The polyphyletic Caucasus-centred Campanula subg. Scapiflorae (Campanulaceae) revisited with a newly circumscribed C. sect. Tridentatae for its core clade
Nana Silakadze, Marine Mosulishvili, Thomas Borsch, Norbert Kilian

TL;DR
This study reclassifies a group of alpine plants in the Caucasus, confirming their genetic diversity and introducing a new section for their core group.
Contribution
A revised classification of the Campanula subg. Scapiflorae group and a newly defined C. sect. Tridentatae for its core clade.
Findings
Molecular analysis confirmed the polyphyly of Campanula subg. Scapiflorae in all its previous circumscriptions.
The core clade of the group is composed solely of Caucasus endemics and is reclassified as C. sect. Tridentatae.
Phylogenetic relationships of other elements in the Scapiflorae group are outlined.
Abstract
Campanula L. is among the genera with the highest number of endemics in the Caucasus ecoregion. A group of attractive alpine and subalpine perennial rosette plants with short single-flowered stems centred in the Caucasus has been treated as Campanulasubg.Scapiflorae or at other ranks, with considerably varying circumscription and classification. Molecular phylogenetic analysis of three plastid DNA regions (trnK/matK, petD, rpl16) of a strongly extended sampling, comprising 23 of the 27 commonly accepted taxa (85%) with 330 accessions built on and guided by the results of our previous study of the group, confirmed the polyphyly of C.subg.Scapiflorae in any of its circumscriptions. The core clade of the group comprises exclusively endemics and near-endemics of the Caucasus and is treated here as C.sect.Tridentatae in a revised circumscription. The phylogenetic relationships of the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
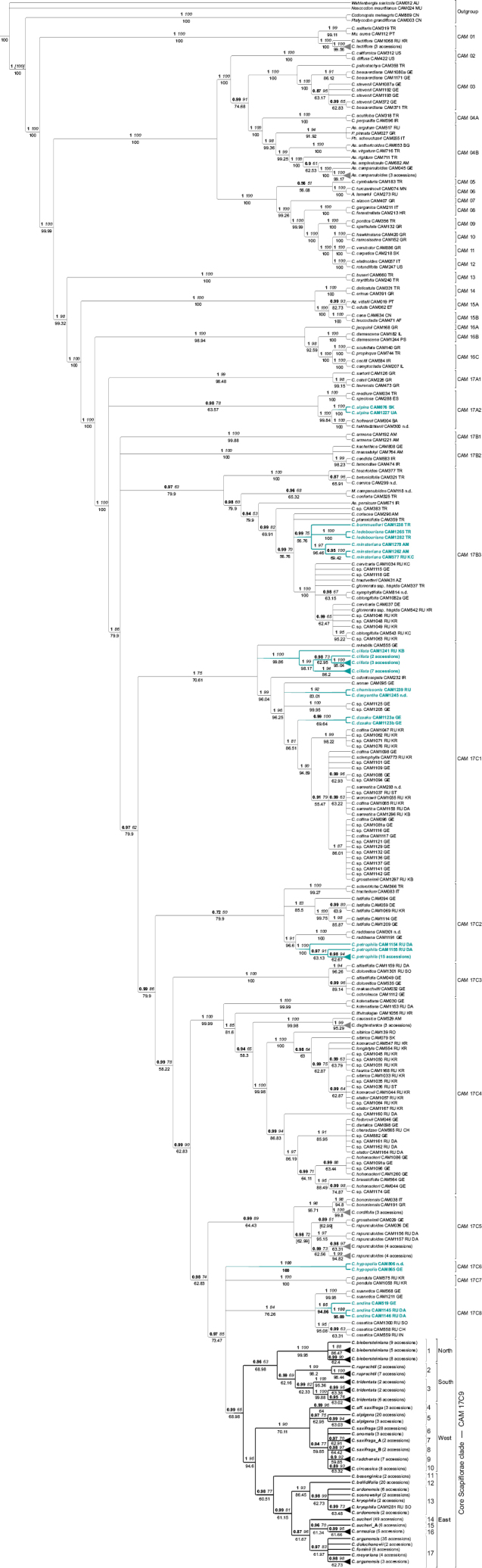 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Plant Parasitism and Resistance · Plant and Fungal Species Descriptions
Introduction
With approximately 600 species (Lammers 2007; Mansion et al. 2012; Jones et al. 2017), Campanula in the wide sense is among the one hundred largest plant genera (Frodin 2004), representing also the largest genus in the family Campanulaceae (order Asterales). Although Campanula was formally established by Linnaeus, it was first described more than 200 years earlier by Fuchs (1542), based on the species later named C.trachelium L. Campanula includes mostly perennial herbs with alternate leaves, bell-shaped, mostly purplish blue, pentamerous, bee-pollinated flowers and capsule fruits (Candolle 1830; Boissier 1875; Fedorov 1957). The members of the genus are found in forests, meadows, steppes and semi-deserts in temperate and subtropical zones of the northern hemisphere and is most abundant and diverse in the subalpine and alpine zones of mountain ranges (Fedorov 1957; Kovačić 2004, Jones et al. 2017). In the Caucasian flora, Campanula is one of the 17 genera with the highest number of endemic species (Dolukhanov 1966; Zazanashvili and Mallon 2009).
