ( | – |
| |
| Jilin, China | SJH2017052107371 | HSN |
|
| |
| Hunan, China | TPS2017042407504 | HSN |
|
| |
| Shanxi, China | HSN7761 | HSN |
|
| |
| Tibet, China | HSN7380 | HSN |
|
| |
| Shanxi, China | HSN13356 | HSN |
|
| |
| Shimane-ken, Japan | DG2019032310003 | HSN |
|
| |
| Nunavut, Canada | Brysting_01-065_CAN | CAN |
|
| |
| Yukon, Canada | Bennett_08-125_CAN | CAN |
|
| |
| Chile | P04_WG8 | HSN |
|
| |
| Hunan, China | ZJ2016031506369 | HSN |
|
| |
| Henan, China | ( |
|
| ||
| Hubei, China | S313 | ( |
|
| |
| Hunan, China | S371 | – |
|
|
| Characters |
|
|
|---|---|---|
| Rhizome | Rhizome thick, crossed and nodular | absent |
| Basal leaves | base long cuneate, margin 10–20-crenate | base rounded to subcordate; margin 20–36-crenate |
| Sepals | margin curl | margin uncurl |
| Seed | red, 0.59–0.85 × 0.48–0.63 mm | black, 0.50 × 0.40 mm |
| Characters |
|
|
|---|---|---|
| Basal leaves and cauline leaves | Isophyllous | heterophyllous |
| Basal leaves | long elliptic, margin 14–24-crenate; base long cuneate | orbicular, margin 34–39-crenate; base reniform |
| Characteristic |
|
|---|---|
| Size (base pair, bp) | 154,004 |
| LSC length (bp) | 80,646 |
| SSC length (bp) | 17,118 |
| IR length (bp) | 28,120 |
| Number of genes | 111 |
| Protein-coding genes | 78 |
| rRNA genes | 4 |
| tRNA genes | 29 |
| GC content | 37.51% |
| Group of Genes | Gene Name | Number |
|---|---|---|
| tRNA genes | 29 | |
| rRNA genes | 4 | |
| Ribosomal small subunit | 12 | |
| Ribosomal Large subunit | 8 | |
| DNA-dependent RNA polymerase | 4 | |
| Photosystem Ⅰ | 5 | |
| Large subunit of rubisco |
| 1 |
| Photosystem Ⅱ | 15 | |
| NADH dehydrogenase | 11 | |
| Cytochrome b/f complex | 6 | |
| ATP synthase | 6 | |
| Maturase |
| 1 |
| Subunit of acetyl-CoA carboxylase | 1 | |
| Envelope membrane protein |
| 1 |
| Protease | 1 | |
| Translational initiation factor |
| 1 |
| c-type cytochrome synthesis |
| 1 |
| Conserved open reading frames( | 4 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and Fungal Species Descriptions · Plant Diversity and Evolution · Plant Taxonomy and Phylogenetics
Introduction
Chrysosplenium L. (Saxifragaceae) comprises more than 70 species of perennial herbs (Kim et al. 2019; Fu et al. 2020, 2021). Chrysosplenium is distributed throughout Asia, America and Europe (Pan and Ohba 2001; Soltis 2007). The latest checklist of Chinese Chrysosplenium included 35 species (Pan and Ohba 2001). Their earlier revisions classified the genus into two subgenera (subg. Chrysosplenium and subg. Gamosplenium) based on leaf arrangement (Pan 1986a, b). This character was also considered by Hara, who divided Chrysosplenium into two sections, namely sect. Alternifolia and sect. Oppositifolia (Hara 1957). The following molecular analyses (Soltis et al. 2001; Fu et al. 2021) demonstrated that these two subgenera/sections are monophyletic and sister to each other, further confirming that leaf arrangement is a good indicator of the relationships within the genus. However, a recent systematic study based on a complete chloroplast genome and nrDNA data challenged this relationship as their results recovered an additional clade composed of two species with alternate leaves (as members of sect. Alternifolia). The newly defined clade was recognized as a basal clade sister to the rest of the species of Chrysosplenium (Yang et al. 2023).
