Sodium Metabisulfite Inhibits Acanthamoeba Trophozoite Growth through Thiamine Depletion
Ronnie Mooney, Elisa Giammarini, Erin Corbett, Scott Thomson, Kevin McKinley, Paula Sinisterra Sebastian, Kiri Rodgers, Jana O’Donnell, Charles McGinness, Craig W. Roberts, Kanna Ramaesh, Fiona L. Henriquez

TL;DR
Sodium metabisulfite, found in eye drops, inhibits Acanthamoeba growth by depleting thiamine, a nutrient the organism cannot make itself.
Contribution
The study identifies sodium metabisulfite as a novel inhibitor of Acanthamoeba trophozoites through thiamine depletion.
Findings
Sodium metabisulfite inhibits Acanthamoeba trophozoites at low concentrations (IC50 0.03 mg/mL).
Acanthamoeba is a thiamine auxotroph, requiring external thiamine for growth.
Thiamine supplementation reverses the inhibitory effects of sodium metabisulfite.
Abstract
Acanthamoeba keratitis (AK) is a severe infection of the cornea. Prevention and treatment are difficult due to the inefficacy of currently available compounds. The impact of many commonly used compounds for routine examinations of Acanthamoeba is unexplored but might offer insight useful in combatting AK. In this study, we demonstrate that sodium metabisulfite, a common preservation constituent of eye care solutions, was found to be active against Acanthamoeba trophozoites at concentrations lower than that commonly found in eye drops (IC50 0.03 mg/mL). We demonstrate that sodium metabisulfite depletes thiamine from growth medium and that Acanthamoeba is a thiamine auxotroph, requiring thiamine