One group of species in Campanula, hitherto recognized as C.subg.Scapiflorae (Boiss.) Oganesian, is mainly endemic to the Caucasus. The group is characterised by single flowers, basal leaf rosettes and preference of alpine and subalpine habitats with rocky substrate (Silakadze et al. 2019). Sharing reflexed appendages between the calyx lobes and 3-locular capsules dehiscing down to the base led Fedorov (1957) to conclude that they belong to the core of Campanula. This core group corresponds to the large clade CAM 17 found by Mansion et al. (2012) in their overall analysis of Campanula and allies, which also includes C.latifolia L., the type of the name Campanula. In contrast, the genus in any of its current wider circumscriptions is excessively paraphyletic to a number of segregate genera mostly differing in floral traits (Mansion et al. 2012; Xu and Hong 2020).
The Scapiflorae group is an example of a complex species group in Campanula in the sense that different authors arrived at very different conclusions on the numbers and boundaries of the taxa to be recognized in this group. Whereas Ruprecht (1867), Boissier (1875) and Fomin (1905) had recognized 13, eight and 12 species, respectively, Fedorov (1957) and Kharadze (1949, 1976) recognized many more species (26 and 24, respectively). Oganesian (2000) accepted a similar number of taxa (25), of which 20 were at the species level and five additional subspecies. Seven of the species accepted by Fedorov (1957) and Kharadze (1976) were treated by her as synonyms, whereas, on the other hand, she added eight more species not included to the Scapiflorae before. In the most recent treatment Victorov (2001, 2002) recognized 11 species with seven subspecies. Whereas Mansion et al. (2012) just placed several of these species in one of the major Campanula clades (CAM 17), the addition of further molecular characters by Jones et al. (2017) and Silakadze et al. (2019) revealed many of them as part of a well-supported subclade of CAM 17, whereas others appeared distantly. However, several species hitherto classified in Scapiflorae remained unsampled.
The majority of species that were considered as part of the Scapiflorae group are endemics of the Caucasus. The distribution area of two species Campanulaledebouriana Trautv. and C.minsteriana Grossh. extends from the Caucasus into Anatolia and adjacent mountains south-eastwards; two species, C.bornmuelleri Nábělek and C.pulvinaris Hausskn. & Bornm., occur in Anatolia only, and two further are European, of which one (C.alpina Jacq.,) is restricted to the eastern Alps and the Carpathians, the other (C.orbelica Pančić) to the mountains of the central and eastern Balkans (Ronikier and Zalewska-Gałosz 2014); two species, C.capusii (Franch.) Fed. and C.lehmanniana Bunge, occur in middle Asia and two species, C.chamissonis Fed. and C.dasyantha M. Bieb., are distributed in North-East Asia. Moreover, recent phylogenetic studies (Silakadze et al. 2019) revealed that C.ciliata Steven, the type of the name C.subg.Scapiflorae, is only distantly related to the core clade of this group, and more closely related to C.latifolia, the type species of the name Campanula.
The present paper has three aims: Based on a considerably increased sampling of Campanula species in the Caucasus region and species previously assigned to C.subg.Scapiflorae (1) to further evaluate the composition of the Scapiflorae clade (in the sense of Silakadze et al. 2019), (2) to update the classification of the Scapiflorae group at the supraspecific level; and (3) to further examine phylogenetic relationships within this clade.
Materials and methods
Taxon sampling
We densely sampled the species diversity of Campanulasubg.Scapiflorae across its entire geographical range. Samples were collected during fieldwork in Armenia, Georgia, and Russia, and additional material from various countries was examined from the herbaria of B, DAG, ERE and TBI (Appendix 1). Compared to our previous study (Silakadze et al. 2019), we added nine more taxa with multiple accessions and covering a wider geographical range of the Caucasus region. These include eight Caucasian endemics, i.e., C.anomala Fomin, C.ardonensis Rupr., C.besenginica Fomin, C.doluchanovii Kharadze, C.fominii Grossh., C.kadargavanica Amirkh. & Komzha, C.kryophila Rupr. C.sosnowskyi Kharadze and the Caucasian and East Anatolian C.ledebouriana (see Suppl. material 3). One accession previously identified as C.cf.aucheri A. DC. (CAM217) was excluded from our extended dataset, as we were not sure about the accuracy of the sequences. For C.doluchanovii and C.meyeriana Rupr., we were able to include sequences from type material. We also included additional accessions of two species C.ciliata and C.petrophila Rupr., to test with material from different localities if these species are indeed not part of the Scapiflorae clade.