Chrysospleniumguangxiense H.G.Ye & Gui C.Zhang was first described as a new species in 1994 by having ovate-elliptic leaves, an acuminate apex, a cuneate base, fewer dentate margins, and a depressed sepal apex, enabling it to be distinguished from its similar species, C.glossophyllum H.Hara (Ye and Zhang 1994). Subsequently, it appeared as a synonym of the latter in Flora of China without additional explanation (Pan and Ohba 2001). We assumed that the authors considered these differences were insufficient to distinguish them. It is possible that the scarcity of C.glossophyllum species in China, with its only population in Sichuan Province, may have led to this misinterpretation. Molecular data, however, could provide a means to confirm the systematic position of morphological similarities and to evaluate the phylogenetic informativeness of morphological characters (Scotland et al. 2003).
In 2019, we conducted an extensive investigation in Tianlin County, Baise City, Guangxi, China, the type locality of Chrysospleniumguangxiense. We collected a plant of Chrysosplenium, which was then confirmed as C.guangxiense. Following a thorough literature survey (Hara 1957; Pan 1992; Pan and Ohba 2001; Liu et al. 2016; Wei 2018; Kim et al. 2019; Fu et al. 2020, 2021; Wei et al. 2022), along with the molecular evidence, it was confirmed that C.guangxiense is a different species from C.glossophyllum.
Materials and methods
Morphology observations and conservation assessments
All morphological characters were studied based on the material from field and herbarium specimens using a dissecting microscope (SMZ171, Motic, China). For seed morphology, we also undertook scanning electron microscope (SEM) observations; seeds were collected from the field and dried with silica gel. The pre-treatments, including impurity removal, air-drying, and gold-coating, were performed following Fu et al. (2020). Observations and photographs were taken under a Hitachi SU8010 scanning electron micrograph. At least 15 seeds were used to determine their size and ornamentation. A conservation assessment was undertaken following the IUCN (2019).
Genomic DNA extraction, sequencing, plastid genome and nrDNA assembly and annotation
The genomic DNA was extracted using the modified CTAB method (Doyle and Doyle 1987). The short-insertion library (300 bp) was constructed and then sequenced to obtain 2×150 bp paired-end data using the Illumina NovaSeq platform at Majorbio Company (Shanghai, China). The raw data was filtered through Trimmomatic v. 0.39 (Bolger et al. 2014) to obtain clean data, and then the clean data were quality-controlled using FastQC v. 0.11.9 (Simon 2020). The complete plastid genome and nrDNA sequence were assembled using GetOrganelle v. 1.7.5 (Jin et al. 2020), and annotation was performed using CPGAVAS2 (Shi et al. 2019) and PGA (Qu et al. 2019).
Phylogenetic analysis
To confirm the phylogenetic placement of Chrysospleniumguangxiense, we undertook phylogenetic studies using the chloroplast (CP) genomes and nrDNA sequences obtained in a previous study (Yang et al. 2023). Forty-seven species of Chrysosplenium as in-group, and two species from other genera in Saxifragaceae and Iteachinensis Hook. & Arn. from Iteaceae as an out-group were sampled. The species names and GenBank accession numbers are listed in Table 1.
The chloroplast protein-coding genes (cpPCGs) were extracted from the CP genome using PhyloSuite v.1.2.3 (Zhang et al. 2020). These cpPCGs and nrDNA sequences were aligned by MAFFT v. 7.4 (Katoh and Standley 2013), and concatenated using PhyloSuite v.1.2.3 (Zhang et al. 2020) to form the cpPCGs+nrDNA matrix. The phylogenetic analyses of Chrysosplenium based on cpPCGs, nrDNA and cpPCGs+nrDNA matrices were performed using maximum likelihood (ML) and Bayesian inference (BI), respectively. The ML analyses were conducted using IQ-TREE v. 2.1.2 (Nguyen et al. 2015) with 1,000 bootstrap replicates and the default ModelFinder (Kalyaanamoorthy et al. 2017) to find GTR+F+I+G4 as the best-fit substitution model. Tree visualization was achieved in Figtree v. 1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/). For BI analysis, MrBayes v. 3.2.6 (Ronquist et al. 2012) was employed to obtain a maximum clade credibility (MCC) tree. BI analysis was performed using one million generations, two runs, 25% trees discarded as burn-in, and trees sampled every 1,000 generations (1,000 trees sampled in total) with the GTR model.
