Fauna and Ecology of Macromycetes (Basidiomycota) in the Arctic Tree and Shrub Ecosystems of Central Siberia
Sergey Sergeevich Kulakov, Andrey Ivanovich Tatarintsev, Denis Aleksandrovich Demidko, Natalia Pavlovna Khizhniak

TL;DR
This study explores the diversity and ecological roles of fungi in Arctic tree and shrub ecosystems in Central Siberia, revealing insights into their biodiversity and function in permafrost regions.
Contribution
The study provides a detailed taxonomic and ecological analysis of macromycetes in under-researched Arctic ecosystems of Central Siberia.
Findings
Seventy species of macromycetes were identified, with mycorrhizal and wood decay fungi dominating (71%).
Fungal decomposers are prevalent in tundra areas due to the lack of woody substrates.
The taxonomic and ecological structure of Basidiomycota in Arctic ecosystems resembles that of taiga and temperate forests.
Abstract
The Arctic tree and shrub ecosystems of Central Siberia are distinctive in their ability to perform crucial biosphere functions. However, such forests have been the subject of relatively limited research. The understanding of the dynamics of these forests, including their composition and structure, is of significant relevance for the conservation of permafrost ecosystem biodiversity. Nevertheless, the species composition of the mycobiota of this region remain under investigation. The aim of the present study is to determine the macromycetes diversity in the main ecotopes of the Krasnoyarsk Arctic (Norilsk). An understanding of the ecological attributes of macromycetes within these ecosystems is essential for understanding of the decomposition of organic matter, the absorption of nutrients by trees and shrubs, and other fundamental ecological processes. Moreover, basidiomycetes may be…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4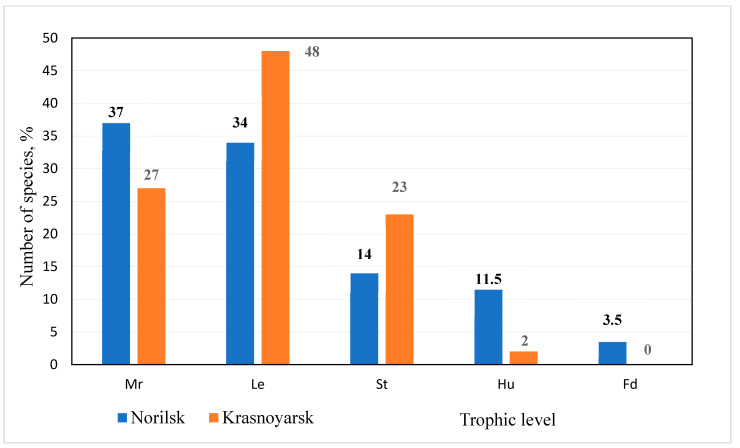 Figure 5
Figure 5- —Methodological bases for assessment of forest pathology risks in southern Central Siberia
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Lichen and fungal ecology · Forest Ecology and Biodiversity Studies
1. Introduction
Larch forests and woodlands (low-density forests where trees and shrubs form a light canopy) in the Siberian permafrost zone are unique. They perform crucial biosphere functions and are a source of various natural resources. Despite their wide geographical distribution, such forests have been studied unevenly. This is especially true for sub-tundra (boundary region between tundra and boreal forests) and northern taiga forest ecosystems. Notably, their ecological role is much more significant than their resource potential [1,2,3]. Understanding high-latitude Siberian forest dynamics, as well as their composition and structure, is of high relevance. For the conservation of permafrost ecosystem biodiversity, a scientific approach to their sustainable management and resource protection is needed [4]. Silvicultural features of such forests have been studied by many Russian researchers [5,6]. However, the species composition and structure of mycobiota of this region are still under research [7,8,9,10,11,12,13]. An understanding of the ecological attributes of macromycetes within these ecosystems is essential for gaining insight into their underlying processes, such as the decomposition of organic matter, the absorption of nutrients by trees and shrubs, and so forth.