Our sampling of the Scapiflorae group contained 23 of the 27 commonly accepted taxa (85%) with 330 (271 newly included) accessions. In addition, we increased the sampling of other Caucasian Campanula species, located in the CAM 17C clade (Mansion et al. 2012; Jones et al. 2017; Silakadze et al. 2019), by 54 new accessions, some species from multiple localities. Overall, our sampling of Campanula was increased by 325 additional sequences representing three genomic regions of the plastid DNA (trnK/matK, petD and rpl16), more than doubling the sampling of our previous study (Silakadze et al. 2019).
DNA extraction, amplification, sequencing, and alignment
DNA extraction, amplification, sequencing and alignment followed Silakadze et al. (2019). For newly generated sequences from the leaf tissue of older herbarium specimens, we used a CTAB protocol with extraction of three fractions for each plant sample (Borsch et al. 2003). As DNA was often much degraded and also contained secondary metabolites, we combined fractions I, II and III of each sample and cleaned them using DNeasy PowerClean Pro Cleanup Kit. DNA size (bp) and concentration (ng/μl) were checked using Fragment Analyser (www.aati-us.com), and 10 ng/μl was considered as ideal for the polymerase chain reaction (PCR). DNA samples were diluted with purified water depending on the concentration; if the concentration was less than 10 ng/μl, the amount of DNA was duplicated in the PCR mix. DNA stocks were kept at −20 °C.
For amplification and sequencing, we mostly used the same primers as in our previous study (Silakadze et al. 2019). For petD we used shorter primers designed by Schäferhoff (unpublished), and, in addition, we designed new, shorter internal primers for rpl16 and trnK/matK (see Suppl. material 4). Primer design was carried out using the Campanula alignment published in Silakadze et al. (2019), using the program Seqstate 1.4.1 (Müller 2005).
All pherograms were checked using PhyDE version 0.9971 (Müller et al. 2010) and manually assembled using the motif-based alignment approach for non-coding plastid DNA (Kelchner 2002; Borsch et al. 2003; Löhne and Borsch 2005). Indels were coded as binary characters using the Simple Indel Coding approach (Simmons and Ochoterena 2000) as implemented in SeqState version 1.4.1 (Müller 2005). Consensus DNA sequences were submitted to ENA (European Nucleotide Archive) (www.ebi.ac.uk/ena/), using the software tool EMBL2checklists (Gruenstaeudl and Hartmaring 2019).
Phylogenetic analysis
We used the matrix including the same plastid DNA regions (trnK/matK, petD and rpl16) as in Silakadze et al. (2019). Phylogenetic analyses were performed using maximum parsimony (MP), maximum likelihood (ML) and Bayesian inference (BI) approaches.
Maximum parsimony (MP) analysis was done in PAUP version 4.0b10 (Swofford 2002), using the parsimony ratchet settings (Nixon 1999) conducted in PRAP version 2.0b3 (Müller 2004). As ratchet parameters, we selected 1,000 iterations, unweighting 25% of the positions randomly (weight = 2), and 100 additional random cycles. Jackknife (JK) support was derived through a single heuristic search in PRAP with 10,000 replicates using tree bisection-reconnection (TBR) branch swapping and in each replicate, 36.79% of the characters were deleted.
Maximum likelihood (ML) analyses were executed with RAxML version 8.2.12 (Stamatakis 2014) on the CIPRES Science Gateway V 3.3 (Miller et al. 2011), using the CAT approximation (Stamatakis 2006) of the GTR model of the DNA partitions and BINCAT for the binary indel partitions (Stamatakis 2014). Rapid bootstrap analyses (BS) were conducted with 1,000 iterations integrated with a thorough ML search.
For Bayesian inference, the nucleotide dataset was divided into six partitions and the likelihood scores of models of sequence evolution were calculated for each using jModelTest version 2.1.7 (Darriba et al. 2012), choosing the best-fitting model under Akaike’s Information Criterion (AIC). Three partitions represented trnK/matK (trnK 5’ intron = GTR+G, matK gene = TVM+G, and trnK 3’ intron = GTR+G), two partitions petD (petB-petD intergenic spacer = TVM+I+G and petD exon/intron = TVM+G), and one partition the rpl16 intron (TVM+I+G). The indel matrices were added using the restriction site model (Ronquist and Huelsenbeck 2003).
Bayesian inference analyses was carried out in MrBayes v.3.2.7.a (Ronquist et al. 2011) on CIPRES (Miller et al. 2011), with four runs and four chains each performed for 20 million generations, sampling every 5000^th^ generation. We checked convergence of the runs into stationarity by examining the average standard deviation of split frequencies and post-burn-in effective sampling size (ESS). The first 10% of trees were discarded as burn-in; the remaining trees were used to construct a 50% majority-rule consensus tree.
Additionally, Maximum parsimony (MP) and Maximum likelihood (ML) were applied to a matrix with two further species, Campanulakadargavanica, and C.pulvinaris, for which only petD and rpl16 sequences were available.