salvage for growth. The inhibitory effects of sodium metabisulfite can be overcome by thiamine supplementation. These results are consistent with the lack of key enzymes for thiamine biosynthesis…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
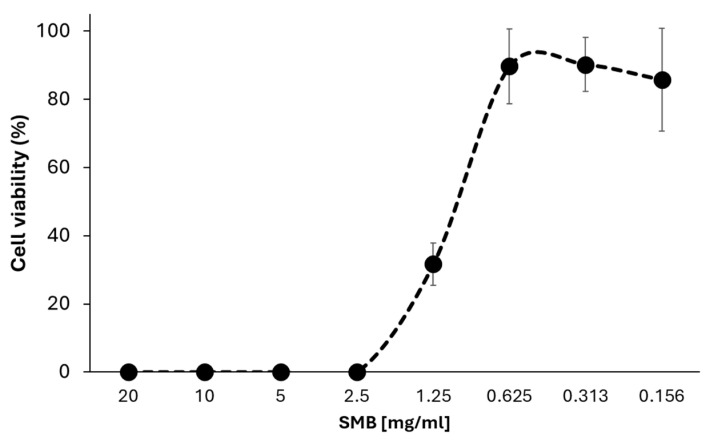 Figure 1
Figure 1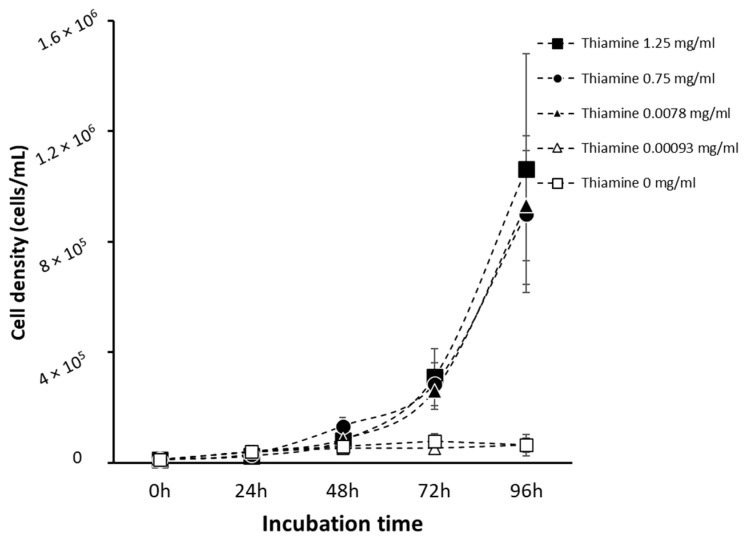 Figure 2
Figure 2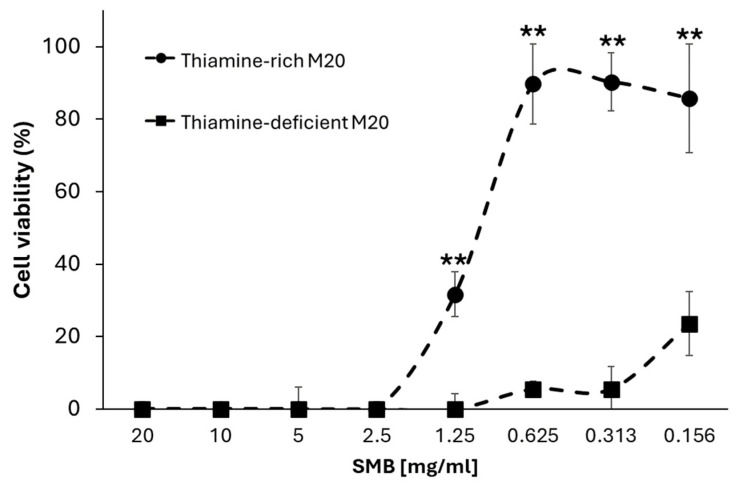 Figure 3
Figure 3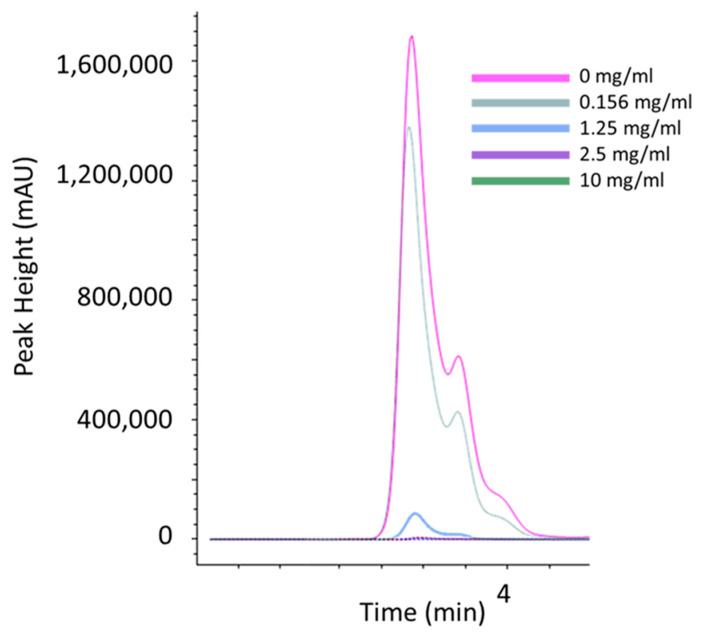 Figure 4
Figure 4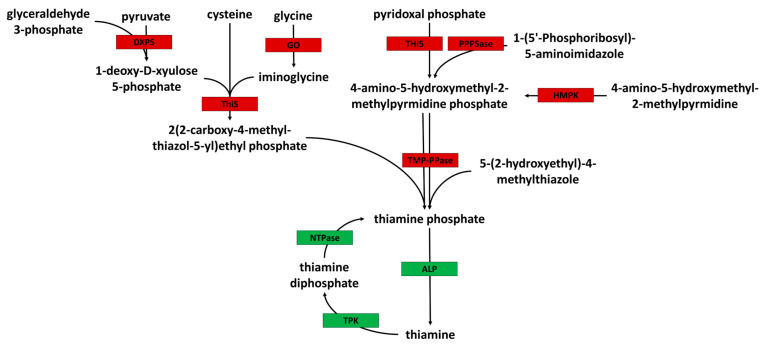 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAlcoholism and Thiamine Deficiency · Legionella and Acanthamoeba research · Heme Oxygenase-1 and Carbon Monoxide
1. Introduction
Acanthamoeba keratitis (AK) is an infection that affects between 1 and 9 people per 100,000 contact lens wearers [1], with incidence rates increasing in several countries globally [2,3,4,5]. Patients typically present with severe eye pain, eye redness, blurred vision, sensitivity to light, and excessive tearing [6]. Clinical examination of the infected area necessitates the use of solutions containing mydriatic agents such as phenylephrine hydrochloride eye drops, and confirmation of AK is determined using culture-based, molecular, and microscopic analysis of patient lens cases or corneal scrapings [6,7]. The impact of routine compounds used in initial clinical examinations of Acanthamoeba spp. is as yet undescribed, but further investigation is warranted to inform diagnostic and therapeutic approaches and improve patient outcome.
