The Enigma of NTH2 Gene in Yeasts
Sergi Maicas, Ruth Sánchez-Fresneda, Francisco Solano, Juan-Carlos Argüelles

TL;DR
This paper investigates the role of the NTH2 gene in yeasts, finding it does not encode a functional trehalase enzyme despite genomic predictions.
Contribution
The study provides biochemical evidence that the NTH2 gene in some yeasts does not produce a functional neutral trehalase.
Findings
Nth2p lacks measurable neutral trehalase activity in S. cerevisiae.
Disruption of the NTH2 gene does not affect neutral trehalase activity or cause significant phenotypic changes.
No duplicated NTH gene has been found in other yeast species like C. albicans or C. parapsilosis.
Abstract
The enzymatic hydrolysis of the non-reducing disaccharide trehalose in yeasts is carried out by trehalase, a highly specific α–glucosidase. Two types of such trehalase activity are present in yeasts, and are referred to as neutral and acid enzymes. They are encoded by distinct genes (NTH1 and ATH1, respectively) and exhibit strong differences in their biochemical and physiological properties as well as different subcellular location and regulatory mechanisms. Whereas a single gene ATH1 codes for acid trehalase, the genome of some yeasts appears to predict the existence of a second redundant neutral trehalase, encoded by the NTH2 gene, a paralog of NTH1. In S. cerevisiae the corresponding two proteins share 77% amino acid identity, leading to the suggestion that NTH2 codes for a functional trehalase activity. However, Nth2p lacks any measurable neutral trehalase activity and disruption…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
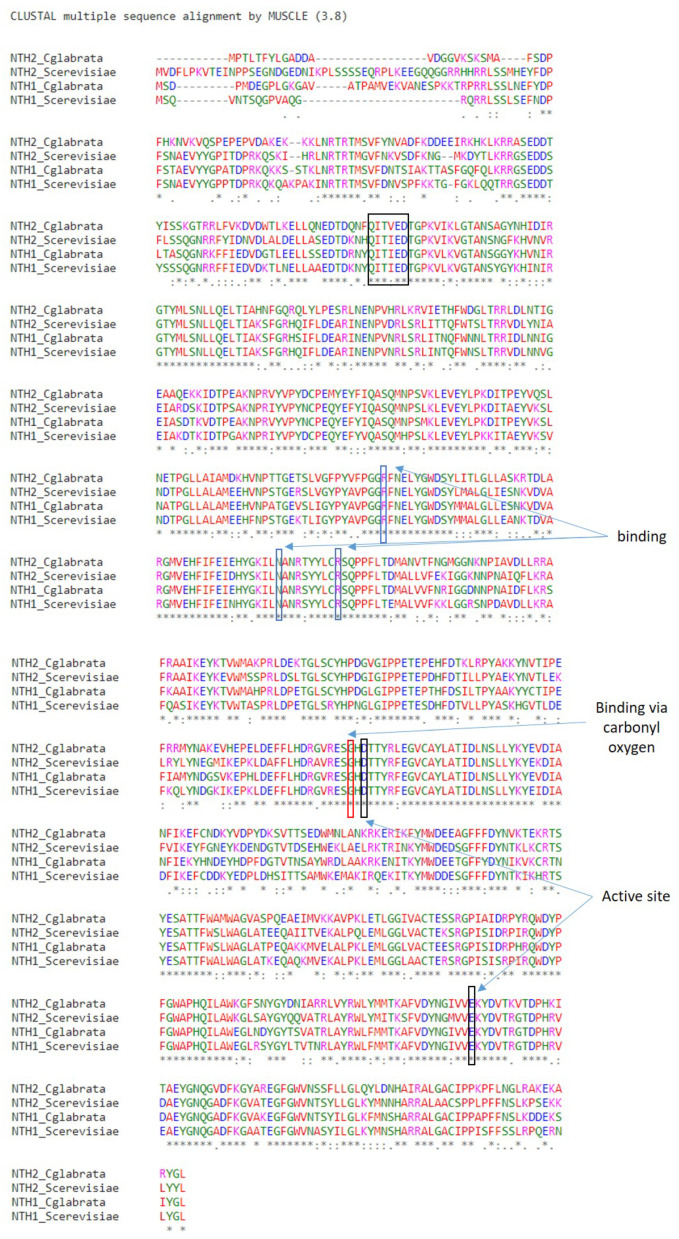 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal and yeast genetics research · Biofuel production and bioconversion · Polysaccharides and Plant Cell Walls
1. Introduction
Trehalose is a widely conserved and physiologically relevant non-reducing disaccharide which consists of two glucose molecules linked by an (1,1) bond. This sugar plays a crucial role in various biological processes that are essential for maintaining cellular homeostasis in numerous organisms [1]. The importance of trehalose is underscored by its involvement in cell protection against both nutritional and environmental stress conditions (i.e., starvation, dehydration, heat or freezing). This success can largely be explained by its chemical structure and singular physical properties, as it is the thermodynamically and kinetically most stable disaccharide in the biosphere [1,2,3]. Furthermore, recent intensive research also suggests promising applications of trehalose in food, cosmetic or clinical therapy [1]. Notably, trehalose is undergoing intensive investigation for its relevant neuroprotective effects, particularly when it is used to treat devastating neurodegenerative diseases like Huntington’s, Alzheimer’s or Parkinson’s disease [1]. Indeed, trehalose is able to cross the blood–brain barrier and may induce direct protection of proteins prone to forming aggregates, and it could be involved in the activation of autophagy [1]. In most organisms, hydrolysis of trehalose is carried out by enzymes known as trehalases (E.C. 3.2.1.28). These are a kind of -glucosidase rigorously specific to trehalose as a sole substrate, which is cleaved off in two molecules of glucose [3,4,5]. Several prokaryotes and eukaryotes display two mechanistic exceptions to this rule: (i) trehalose phosphorylase, which catalyzes the phosphorolysis of trehalose to produce glucose-1-phosphate and glucose, and (ii) phosphotrehalose that converts the intermediate trehalose-6-phosphate into glucose and glucose-6-phosphate [3,4,5].