Results
Phylogenetic analysis
The final alignment of three combined plastid genomic regions (trnK/matK, petD, and rpl16) containing 536 concatenated sequences (325 newly generated) had a total length of 5,361 positions, of which trnK/matK had 2,854, petD 1,099 and rpl16 1408. To document intraspecific variation, multiple samples for the same taxa were maintained in the alignment. In the multiple sequence alignment of trnK/matK, we excluded six hotspots of uncertain homology or poly-A/T microsatellites [positions 633–738; 746–779; 785–788; 2539–2627; 2754–2763; 2769–2773]. In petD one hotspot was excluded with poly-A [position 3681–3684] and in rpl16 ten hotspots were excluded, including some poly A/T microsatellites or other sequence elements [positions 3962–3969; 4066–4083; 4123–4132; 4239–4249; 4380–4390; 4417–4419; 4463–4470; 4811–4820; 4835–4846; 5219–5252]. The final concatenated plastid matrix contained 4,984 bases (2,606 bp of trnK/matK, 1,095 bp of petD, and 1,283 bp of rpl16, respectively). Simple Indel Coding provided further 270 binary characters (104 of trnK/matK, 61 of petD, and 105 of rpl16).
Bayesian inference, maximum likelihood, and maximum parsimony analyses of the concatenated plastid dataset produced largely identical topologies and revealed significant statistical support values for various nodes at PP > 0.95 and BS and JK values > 70%, and strong support values at PP > 0.99 and BS and JK values > 90%.
Twelve species hitherto considered as members of the Scapiflorae group were found distantly related to the core Scapiflorae clade (Fig. 1; see also Suppl. materials 1, 2, 3).
Bayesian 50% majority-rule consensus tree of the combined dataset of the Caucasian Campanula species based on three plastid markers (trnK/matK, petD and rpl16). Values above nodes indicate posterior probabilities (bold) and maximum likelihood bootstrap support (italic), values below nodes indicate maximum parsimony jackknife support, values in square brackets indicate conflicting topologies. Sample designations include the taxon name, DNA lab code, and ISO (international organization for standardization) country code, in case of the Russian part of the Caucasus also the TDWG (Biodiversity Information Standards) code of the territory (n.d. – sample not documented); for DNA lab codes and ISO codes of collapsed terminals with multiple accessions, see Suppl. materials 1, 2. Abbreviations in species names: A. – Adenophora, As. – Asyneuma, Az. – Azorina, C. – Campanula, G. – Githopsis, M. – Michauxia, Mu. – Musschia, P. – Petromarula, Ph. – Phyteuma. Sample designations in bold (in black) indicate species from the core Scapiflorae clade, sample designations in blue indicate Scapiflorae group members phylogenetically distant from the core clade. Subdivision of the Scapiflorae clade by numbered terminal clades (1-17) and geographical distribution of the lineages.
Campanulaalpina, which is sister to C.hofmanii (Pant.) Greuter & Burdet and C.takhtadzhianii Fed. with strong support (PP = 1, BS = 100, JK = 99.84), was resolved in clade CAM 17A2.
Campanulabornmuelleri, C.ledebouriana and C.minsteriana were found related to each other in a polytomy with good support (PP = 0.99, BS = 75, JK = 56.76), all nested in clade CAM 17B3.
Campanulaciliata, C.dasyantha, C.chamissonis and C.dzaaku Albov were resolved in the clade CAM 17C1 with good support (PP = 1, BS = 75, JK = 70.61). The various C.ciliata samples appear all in one clade which forms a trichotomy with C.mirabilis Albov and a clade including the remainder of CAM 17C1. In the last clade, C.dasyantha and C.chamissonis formed a sister-group with strong support (PP = 1, BS = 92, JK = 83.01), and the two C.dzaaku samples emerged in a separate subclade of their own with strong support (PP = 0.99, BS = 100, JK = 69.64).
Campanulapetrophila was found sister to C.raddeana Trautv. with strong support (PP = 1, BS = 91, JK = 96.6), nested in clade CAM 17C2. Notably, C.latifolia, the type of the genus name, fell in the same clade as C.petrophila.
Campanulahypopolia Trautv. (Clade CAM 17C6) and C.andina Rupr., forming a trichotomy (Clade CAM 17C8) with C.suanetica Rupr. and C.ossetica M. Bieb., were found nested in the polytomy with the C.pendula (Clade CAM 17C7) and the core Scapiflorae clade (CAM 17C9).
The additional tree (see Suppl. material 2; ML and MP), including further species of which only petD and rpl16 sequences were available, revealed that also Campanulapulvinaris is not part of the Scapiflorae clade and instead nested in clade CAM 17B3 with C.bornmuelleri, C.ledebouriana and C.minsteriana, although without support. Notably, C.latifolia, the type of the genus name, fell in the same clade as C.petrophila, being sister in a polytomy to two clades, one containing C.petrophila and C.raddeana and another C.sclerotricha Boiss. and C.trachelium; all are nested in clade CAM 17C2, which is, however, weakly supported (PP = 0.72, BS = 50, JK = 79.9).