Delayed diagnosis due to inconclusive results or failure of polymerase chain reaction (PCR) and/or culture to detect infection negatively impact the likelihood of resolving AK infections without surgical intervention [6,8]. Currently, treatment regimens for AK are arduous, typically consisting of prolonged combination therapies using polyhexamethylene biguanide (PHMB) or chlorhexidine and a diamidine such as propamidine or hexamidine [8]. Despite this, infection can be non-responsive to treatment, and corneal transplantation is often used to clear infection [9]. Identifying new preventative or therapeutic compounds to ease the burden of AK is important, and elucidating the role of existing compounds is useful in that it provides opportunity for compound repurposing or opens avenues for synergistic combinations to be trialled. Indeed, eye drops containing phenylephrine hydrochloride that are frequently used to dilate the pupil and facilitate fundus examination often contain excipients such as sodium metabisulfite [10,11,12,13,14,15], shown to have antimicrobial activity against organisms such as Staphylococcus spp., Escherichia coli, Candida albicans, and Aspergillus flavus [16,17] and widely used as a food preservative [18,19,20,21]. The antimicrobial mode of action of sodium metabisulfite is unclear, but it has been suggested that sulfite can react with various biological molecules. Interestingly, sodium metabisulfite has been shown to degrade thiamine in solutions [22], potentially limiting thiamine availability, resulting in an indirect antimicrobial effect. Thiamine is an essential micronutrient and serves as a co-factor for many enzymatic reactions involved in energy production and the synthesis of downstream metabolites [23]. Here, we investigate the antimicrobial activity of these compounds against Acanthamoeba at concentrations comparable to those found in commonly used eye drop solutions (e.g., phenylephrine hydrochloride; 1 mg/mL sodium metabisulfite).
This research demonstrates that sodium metabisulfite, a constituent of eye care products, can restrict the growth of A. castellanii. Through culture growth assays, bioinformatic analysis, and LC-MS analysis we demonstrate that Acanthamoeba is a thiamine auxotroph, scavenging this nutrient from the environment. The data presented here provide a platform from which potential new antimicrobial therapies and preventatives can be developed and incorporated into synergistic combinations that aid in the prevention and treatment of those suffering with AK. Furthermore, the inhibitory effect of sodium metabisulfite has the potential to influence and delay accurate diagnosis of the disease, ultimately affecting disease outcome, and so should be considered during initial screening.
2. Materials and Methods
2.1. Acanthamoeba Species
Acanthamoeba castellanii (ATCC 50370) trophozoites were maintained in Proteose Peptone and Glucose (PG)-supplemented medium [24]. Prior to experimental work, trophozoites were transferred to the chemically defined medium M20 [25] containing a thiamine concentration of either 1.25 mg/mL [25] or 0.0078 mg/mL (minimum concentration to support growth) and incubated at 25 °C for four days to allow cell acclimatisation.
2.2. Determination of Minimum Thiamine Concentration for A. castellanii Growth
A. castellanii trophozoites (1 × 10^4^ cells/mL) were seeded in chemically defined medium M20 [25] lacking thiamine or with thiamine concentrations of 1.25 mg/mL, 0.75 mg/mL, 0.0078 mg/mL, or 0.00093 mg/mL, and incubated at 25 °C. Growth was monitored by enumerating A. castellanii trophozoites using a haemocytometer (BlauBrand—Brand GMBH, Wertheim, Germany) and the data were recorded daily in a 24 h interval for 4 days (96 h in total).