Results
Characteristics of the complete chloroplast genome
The CP genome of Chrysospleniumguangxiense comprised 154,004 bp (Fig. 1). The characteristics and statistics of the CP genome are summarized in Tables 4, 5.
Plastid genome map of Chrysospleniumguangxiense. The thick lines on the outer complete circle identify the inverted repeat regions (IRa and IRb). The innermost track of the plastome shows the GC content. Genes on the outside and inside of the map are transcribed in clockwise and counter directions, respectively.
Molecular phylogenetic studies
The cpPCGs matrix length was 71,919 bp, including 6,392 parsimony informative sites, 13,645 variable sites, and 55,865 conserved sites. The nrDNA matrix was 6,738 bp in length, with 765 parsimony informative sites, 1,200 variable sites, and 5,231 conserved sites. The cpPCGs+nrDNA matrix was 78,657 bp in length, with 7,157 parsimony informative sites, 14,845 variable sites, and 61,096 conserved sites. The phylogenetic tree of the cpPCGs matrix exhibited high confidence, while the phylogenetic tree of the nrDNA matrix had some branches with low support, and was significantly different from the former (Suppl. materials 1, 2). However, Chrysospleniumguangxiense was consistently related to C.hydrocotylifolium H.Lév. & Vaniot in both chloroplast and nuclear gene trees (Suppl. materials 1, 2). The phylogenetic tree of the cpPCGs+nrDNA matrix received a higher confidence value compared to trees generated from subsets (cpPCGs and nrDNA). Topologies obtained from BI and ML methods were congruent and showed that Chrysosplenium species clustered in a strongly supported clade (BS = 100%, PP = 1) which was further divided into three well-supported clades (defined as A-C clades; Fig. 2). Chrysospleniumguangxiense was recognized as a member of clade B and fell in its basal clade (BS = 100%, PP = 1; Fig. 2), which also included C.macrophyllum Oliv., C.zhangjiajieense X.L.Yu, Hui Zhou & D.S.Zhou, C.hydrocotylifolium, C.flagelliferum F.Schmidt, and C.zhouzhiense Hong Liu. Of these, C.guangxiense was most closely related to C.hydrocotylifolium (BS = 100%, PP = 1; Fig. 2). Although C.glossophyllum was also a member of clade B, it fell into a much more distant clade from C.guangxiense (Fig. 2).
Phylogenetic tree of Chrysosplenium generated from maximum likelihood (ML) and Bayesian inference of cpPCGs+nrDNA dataset. Numbers below the branches indicate bootstrap values (≥50%) of the ML analyses and the posterior probability (≥0.5) of Bayesian analyses.
Morphological observations
A suite of morphological characters including rhizome size, leaf shape, leaf margin dentate, sepal apex shape, and seed size of Chrysospleniumguangxiense and C.glossophyllum was consulted or observed. Chrysospleniumguangxiense had a robust rhizome, basal leaves with a long cuneate base and fewer teeth in the margin, curled sepal margins, and red, larger seeds that make it easily distinguished from C.glossophyllum (Table 2). Considering the phylogenetic results, a morphological comparison between C.guangxiense and C.hydrocotylifolium was also conducted. Chrysospleniumguangxiense had long elliptic leaves and a long cuneate leaf base, which can be easily distinguished from C.hydrocotylifolium (Table 3).
Discussion
Our phylogenetic result supported the monophyly of Chrysosplenium (Soltis et al. 2001; Fu et al. 2021; Yang et al. 2023). Besides two well-defined clades (denoted as sect. Oppositifolia and sect. Alternifolia), our result also revealed a third clade comprising two species from sect. Alternifolia, the topology of which is consistent with the previous study (Yang et al. 2023). This phylogenetic relationship indicated a non-monophyletic status of sect. Alternifolia and suggested that a deeper morphological character evolution across this phylogenetic framework is needed to evaluate the phylogenetic informativeness of characters.