Using yeasts as model organisms, an important distinction can be envisaged between two key physiological processes that require trehalose hydrolysis. The first course is the rapid mobilization of metabolic-responsive endogenous trehalose, which serves as an internal energy reserve that is crucial to the cell’s rapid response to sudden demands. This form of trehalose serves as an internal energy reserve that is crucial to the cell’s rapid response to metabolic demands, particularly under stress conditions like nutrient scarcity or environmental stress. The second process is the utilization of exogenous free trehalose, which may be available in the yeast’s environment. After hydrolysis, the released glucose can be imported into the cells and further used either as an energy source or for other physiological activities. This external trehalose can be imported into the cell and used as an energy source or for other metabolic functions. These two processes are facilitated by two distinct trehalases, each encoded by different, unrelated genes. These trehalases exhibit significant biochemical and genetic differences. One of the key differences lies in their subcellular locations: the trehalase responsible for mobilizing endogenous trehalose is typically found in the cytosol, whereas the trehalase involved in utilizing exogenous trehalose is often associated with the cell membrane or other compartments. Moreover, these trehalases differ in their catalytic parameters. The cytosolic trehalase generally has a higher affinity for trehalose and is regulated in a way that allows a rapid response to internal metabolic cues. In contrast, the membrane-associated trehalase may have different kinetic properties, which are optimized for processing external sources of trehalose that are less consistently available. Regulatory controls also vary between these trehalases. The endogenous trehalase is tightly regulated by intracellular signals that reflect the cell’s metabolic state, ensuring that trehalose is mobilized when the cell requires energy. On the other hand, the exogenous trehalase is regulated in a manner that aligns with the availability of trehalose in the environment, allowing the cell to take advantage of external nutrient sources when present. This differentiation between trehalase functions highlights the complex regulatory networks that enable yeast cells to efficiently manage their energy resources and adapt to fluctuating environmental conditions. Understanding these mechanisms not only provides insights into yeast metabolism but also offers broader implications for the study of energy regulation and stress responses in other organisms [3,5,6]. They only share a strict specificity for trehalose as a substrate and are usually distinguished as cytosolic or neutral trehalases (Nth1p/Nct1p), which are engaged in the mobilization of endogenous trehalose, which is activated by some divalent cations (Mg^2+^ and Ca^2+^) and regulated by reversible phosphorylation mediated by cAMP-dependent protein kinases. In contrast, the so-called acid trehalases (Ath1p/Atc1p) are cell-linked external or vacuolar enzymes that are responsible for cell growth on exogenous trehalose and subjected to glucose repression [3,4,5].
It should be mentioned, however, that in some yeasts that are considered important for biotechnological processes, such as Saccharomyces cerevisiae, which is used in the production of bioethanol and other fermentation products, and in yeasts relevant as human pathogens, such as Candida albicans, which can cause opportunistic infections, a second gene known as NTH2 has been described. This gene codes for a putative neutral trehalase.