2.3. Sodium Metabisulfite Acanthamoeba Inhibition Assay
A. castellanii trophozoites (1 × 10^5^ cells/mL) were seeded in standard M20 (with 1.250 mg/mL thiamine) [25] or modified M20 (with 0.0078 mg/mL thiamine). Sodium metabisulfite (40 mg/mL) (Sigma, Poole, UK) was added at a starting concentration of 20 mg/mL and serially diluted to a final concentration of 0.156 mg/mL and incubated for 96 h at 25 °C. Acanthamoeba viability was assessed by Alamar Blue™ (Thermo Fisher Scientific, Renfrew, UK) after the incubation time for a further 24 h [26].
2.4. Thiamine Chromatography
Sodium metabisulfite (Sigma Aldrich, Dorset, UK) was added to PG and M20 media in concentrations of 10 mg/mL, 2.5 mg/mL, 1.25 mg/mL, and 0.156 mg/mL and to the 10 ppm thiamine standard for a final sodium metabisulfite concentration of 10 mg/mL. Thiamine, thiamine/sodium metabisulfite, media only, and media/sodium metabisulfite solutions were incubated at room temperature for 24 h in the absence of light. An Agilent 1260 Infinity HPLC coupled to an Agilent 6120 Quadrupole MS was used for all analyses. Agilent OpenLab software v2.8 was used for instrument control and data analysis. Separation of analytes was performed on a Gemini 5u C18 110A column (150 × 4.6 mm, 5 µm particle size) (Phenomenex, Macclesfield, UK). Column temperature was set to 30 °C. The mobile phase was 20 mM ammonium acetate and 0.1% v/v forming acid in water at a flow rate of 1 mL/min, and sample injection volume was 20 µL. Total run time of the method was 8 min. The ion source was atmospheric pressure ionisation—electrospray (API-ES) in positive mode. Spray chamber conditions were as follows: drying gas flow—12 L/min; nebuliser pressure—35 psig; drying gas temperature—250 °C; capillary voltage—3000 V. Experiments were performed in single-ion monitoring (SIM) mode, with a target SIM ion m/z 265.1 and dwell time of 290 ms.
2.5. In Silico Analysis
The protein database (www.ncbi.nlm.nih.gov, accessed on 16 February, 2024) and the genome database for A. castellanii (www.amoebadb.org, accessed on 16 February, 2024) were mined for proteins involved in thiamine biosynthesis. Names of characterised proteins involved in thiamine biosynthesis from other kingdoms were used as queries for BLAST searches. Hits with low expectation values (<0.01) were subjected to further back-BLAST to assign putative functions by mining the NCBI protein database for sequences closely aligned to those available on Amoebadb. If back-BLAST results were in agreement with the queried protein but had an E-value below 0.01 and/or a percentage identical score below 40%, sequences were aligned using the Muscle alignment tool [27] and visualised using Jalview v2.11.3.2 [28,29]. Upon alignment, sequences were screened for the presence of active sites, and proteins lacking key active sites were considered false.
2.6. Statistical Analysis
Statistical significance between concentrations was determined by using a two-sided, two-sample t-test with a type I error rate of 0.05 in Microsoft Excel. ANOVA single-factor analysis, with alpha = 0.01, was conducted in Microsoft Excel, based on the cell viability values obtained. For all work undertaken in this study, a total of 3 independent experiments were conducted, with each independent experiment performed in triplicate.
3. Results
3.1. Sodium Metabisulfite Inhibits the Growth of A. castellanii Trophozoites
Sodium metabisulfite is a constituent of several eye care solutions [11,12,13,14,15] and has been shown to have antimicrobial properties in other organisms [16,17]. Further investigation examined the effect of different concentrations of this compound on Acanthamoeba growth (Figure 1). Sodium metabisulfite was found to inhibit the growth of Acanthamoeba with an IC_50_ of 1.21 mg/mL (Figure 1).