In our phylogenetic tree, Chrysospleniumguangxiense was recovered as a member of sect. Alternifolia, most closely related to Chrysospleniumhydrocotylifolium (BS = 100%, PP = 1) but had a distant relationship with C.glossophyllum. It was easy to distinguish C.guangxiense from C.hydrocotylifolium by the long elliptic leaves and long cuneate leaf bases (Table 3). Our morphological comparison between C.guangxiense and C.glossophyllum also showed a suite of characters, including having a robust rhizome, basal leaves with a long cuneate base and fewer teeth in margin, and larger seeds in C.guangxiense which helped distinguish it from C.glossophyllum (Table 2). Furthermore, there was a typical viviparous phenomenon of C.guangxiense; the mature seeds were able to germinate directly in the opening capsule (Figs 3E, 4G, H). This feature has not been reported in any other Chrysosplenium species so far. Therefore, our molecular and morphological evidence supports C.guangxiense as a distinct species that resurrected from C.glossophyllum. We presented the following detailed taxonomic treatment for C.guangxiense.
Illustration of Chrysospleniumguangxiense H.G.Ye & Gui C.Zhang A habit in flowering phase B flower C indehiscent capsule D dehiscent capsule and seeds E germinated seeds in capsule F seeds G caulline leaf H, I bracteal leaf.3
Plate of Chrysospleniumguangxiense H.G.Ye & Gui C.Zhang A habit B, C inflorescence with flowers D, E basal leaves F fruit and seed G, H germinated seeds and seedlings in capsule I, JSEM of seed (Photos by Hong Liu).
Taxonomic treatment
Chrysosplenium
guangxiense
Taxon classificationPlantaeSaxifragalesSaxifragaceae
H.G.Ye & Gui C.Zhang in Acta Bot. Austro Sin. 9: 57, f. 1 (1994)
5F00CD5C-24B3-5D92-AB92-0C2B3AD0555D
Type.
Tian Lin, 11 Oct. 1989, South China Exped. 2458 (holotype: IBSC!; isotype: IBSC!).
Description.
Perennial herbs, 5.5–17 cm high. Root fibrous and robust. Long creeping rhizome developed, thick, crossed and nodular, 1–2 cm between each node, without stolons and bulbs. Flowering stem(s) always 1, erect, branched, 10–17 cm high, sparsely pilose, green, squared. Sterile branches arise from all basal leaves. Isophyllous, Basal leaves 3–6, alternate and clustered; petiole 1–6.5 cm long, white pilose; leaf blade long elliptic, 2.2–10.3 × 1.8–3.3 cm, abaxially glabrous, light green, adaxially with sparse long hispid, dark green, apex rounded, margin 14–25-crenate, base long cuneate. Cauline leaves always 1, petiole 1.3–2.2 cm long; blade 2.2–4.0 × 1.2–1.9 cm, long elliptic, glabrous in the abaxial side and with sparse hispid in the adaxial side; apex obtuse; margin obtusely dentate (9–13 teeth); base broadly cuneate; veins obvious in adaxial. Pleiochasium 9–16 cm wide, 10–15 cm high, extremely diffused, with 5–20-flowered cyme, branches sparsely hispid, surrounded by bracts; bracteal leaves green, elliptic to broadly ovate or round, glabrous. Flowers tetramerous, actinomorphic; sepals 4 (2 pairs), flat, green, 0.9–1.2 × 2.1–4.3 mm, broadly ovate, apex acute, with margin curls outward in fruiting time; disk obvious; stamens 8, ca. 0.2 mm long, shorter than sepals; anthers orange, 2-locular, longitudinally dehiscent; ovary 2-locular, semi-inferior; stigma 2; styles erect, ca. 0.2 mm long. Fruit a capsule, 5–7 mm long, green, smooth, 2-lobed (horn-shaped), equal, dehiscent along the adaxial suture; seeds numerous, red or reddish brown, obovoid, a raphe on one side, 594.19–855.33 × 475.41–625.7 μm, long papillose. Viviparous.
Additional specimen examined.