This marked distinction between the functions of trehalase–trehalose systems in fungi highlights the complex regulatory networks that enable fungal cells to efficiently manage their energy resources and adapt to fluctuating environmental conditions. A precise understanding of the mechanisms involved would not only provide new insights into fungal metabolism, but would also have broader implications for the study of energy regulation and stress-response circuits in other organisms [1,3,4,5]. In this way, the main features and putative relationships between both trehalases have been exhaustively revised and will not be analyzed here [3,4,5]. It should be mentioned, however, that in some yeasts that are considered important for biotechnological processes (i.e., Saccharomyces cerevisiae utilized in the production of bioethanol and other fermentation products) as well as in yeasts that are relevant as human pathogens (i.e., Candida albicans and other “non-albicans” species of Candida that cause both superficial and systemic opportunistic infections) a second gene (NTH2) has been described, coding for a putative neutral trehalase. Notably, despite its identification, the main features and biological roles of which remain elusive [2,7].
2. Does the NTH2 Gene Code for a Functional Neutral Trehalase in Yeasts?
Unlike acid trehalases, where a unique gene (ATH1) codes for acid trehalase, a search in the genome databases predicts the existence of a redundant neutral trehalase activity in several yeasts, encoded by the NTH2 gene, a paralog of NTH1 [2,7,8]. In Saccharomyces cerevisiae the corresponding proteins appear to share 77% identity. While it has been proposed that Nth2p is a true functional trehalase that is directly involved in the mobilization of stored endogenous trehalose [2,7,8], we are inclined to believe that the available data are inconsistent with this statement. Indeed, much more experimental evidence would be necessary to firmly establish the above-indicated hypothesis. The following arguments support this reluctant view: Apart from in S. cerevisiae, the presence of two neutral trehalase activities has only been extensively demonstrated in the yeast Candida glabrata (Nth1p consists of 769 amino acids/87.4 kDa, whereas Nth2p contains 750 amino acids/86.5 kDa) [7]. This coincidence reinforces the suggestion that C. glabrata fits better into Saccharomycetaceae clade than in the Candida clade [9]. Notably, in S. cerevisiae the Nth2p enzyme shows a higher resemblance with its own Nth1p than with the ortholog Nth2p present in C. glabrata (Figure 1) Furthermore, the highest degree of homology (71% amino acids identity) corresponds to neutral trehalases from C. glabrata [7], while an elevated similarity also exists between the two Nth1 enzymes from these two yeasts (Figure 1). Furthermore, a comparison of sequences between NTH1 and NTH2 genes confirms they do not contain a transmembrane domain or a signal sequence (Figure 1) [3,5]. In addition, a careful database search (Swiss Prot) points to the presence of NTH2-related genes in the phytopathogenic filamentous fungi Eremothecium gossypii and in the lager yeast Saccharomyces pasterianus, which have not been characterized yet. Nevertheless, thus far, no evidence concerning this duplicated NTH2 gene has been found in other relevant yeasts. Thus, in the opportunistic pathogen C. albicans, only one gene appears to code for neutral trehalase, although there is still a need for further research [10]. Remarkably, the homozygous deletion of the single ATH1 and NTH1 genes completely abolishes trehalase activity in C. parapsilosis [11,12]. More intriguing is the observation that after precise catalytic measurements, the Nth2p has no neutral trehalase activity in S. cerevisiae and disruption of NTH2 gene does not modify this activity compared with a parental strain [2]. Likewise, single nth1 and double nth1 /nth2 null mutants show no detectable neutral activity [7,9]. It might be expected that, if NTH2 encodes a genuine functional neutral trehalase, it would compensate for the lack of Nth1p activity. In contrast, although overexpression of NTH2 in a nth1 mutant induces a certain increase in trehalose hydrolysis, the result is unclear, since acid trehalase may be operative in these cells [8,12]. Another important clue is the absence of a specific associated phenotype to disrupt NTH2. Thus, in S. cerevisiae an intact NTH2 gene appears to be required for cell recovery upon heat shock and saline stress [7,13]. As a rule, the phenotypes displayed by heterozygous nth1 /Nth2 and nth1 /nth2 null mutants appear quite similar, with a slight difference concerning the capacity of trehalose accumulation [8,14]. In turn, the role of Nth2p in C. glabrata is a mystery. In tests on virulence factors involved in gut colonization, NTH2 gene is upregulated inside macrophages and its deletion reduces cell survival, but it does not play a conspicuous role in this process [7], although in many pathogenic fungi, stored trehalose is an element of virulence [13,15].