3.2. Acanthamoeba Trophozoites Require Thiamine for Growth
Sodium metabisulfite has been shown to degrade thiamine in infusion solutions [22]. To assess the role of thiamine depletion on A. castellanii, it is necessary to define the minimum concentration of thiamine required to support Acanthamoeba growth. Trophozoites (1 × 10^4^ cells/mL), seeded in M20 medium with varying concentrations of thiamine (Figure 2), were monitored daily for 96 h. Cell counts show a significant increase in the growth of A. castellanii at thiamine concentrations above 0.0078 mg/mL (p < 0.05). Cell growth was significantly inhibited at lower concentrations of thiamine (p < 0.05), suggesting a reliance on exogenous thiamine for cell growth (Figure 2). Growth medium containing 0.0078 mg/mL of thiamine was established as a minimal medium capable of sustaining Acanthamoeba growth.
3.3. Sodium Metabisulfite Inhibits Acanthamoeba Growth through Degradation of Thiamine
A. castellanii trophozoites were seeded in either thiamine-rich (thiamine exceeding growth requirements) M20 medium (1.25 mg/mL) or thiamine-minimal M20 medium (0.0078 mg/mL) for 96 h with doubly diluted concentrations of sodium metabisulfite (from 20 mg/mL to 0.156 mg/mL). Sodium metabisulfite inhibited A. castellanii trophozoites in a dose-dependent manner in both medium types (Figure 3). Amoebae incubated in thiamine-rich M20 showed a significantly higher tolerance to sodium metabisulfite (40.3-fold higher, p < 0.01) than those in thiamine-minimal M20 (Figure 3; IC_50_: 1.21 mg/mL and 0.03 mg/mL, respectively), suggesting that thiamine depletion is in part responsible for sodium metabisulfite efficacy against A castellanii. Supporting this observation, the thiamine levels in thiamine-rich M20 medium (Figure 4 and Figure S2a), PG medium (Figure S2b), and a thiamine standard (Figure S2c) were depleted in the presence of sodium metabisulfite in a dose-dependent manner.
3.4. Acanthamoeba Is a Thiamine Auxotroph
Detailed in silico analysis of the A. castellanii genome was carried out to identify enzymes necessary for the de novo synthesis of thiamine (Table S1). The data suggest that Acanthamoeba is unable to synthesize thiamine de novo. In brief, BLAST analysis suggests that Acanthamoeba does not possess a thiamine phosphate pyrophosphorylase enzyme necessary for the synthesis of thiamine phosphate from 4-amino-5-hydroxymethyl-2-methylpyrmidine diphosphate together with either 2(2-carboxy-4-methyl-thiazol-5-yl)ethyl phosphate or 5-(2-hydroxyethyl)-methylthiazole. However, alkaline phosphatase enzymes were identified (ACA1_194910 and ACA1_182290) for the conversion of thiamine phosphate to thiamine (Figure 5), as were thiamine pyrophosphokinase enzymes necessary for the conversion of thiamine to the active thiamine diphosphate (ACA1_225950 and ACA1_050760). Despite this, genome mining revealed no evidence of the cysteine-dependent adenosine diphosphate thiazole synthase enzyme required for the synthesis of 2(2-carboxy-4-methyl-thiazol-5-yl)ethyl phosphate from glycine and the thiazole synthase and thiazole tautomerase enzymes necessary for the synthesis of 2(2-carboxy-4-methyl-thiazol-5-yl)ethyl phosphate from iminoglycine, 1-deoxy-D-xyulose 5-phosphate, and cysteine. More significantly, we found no evidence of a hydroxymethylpyrimidine kinase gene in the A. castellanii genome, essential in the synthesis of the 4-amino-5-hydroxymethyl-2-methylpyrmidine diphosphate metabolite from 4-amino-5-hydroxymethyl-2-methylpyrmidine phosphate (Figure 5), with the putatively annotated gene ACA1_061800 lacking almost all functional sites described in the literature (Figure S1) [30].