Chrysospleniumglossophyllum. China. Sichuan: Kuan County (Dujiangyan City), 19 April 1930, F. T. Wang 20553 (PE!, NAS!); same locality, 6 May 1987, Xintang Ma & Zhilong Zhao 87-0521 (WCSBG!); same locality, 15 April 2013, LiXJ 353 (KUN!); same locality, 24 May 2016, Hong Liu, HSN06644 (HSN!); same locality, 26 October 2017, Hong Liu, HSN08105 (HSN!). Chrysospleniumguangxiense. China. Guangxi: Lingyun County, Baise City, 6 March 2014, Lingyun team 451027140305005 (GXMG!); Tianlin County, Baise City, 27 November 2019, Hong Liu HSN13356 (HSN!).
Conservation status.
Chrysospleniumguangxiense is only known from two localities (IUCN criterion D2). At these two localities, the populations included ca. 200 mature individuals (IUCN criterion D1) growing in several patches. Using the IUCN methodology, C.guangxiense is classified as Vulnerable (VU) based on criteria D1 and D2: population size and the number of locations, combined with a plausible future threat that could drive this taxon to Critically Endangered or Extinct in a very short time. However, the vivipary of C.guangxiense may strengthen its adaptability to cope with future climate and environmental changes. The future threat is mainly due to grazing.
Conclusions
The phylogenetic analyses using plastomes and nuclear gene sequences of Chrysospleniumguangxiense reveal that C.guangxiense belongs to the sect. Alternifolia, is closely related to Chrysospleniumhydrocotylifolium, but distant from C.glossophyllum based on leaf morphology and other traits. Our findings support the resurrection of C.guangxiense as a distinct species and provide a detailed taxonomic treatment for its identification. The phylogenetic analyses confirm the monophyly of Chrysosplenium and reveal a non-monophyletic status of sect. Alternifolia. Further systematic studies of Chrysosplenium should focus on finding additional morphological characters with phylogenetic informativeness to disentangle the non-monophyletic sect. Alternifolia, and propose a new infrageneric classification and provide a stable framework for answering broader questions in evolutionary biology.
Supplementary Material
XML Treatment for Chrysosplenium guangxiense
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bolger AM Lohse M Usadel B (2014) Trimmomatic: A flexible trimmer for Illumina sequence data.Bioinformatics (Oxford, England)30(15): 2114–2120. 10.1093/bioinformatics/btu 17024695404 PMC 4103590 · doi ↗ · pubmed ↗
- 2Chen Z Yu X Yang Y Wei P Zhang W Li X Liu C Zhao S Li X Liu X (2022) Comparative analysis of chloroplast genomes within Saxifraga (Saxifragaceae) takes insights into their genomic evolution and adaption to the high-elevation environment.Genes 13(9): 1673. 10.3390/genes 1309167336140840 PMC 9498722 · doi ↗ · pubmed ↗
- 3Doyle JJ Doyle JL (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissues.Phytochemical Bulletin 19: 11–15.
- 4Folk RA Stubbs RL Mort ME Cellinese N Allen JM Soltis PS Soltis DE Guralnick RP (2019) Rates of niche and phenotype evolution lag behind diversification in a temperate radiation.Proceedings of the National Academy of Sciences of the United States of America 116(22): 10874–10882. 10.1073/pnas.181799911631085636 PMC 6561174 · doi ↗ · pubmed ↗
- 5Fu LF Liao R Lan DQ Wen F Liu H (2020) A new species of Chrysosplenium (Saxifragaceae) from Shaanxi, north-western China.Phyto Keys 159: 127–135. 10.3897/phytokeys.159.5610932973392 PMC 7486313 · doi ↗ · pubmed ↗
- 6Fu LF Yang TG Lan DQ Wen F Liu H (2021) Chrysospleniumsangzhiense (Saxifragaceae), a new species from Hunan, China.Phyto Keys 176: 21–32. 10.3897/phytokeys.176.6280233958936 PMC 8065015 · doi ↗ · pubmed ↗
- 7Hara H (1957) Synopsis of genus Chrysosplenium L. Journal of the Faculty of Science, University of Tokyo, Section III.Botany 7: 1–90.
- 8IUCN (2019) Guidelines for using the IUCN red list categories and criteria. Version 14. Prepared by the Standards and Petitions Committee.