The majority of the taxa of the Scapiflorae group were resolved in clade CAM 17C9, here considered the core Scapiflorae clade. Within the well-supported Scapiflorae core clade (PP = 0.99, BS = 65, JK = 68.9; Fig. 1, see also Suppl. material 1) 17 terminal clades were resolved, which form two larger lineages and sister clades: the lineage A or north/south clade (PP = 0.86, BS = 63, JK = 68.98; Fig. 1, see also Suppl. material 1) and the strongly supported lineage B or west/east clade (PP = 1, BS = 95, JK = 94.6; Fig. 1, see also Suppl. material 1) .
The clade A1 corresponds to the C.biebersteiniana Roem. & Schult. accessions from the Greater Caucasus, extending to the northern part of the Caucasus region, and hence clade A1 is also referred to as north (N) clade, exhibiting strong supported (PP = 1, BS = 100, JK = 99.95).
The clade A2 contains the terminal clades 2 and 3 with the accessions of C.ruprechtii Boiss. and C.tridentata Schreb., all from the Lesser Caucasus, extending to the southern part of the Caucasus, and so this clade was also referred to as south (S) clade, likewise moderately supported (PP = 99.69, BS = 69, JK = 62.16).
The clade B1 received strong support (PP = 1, BS = 90, JK = 70.11) and comprises the terminal clades 4 through 10, including the accessions of C.alpigena K. Koch, C.anomala, C.circassica Fomin, C.radchensis Kharadze, C.saxifraga M. Bieb. and C.aff.saxifraga, all from the western part of the Caucasus and hence referred to as the west (W) clade.
Finally, the clade B2, which received moderate support (PP = 0.98, BS = 77, JK = 60.51), consists of the terminal clades 11 through 17 and includes the accessions of C.ardonensis, C.argunensis Rupr., armazica Kharadze, C.aucheri, C.bellidifolia Adams, C.besenginica, C.doluchanovii, C.fominii, C.kryophila, C.meyeriana and C.sosnowskyi, all originating from the eastern part of the Caucasus, and hence referred to as the east (E) clade.
The terminal clade 1 contains 22 accessions of C.biebersteiniana and shows strong support (PP = 1, BS = 100, JK = 99.95). Clade 2 covered nine accessions of C.ruprechtii, strongly supported (PP = 1, BS = 99, JK = 98.2). Clade 3 includes ten accessions of C.tridentata with good support (PP = 0.99, BS = 82, JK = 62.33; Fig. 1, see also Suppl. material 1). Clade 4 with strong support (PP = 0.99, BS = 96, JK = 64) consists of three accessions of uncertain identifications, two labelled C.besenginica and one C.saxifraga, which we all treat as C.aff.saxifraga. Clade 5 contains 23 accessions of C.alpigena with moderate support (PP = 0.97, BS = 75, JK = 62.95), whereas clade 6 includes three accessions of C.anomala and 48 of C.saxifraga with weak support (PP = 0.94, BS = 77, JK = 59.85). Clades 7, 8, 9 and 10 are all nested within clade 6, encompassing C.saxifraga A with two accessions with moderate support (PP = 0.97, BS = 70, JK = 62.91), C.saxifraga B also with two accessions with good support (PP = 0.98, BS = 97, JK = 64.42), C.radchensis, comprising seven accessions with moderate support (PP = 0.9, BS = 92, JK = 59.85), and C.circassica with eight accessions, also with moderate support (PP = 0.89, BS = 93, JK = 63.32; Fig. 1, see also Suppl. material 1). Clade 11 consists of two accessions of C.besenginica, forming a polytomy relative to all other terminal clades in this area of the tree. Clade 12 includes 20 accessions of C.bellidifolia and has good support (PP = 1, BS = 93, JK = 86.45), whereas clade 13, nested within clade 12, covered eight accessions of C.ardonensis, three of C.kryophila and two of C.sosnowskyi with good support (PP = 0.98, BS = 99, JK = 62.73). The next terminal clade 14 included 49 accessions of C.aucheri with moderate support (PP = 87, BS = 96, JK = 61.67), with clades 15 and 16 nested within clade 14, consisting of six accessions of C.aucheri A and five of C.armazica, with moderate or good support, respectively (PP = 0.96, BS = 70, JK = 61.34; PP = 0.99, BS = 95, JK = 61.66). Finally, clade 17, also nested within clade 14, included 38 accessions of C.argunensis, two of C.doluchanovii, six of C.fominii and four of C.meyeriana, overall with moderate support (PP = 0.97, BS = 83, JK = 61.97; Fig. 1, see also Suppl. material 1).