4. Discussion
Prevention and treatment of AK is difficult due to the relatively limited understanding of the infectious organisms’ biochemical requirements. Thus, expanding this knowledge can aid in the development of a more targeted approach. The impact of many commercially available compounds on Acanthamoeba is yet to be explored, yet understanding these interactions provides a potential route for compound repurposing using solutions already deemed safe for use in humans [31]. Compounds commonly used for mydriasis often contain sodium metabisulfite [11,12,13,14,15] and have been shown to possess antimicrobial activity in other organisms [16,17]. Here, we have provided evidence to demonstrate that this compound can also reduce Acanthamoeba trophozoite viability after 96 h.
Sodium metabisulfite, a salt of sulphurous acid recognised by the European Commission under regulation 1129/2011, is extensively used for the preservation of fresh and dried fruit, vegetables, juices, and wines due to its antimicrobial properties [18,19,20,21]. Sodium metabisulfite has previously been shown to have antifungal (e.g., Aspergillus spp., Fusarium spp., Rhizoctonia solani) and antibacterial (e.g., Staphylococcus spp., Escherichia coli, Pseudomonas aeruginosa, Bacillus spp.) capabilities [32,33,34,35,36]. The broad-spectrum antimicrobial activity of sodium metabisulfite and its use in eye care solutions warranted further investigation into its activity against Acanthamoeba species. To the best of our knowledge, this is the first study investigating the antimicrobial activity and potential mode of action of sodium metabisulfite against any Acanthamoeba species. The activity of sodium metabisulfite against A. castellanii trophozoites was initially investigated using standard M20 medium [25], containing 1.25 mg/mL of thiamine, and a concentration of 1.21 mg/mL of sodium metabisulfite was necessary to reduce cell viability by 50%. To further elucidate the mechanisms by which sodium metabisulfite acted on Acanthamoeba, we built on work previously undertaken by Scheiner and colleagues [22], in which sodium metabisulfite was shown to reduce the availability of thiamine in infusion solutions. Thiamine is an essential micronutrient and is converted to the active thiamine diphosphate via the activity of thiamine pyrophosphokinase, at which point it can act as a co-factor for enzymes necessary for energy production and normal cellular function [37]. For example, the tricarboxylic acid (TCA) cycle-associated enzymes pyruvate dehydrogenase and 2-oxoglutarate dehydrogenase, necessary for the conversion of pyruvate to acetyl-CoA and pyruvate dehydrogenase to succinyl-CoA, respectively, are reliant on thiamine diphosphate as a co-factor [37,38,39]. Similarly, fatty acid degradation and the pentose phosphate pathway also rely on this co-factor for the synthesis of essential metabolites or as a means of producing energy through α-oxidation and β-oxidation, or glycolysis [37,40,41]. We noted that the depletion of available thiamine in growth media resulted in a decrease in cell growth, with no growth observed in M20 medium containing less than 0.0078 mg/mL of thiamine, suggesting that Acanthamoeba was highly reliant on thiamine supplementation and perhaps lacked one or more necessary proteins for the de novo synthesis of the compound. Indeed, while our in silico analysis of the Acanthamoeba genome did suggest that it possesses thiamine pyrophosphokinase enzymes for the synthesis of thiamine diphosphate, we found no evidence of protein sequences corresponding to essential upstream enzymes, for example, hydroxymethylpyrimidine kinase or thiamine phosphate pyrophosphorylase, necessary for de novo synthesis of thiamine [42], which supported our finding that the scavenging of thiamine from the environment is essential for cell function.