3. Hypothetical Roles of NTH2 Genes in the Physiology of Yeasts
The information currently available does not allow the formulation of a convincing proposition concerning a main function of NTH2 gene in the physiology of yeasts. Some preliminary data suggest that it might have an important role in the fine-tuning of trehalose metabolism, possible by providing an additional level of regulatory control under specific nutritional and/or environmental conditions [2,4,8]. For instance, Nth2p might be involved in trehalose hydrolysis when yeast cells experience a certain type of abiotic stress or when there is a sudden need for energy that cannot be satisfied by the primary trehalase alone. Furthermore, the potential significance of NTH2 in biotechnological applications and pathogenicity adds an intriguing dimension to its extensive research. Thus, in many industrial yeast strains, a clear elucidation of the NTH2 function could lead to enhanced metabolic engineering strategies, improving the yield of bioethanol or other valuable bioproducts by optimizing the endogenous trehalose content. In turn, in pathogenic yeasts, this knowledge would provide new insight regarding the adaptative processes of these organisms to the host body during a productive infection, revealing new potentially interesting targets for antifungal therapies [1,2,3,5,8].
4. Conclusions and Perspectives
We conclude that although the sequence data seem to suggest that NTH2 gene might code for a neutral trehalase in yeasts, extensive and precise genetic and biochemical analysis do not support this proposal. Nth2p lacks any measurable trehalase activity, while disruption of NTH2 gene has no effect on both the endogenous trehalose content and its mobilization recorded in wild type cells. Furthermore, the nth2 null mutants do not display an evident phenotype, beyond a certain involvement in tolerance to heat shock and saline stress, but these observations require further confirmation. Other questions related to catalytic properties, regulatory mechanisms and physiological roles played by this putative neutral trehalase should also be readdressed. Further investigations should include gene knockout studies and in-depth biochemical characterization of Nth2p enzyme and the analysis of its expression patterns displayed by the distinct trehalases under various growth conditions. Insights gained from NTH2 research could promote the development of more robust industrial yeast strains or the obtention of novel more potent and safer antifungals, providing stronger support for the interconnection between basic research and applied sciences. Therefore, although NTH2 gene must play a physiological role in yeasts under still-unknown conditions, to date, it remains an enigma.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kuczynska-Wisnik D. Stojowska-Swedrzynska K. Lakswoska E. Intracellular protective functions and therapeutic potential of trehalose Molecules 202429208810.3390/molecules 2909208838731579 PMC 11085779 · doi ↗ · pubmed ↗
- 2Nwaka S. Kopp M. Holzer H. Expression and function of the trehalase genes NTH 1 and YBR 0106 in Saccharomyces cerevisiae J. Biol. Chem.1995270101931019810.1074/jbc.270.17.101937730323 · doi ↗ · pubmed ↗
- 3Sakaguchi M. Diverse and common features of trehalases and their contributions to microbial trehalose metabolism Appl. Microbiol. Biotechnol.20201041837184710.1007/s 00253-019-10339-731925485 · doi ↗ · pubmed ↗
- 4Nwaka S. Holzer H. Molecular biology of trehalose and the trehalases in the yeast Saccharomyces cerevisiae Prog. Nucleic Acid Res. Mol. Biol.199858197237930836710.1016/s 0079-6603(08)60037-9 · doi ↗ · pubmed ↗
- 5Maicas S. Sánchez-Fresneda R. Argüelles J.C. Yeast trehalase: Two enzymes, one catalytic mission Biochim. Biophys. Acta (BBA)-Gen. Subj.201618602249225410.1016/j.bbagen.2016.04.02027133444 · doi ↗ · pubmed ↗
- 6Sánchez-Fresneda R. González-Párraga P. Esteban O. Laforet L. Valentín E. Argüelles J.C. On the biochemical classification of yeast trehalases: Candida albicans contains two enzymes with mixed features of neutral and acid trehalase activities Biochem. Biophys. Res. Commun.20093839810210.1016/j.bbrc.2009.03.13419336219 · doi ↗ · pubmed ↗
- 7Van Ende M. Timmermans B. Vanreppelen G. Siscar-Lewis S. Fischer D. Wijnants S. Romero C.L. Yazdani S. Rogiers O. Demuyser L. The involvement of the Candida glabrata trehalase enzymes in stress resistance and gut colonization Virulence 20211232934510.1080/21505594.2020.186882533356857 PMC 7808424 · doi ↗ · pubmed ↗
- 8Jules M. Beltran G. François J.M. Parrou J.L. New insights into trehalose metabolism by Saccharomyces cerevisiae: NTH 2 encodes a functional cytosolic trehalase, and deletion of TPS 1 reveals Ath 1p-dependent trehalose mobilization Appl. Environ. Microbiol.20087460561410.1128/AEM.00557-0718065618 PMC 2227697 · doi ↗ · pubmed ↗