Discussion
Revised classification of the core Scapiflorae clade
The re-circumscription and infrageneric re-classification of the iconic bell-flower genus, Campanula, is still in progress (Mansion et al. 2012; Jones et al. 2017; Xu and Hong 2020). It is evident from these and further analyses that applying a phylogeny-based genus concept to Campanula will inevitably shatter the genus as it is known traditionally but a detailed analysis of a representative spectrum of morphological characters in a phylogenetic context will be needed to evaluate if conspicuous floral differences are not just caused by adaptive shifts as part of pollination syndromes. What applies to the genus as a whole is also true for the Scapiflorae group on a small scale. Silakadze et al. (2019) and the present study revealed that C.subg.Scapiflorae is an artificial assemblage but resolved a well-supported core lineage, the core Scapiflorae clade (Fig. 1, see also Suppl. material 1). Compared to Silakadze & al. (2019), the present study provides a much more in-depth analysis of this group, based on 330 accessions, representing 85% of the taxa of this group. The present study also fully corroborates the results of the previous study by Silakadze et al. (2019) that the core Scapiflorae clade and all other elements of the polyphyletic C.subg.Scapiflorae belong to the core of Campanula (Fig. 1, see also Suppl. material 1) defined by inclusion of C.latifolia as the type of the generic name in the clade CAM 17 (sensu Mansion et al. 2012 and later authors). We can thus postulate with certainty that the group will remain in Campanula also after a future reorganisation of the genus.
Based on research conducted by Silakadze et al. (2019), the Scapiflorae clade can be clearly defined and differentiated based on a combination of morphological, distributional and ecological characteristics. These include the presence of calyx appendages, leaf rosettes with a single-flowered stem, and its members typically grow in rocky habitats within the (sub)alpine zone and occur in the Caucasus region and adjacent areas to the south. One of the main results of our studies is that Campanulaciliata, the designated lectotype (Fedorov 1957: 256) of the name Campanulaser.Scapiflorae Boiss. and all combinations based on it, falls far outside the core Scapiflorae clade. Consequently, the name Scapiflorae is not available for a formal taxonomic recognition of the core clade. Among the authors treating the Scapiflorae group, only Fedorov (1957), Kharadze (1976) and Victorov (2002) formally subdivided C.subg.Scapiflorae. Fedorov (1957) and Kharadze (1976) recognized eleven and ten series, respectively, Victorov (2002) recognized seven sections and his treatment is the one most congruent with our result. Our core clade is represented in his treatment by only two species: C.tridentata and C.bellidifolia, the latter with five subspecies. They form C.sect.Tridentatae (Kharadze) Victorov, which also comprises, contrary to our results, the north-east Asian C.dasyantha with C.chamissonis as its subspecies, and the Caucasian C.petrophila and C.andina. At section rank the infrageneric epithet Tridentatae has priority over any other name; at series rank it is one of nine series names of equal priority for the core clade (Fedorov 1957; Kharadze 1976). Considering the phylogenetic position of the core Scapiflorae clade within clade CAM17, we have chosen the rank of section for the core clade, applying the name C.sect.Tridentatae in our revised circumscription restricted to the Caucasian members only. The choice of the rank, of a section, may appear a bit of a long shot given the unsettled circumscription and classification of a monophyletic genus Campanula. However, subgenera may likely be applied at a more inclusive level to name major clades such as CAM 1 to CAM 17, also depending if these can be recognized by morphology. For the time being it should best serve the needs of an unambiguous classification to handle the Scapiflorae clade at the level of a section.
Campanulasect.Tridentatae (Kharadze) Victorov ≡ Campanulaser.Tridentatae Kharadze in Zametki Sist. Geogr. Rast. 15: 25. 1949. – Type: Campanulatridentata Schreb.
= Campanulaser.Anomalae Fed. in Komarov (ed.), Fl. SSSR 24: 197. 1957. – Type: Campanulaanomala Fomin
= Campanulaser.Ardonenses Fed. in Komarov (ed.), Fl. SSSR 24: 198. 1957. – Type: Campanulaardonensis Rupr.
= Campanulaser.Argunenses Fed. in Komarov (ed.), Fl. SSSR 24: 192. 1957. – Type: Campanulaargunensis Rupr.
= Campanulaser.Aucherianae Kharadze in Zametki Sist. Geogr. Rast. 15: 25. 1949 [“Aucheri”]. – Type: Campanulaaucheri A. DC.
= Campanulaser.Bellidifoliae Fed. in Komarov (ed.), Fl. SSSR 24: 194. 1957. – Type: Campanulabellidifolia Adams
= Campanulaser.Besenginicae Fed. in Komarov (ed.), Fl. SSSR 24: 199. 1957. – Type: Campanulabesenginica Fomin
= Campanulaser.Kryophilae Kharadze in Zametki Sist. Geogr. Rast. 15: 25. 1949. – Type: Campanulakryophila Rupr.