In agreement with Scheiner et al. [22], we found that supplementing M20 medium with sodium metabisulfite resulted in a dose-dependent decrease in thiamine. To demonstrate that this interaction was influencing cell viability, the same cytotoxicity assay was performed on cells incubated in M20 medium containing the minimal thiamine concentration necessary for survival (0.0078 mg/mL). A 40-fold decrease in the IC_50_ was noted relative to the thiamine-rich medium used initially (0.03 mg/mL and 1.21 mg/mL, respectively). The results of this study suggest that sodium metabisulfite acts on Acanthamoeba through the depletion of available thiamine. It is interesting to note that the concentrations of sodium metabisulfite capable of inhibiting A. castellanii growth were comparable to those listed in commonly used eye care solutions (1 mg/mL), suggesting it may have use in the prevention of AK, perhaps through integration into contact lens solutions, particularly given the lack of toxicity in ocular studies [31]. Conversely, it could be speculated that the activity of the compound on Acanthamoeba may influence diagnosis, with mydriatic agents containing sodium metabisulfite used for initial diagnostics perhaps reducing the culturability of isolated samples. In both instances, significantly more research is required.
In summary, this study highlights that excipients in eye care solutions such as sodium metabisulfite can have inhibitory effects on Acanthamoeba trophozoites that could be exploited in preventative or therapeutic strategies against AK. It should also be considered that use of these compounds during initial diagnostics could interfere with the accurate detection of Acanthamoeba. Further understanding of how these constituents impact amoeba growth and how these compounds act collectively is important, and the synergy of compounds such as sodium metabisulfite with other widely used compounds should be assessed. Overall, we have demonstrated that sodium metabisulfite is an effective inhibitor of A. castellanii growth and that the organism lacks the molecular tools necessary for de novo thiamine synthesis, presenting a potential area for future drug targeting.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Somani S.N. Ronquillo Y. Moshirfar M. Acanthamoeba Keratitis Stat Pearls Publishing Treasure Island, FL, USA 202431751053 · pubmed ↗
- 2Scruggs B.A. Quist T.S. Salinas J.L. Greiner M.A. Notes from the Field: Acanthamoeba Keratitis Cases—Iowa, 2002–2017 MMWR Morb. Mortal. Wkly. Rep.20196844844910.15585/mmwr.mm 6819 a 631095535 PMC 6522078 · doi ↗ · pubmed ↗
- 3Höllhumer R. Keay L. Watson S.L. Acanthamoeba Keratitis in Australia: Demographics, Associated Factors, Presentation and Outcomes: A 15-Year Case Review Eye 20203472573210.1038/s 41433-019-0589-631554948 PMC 7093427 · doi ↗ · pubmed ↗
- 4Randag A.C. van Rooij J. van Goor A.T. Verkerk S. Wisse R.P.L. Saelens I.E.Y. Stoutenbeek R. van Dooren B.T.H. Cheng Y.Y.Y. Eggink C.A. The Rising Incidence of Acanthamoeba Keratitis: A 7-Year Nationwide Survey and Clinical Assessment of Risk Factors and Functional Outcomes P Lo S ONE 201914 e 022209210.1371/journal.pone.022209231491000 PMC 6731013 · doi ↗ · pubmed ↗
- 5Carnt N. Hoffman J.J. Verma S. Hau S. Radford C.F. Minassian D.C. Dart J.K.G. Acanthamoeba Keratitis: Confirmation of the UK Outbreak and a Prospective Case-Control Study Identifying Contributing Risk Factors Br. J. Ophthalmol.20181021621162810.1136/bjophthalmol-2018-31254430232172 · doi ↗ · pubmed ↗
- 6Lorenzo-Morales J. Khan N.A. Walochnik J. An Update on Acanthamoeba Keratitis: Diagnosis, Pathogenesis and Treatment Parasite 2015221010.1051/parasite/201501025687209 PMC 4330640 · doi ↗ · pubmed ↗
- 7Szentmáry N. Daas L. Shi L. Laurik K.L. Lepper S. Milioti G. Seitz B. Acanthamoeba Keratitis—Clinical Signs, Differential Diagnosis and Treatment J. Curr. Ophthalmol.201931162310.1016/j.joco.2018.09.00830899841 PMC 6407156 · doi ↗ · pubmed ↗
- 8Fanselow N. Sirajuddin N. Yin X.-T. Huang A.J.W. Stuart P.M. Acanthamoeba Keratitis, Pathology, Diagnosis and Treatment Pathogens 20211032310.3390/pathogens 1003032333801905 PMC 7998329 · doi ↗ · pubmed ↗