= Campanulaser.Saxifragiformes Fed. in Komarov (ed.), Fl. SSSR 24: 190. 1957. – Type: Campanulasaxifraga M. Bieb.
In our circumscription, the section includes all taxa belonging to the core Scapiflorae clade. These are: Campanulaalpigena, C.argunensis (incl. C.doluchanovii, C.fominii, C.meyeriana), C.armazica, C.aucheri, C.bellidifolia (incl. C.ardonensisC.sosnowskyi, C.kadargavanica, C.kryophila), C.besenginica, C.biebersteiniana, C.radchensis, C.ruprechtii, C.saxifraga (incl. C.anomala, C.circassica) and C.tridentata. They are Caucasian endemics, or near endemics extending into southerly adjacent mountain ranges, growing on rocky to soil-rich substrates predominantly in the subalpine to alpine zone and are morphologically characterised by the presence of calyx appendages, leaf rosettes with short and strictly single-flowered stems. However, the combination of these characters does not represent an exclusive synapomorphy for the core Scapiflorae clade but has evolved independently also in the single case of C.ciliata (Silakadze et al. 2019), nested in the distant clade CAM 17C1 (see below).
A revised classification of the members of Campanulasect.Tridentatae will be substantiated and elaborated in two further contributions (Silakadze et al. in prep.; Silakadze and Kilian in prep.).
Phylogenetic position of the Scapiflorae members excluded from Campanulasect.Tridentatae
According to our analysis, 12 species do not belong to the Tridentatae clade (Fig. 1, see also Suppl. material 1). These are Campanulaalpina, C.andina, C.bornmuelleri, C.chamissonis, C.ciliata, C.dasyantha, C.dzaaku, C.hypopolia, C.minsteriana, C.ledebouriana, C.petrophila and C.pulvinaris. Their phylogenetic positions are briefly discussed in the following.
The Tridentatae clade CAM 17C9 is part of a polytomy of exclusively Caucasian endemics, together with the three clades CAM 17C6, C7 and C8 (Fig. 1, see also Suppl. material 1). The first of these clades is represented by Campanulahypopolia, which was placed by Victorov in C.sect.Hypopolion (Fed.) Ogan. of his C.subg.Scapiflorae, the second is represented by C.pendula M. Bieb. The third includes C.andina, which was described from the “Andi” range of Dagestan and placed by Victorov (2002) in his C.sect.Tridentatae, together with C.suanetica and C.ossetica in an unresolved clade. Notably, none of them except the members of the Tridentatae clade have unbranched single flowered stems.
Campanulaciliata is resolved as member of clade CAM 17C1, far distant from the Tridentatae clade. All seven authors who have studied the Scapiflorae group agreed that Campanulaciliata belongs to that group (see Suppl. material 3) as defined by a set of mostly convergent morphological characters. The name Scapiflorae, with its type C.ciliata, thus applies to the clade CAM 17C1 (Fig. 1, see also Suppl. material 1), also containing the Scapiflorae members C.chamissonis, C.dasyantha and C.dzaaku, and the other Caucasian species C.annae Kolak., C.collina Sims, C.grossheimii Kharadze, C.mirabilis, C.odontosepala Boiss., C.sarmatica Ker Gawl., C.sclerophylla (Kolak.) Ogan. and C.woronowii Kharadze. At subgenus rank, C.subg.Annae (Kolak.) Ogan. is also available but does not have priority over Scapiflorae, whereas at sectional rank, C.sect.Annae would have priority for clade CAM 17C1. Within this clade, multiple accessions of the Caucasian C.ciliata form a trichotomy with C.mirabilis and the remainder of the clade, which also includes, besides the Caucasian members, C.odontosepala from Iran and the north-east Asian C.dasyantha and C.chamissonis. These Scapiflorae species and the closely related and sympatric C.aldanensis Fed. & Karav. were treated by Fedorov (1957) and Victorov (2002) as C.ser.Dasyanthae Fed. However, Victorov recognised only C.dasyantha, in which he included C.chamissonis as a subspecies (C.dasyanthasubsp.chamissonis) and sank C.aldanensis in the synonymy of the typical subspecies, a treatment we fully agree with. The placement of C.dzaaku, a species without calyx appendages and unusual coriaceous leaves with cartilagineously denticulate margin, was disputed. Kharadze (1949) and Fedorov (1957) placed it close to C.ciliata, Victorov (2002) placed it distant to the latter but as a member of Scapiflorae, and Oganesian (2000) did not at all include it in the Scapiflorae. The lack of calyx appendages was identified by Silakadze & al. (2019: fig. 4) as a synapomorphy of a subclade including C.dzaaku together with C.collina, C.sclerophylla, C.woronowii and, with a reversal in this state, C.sarmatica. As it turns out, clade CAM 17C1 is morphologically heterogenous and its internal relationships and character evolution require further study.
The Central to S European Campanulaalpina, still included by Oganesian (2000) and Victorov (2002) in the Scapiflorae group (see Suppl. material 3), is resolved very distantly to the Tridentatae clade, in the CAM 17A2 clade, sister to C.hofmanii and C.takhtadzhianii, and these in turn sister to C.medium L. and C.speciosa Pourr. (Fig. 1, see also Suppl. material 1). The morphological similarity of C.alpina to the Tridentatae members is only by convergent evolution to similar habitats and its inflorescence can, moreover, also be few-flowered (Silakadze et al. 2019). Another species closely related to C.alpina but not included in our study is the Balkan endemic C.orbelica (Ronikier and Zalewska-Gałosz 2014).
The former Scapiflorae species Campanulabornmuelleri, C.ledebouriana, C.minsteriana and C.pulvinaris are nested in CAM 17B3 is (Fig. 1, see also Suppl. material 1; for the C.pulvinaris see Suppl. material 2). They are distributed mainly in Anatolia (Turkey) and the Armenian Highlands. C.karakuschensis was included in the Scapiflorae group only by Victorov (2002) (see Suppl. material 3) and as a synonym of C.minsteriana, in line with Oganesian (2000), who classified that species under C.subg.Theodorovia (Kolak.) Ogan. C.minsteriana according to our phylogeny, is closely related to C.ledebouriana, corroborating the view of Fedorov (1957) and Kharadze (1976), who placed C.minsteriana with C.ledebouriana into C.ser.Ledebourianae Fed. of the Scapiflorae group. Also, Oganesian (2000) recognized C.ledebouriana as a member of the Scapiflorae, and additionally included C.bornmuelleri and C.pulvinaris. Our phylogenetic results confirm these to belong to one lineage together with C.coriacea P. H. Davis, C.ptarmicifolia Lam. and the members of the clade of C.glomerata L. (Fig. 1, see also Suppl. materials 1, 2). For unknown reasons, Fedorov (1957) and Kharadze (1976) treated C.karakuschensis not as conspecific with C.minsteriana but as a quite different species and outside the Scapiflorae group in C.ser.Saxicolae (Boiss.) Kharadze, together with C.lehmanniana and C.capusii. These two species are distributed in Kirgizistan and Tadzhikistan and were considered only by Victorov to belong to C.subg.Scapiflorae. They were not included in our phylogenetic analyses but with their branched inflorescences and Middle Asian distribution we postulate that they are certainly not closely related to the Tridentatae lineage.
The Caucasian endemic Campanulapetrophila is nested in clade CAM 17C2 sister to C.raddeana, another Caucasian endemic species, and together with C.latifolia, the type of the name Campanula, C.trachelium and C.sclerotricha (Fig. 1, see also Suppl. material 1). C.petrophila and C.raddeana have similar ecological patterns like growing only in vertical rocks, but somewhat differ in leaf shape and develop slightly branched inflorescence.
Campanulaczerepanovii Fed. is a little known local endemic of Dagestan with branched inflorescences and was included in the Scapiflorae group only by Kharadze (1949). It has not been included in any phylogenetic study but is certainly no member of C.sect.Tridentatae, because of its different stem morphology.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Boissier E (1875) Flora Orientalis 3. H. Georg, Geneva & Basel.
- 2Borsch T Hilu KW Quandt D Wilde V Neinhuis C Barthlott W (2003) Noncoding plastid trn T‐trn F sequences reveal a well resolved phylogeny of basal angiosperms.Journal of Evolutionary Biology 16(4): 558–576. 10.1046/j.1420-9101.2003.00577.x 14632220 · doi ↗ · pubmed ↗
- 3Candolle AP de (1830) Monographie de Campanulacées. Veuve Desray, Paris. 10.5962/bhl.title.111415 · doi ↗
- 4Darriba D Taboada GL Doallo R Posada D (2012) j Model Test 2: More models, new heuristics and parallel computing.Nature Methods 9(8): 772–772. 10.1038/nmeth.2109 PMC 459475622847109 · doi ↗ · pubmed ↗
- 5Dolukhanov AG (1966) Vegetation cover. In: Gerasimov IP (Ed.) The Caucasus.Nauka, Moscow, 223–255. [in Russian]
- 6Droege G Barker K Astrin J Bartels P Butler C Cantrill D Seberg O (2014) The Global Genome Biodiversity Network (GGBN) data portal. Nucleic Acids Research 42(D 1): 607–612. 10.1093/nar/gkt 928PMC 396510624137012 · doi ↗ · pubmed ↗
- 7Fedorov AA (1957) Campanulaceae. In: Schischkin BK (Ed.) Flora SSSR (Vol.24). Academy of Sciences of the USSR, Moscow, 126–450.
- 8Fomin AV (1905) Campanulaceae. In: Kuznetsov N Busch N Fomin A (Eds) Матеріалы Флора кавказа [Flora Caucasica Critica] 4(6).– Jurjev: K. Mattison, 6–157.