The available literature does not provide complete information on the mycobiota of Siberia, especially its northern part. The most studied is the Altai-Sayan mountainous ecoregion [14,15,16,17,18,19,20,21,22]. Karatygin and colleagues provided an annotated list of fungi in “Russian Arctic Fungi” [7]. The book combines the research of multiple authors who studied the Russian North, including Central Siberia (Krasnoyarsk Krai). However, the book in question contains only fragmentary information about basidiomycetes for the region under study. The species composition of this taxonomic group is incomplete, and there is a paucity of information about their ecology. Notably, one of the study areas, described in “Russian Arctic Fungi”, is located 230 km north of Norilsk. Poor knowledge of the mycobiota of the Russian North makes it difficult to reveal patterns in the geographical distribution of fungi. The available data on the ecology of agaric fungi mostly refers to their caprophores, which have a narrower ecological amplitude than mycelium [23]. Furthermore, basidiomycetes may serve as indicator species in ecological studies. The presence of their fruiting bodies, which can be readily and rapidly identified, indicates the occurrence of specific processes within the plant community. Ascomycetes, which are represented in the Arctic tree and shrub communities by numerous species, are a less optimal choice due to the brief lifespan of their fruiting bodies. Another reason they are an inferior indicator species is that they are challenging to rapidly identify in the field. For this reason, we excluded Ascomycota from further consideration.
The aim of the present study is to determine the macromycetes diversity in the main ecotopes of the Krasnoyarsk Arctic (Norilsk). The subject of our investigation is the ecosystem of trees and shrubs. In this study, we concentrated on basidiomycetes, a group of fungi that includes numerous species closely related to trees and shrubs.
2. Material and Methods
2.1. Study Area
The study area belongs to the Norilsk-Evenki Ecoregion [24], covering an area between Nizhnyaya Tunguska in the south and Khatanga in the north. The most elevated part of the Central Siberian Plateau (table mountains and a plateau in the center) is also located in the study area. The vegetation is represented by mountain larch forests (forest floor dominated by reindeer moss and shrubs), woodlands, shrub communities at the tree line (alder and birch), and mountain tundra. There is no vegetation at the highest elevations.
The study area is located in the subarctic (boreal) climate zone. An Arctic air mass determines the climate of the region. The study area is dominated by sparse forests and tundra biomes due to continuous permafrost [25]. The number and nature of seasons are based on the polar day and polar night. In Norilsk, for example, polar day lasts 68 days, and polar night lasts 45 days. The warmest month is July (average monthly temperature is 14 °C) (Norilsk), and the coldest month is January (average monthly temperature is minus 25 °C) (Kayerkan). The amplitude of air temperature variations is about 87° [26,27]. The average annual precipitation ranges from 400 mm (Norilsk) to 700 mm (Talnakh). Rain/snow falls mainly in July–October; precipitation exceeds evaporation, generating wet soil conditions. The annual snow season typically lasts from October to May (240–260 days) [26]. Weather conditions during the growing season determine not only the abundance and diversity of agaricoid fungi but also their fruiting. Despite stressful environmental conditions, macromycetes do form fruit bodies [28,29,30,31,32,33].
Field studies were conducted at four 1 ha (100 × 100 m) research plots (RP), placed in the most typical biotopes for the study area. The research plots’ characteristics are given below.
Research plot I. Larch–alder forest (dominant tree species is Larix sibirica; dominant shrub species is Duschekia fruticosa). The field layer is dominated by horsetail ferns (mostly, Equisetum pratense) and herbs (Anthoxanthum alpinum prevail). The research plot is located 51 km west of Norilsk and 34 km west of Kayerkan: in the upper part of a southern convex–concave slope in the Bolgokhtokh River valley. The research plot I is located 50 m above sea level (Figure 1).
Research plot II. Larch woodland. Larix sibirica mean high is 5–6 m (low canopy density). Shrubs cover about 70% of the research plot (dominant species is Betula nana). The research plot is located 51 km west of Norilsk and 33 km west of Kayerkan: on the terrace of the Bolgokhtokh River valley. The relief is mosaic: 60% of the research plot is micro hills (covered by subshrubs and lichens), and 40% of the research plot is micro depressions (covered by sedges and mosses). The research plot II is located 76 m above sea level (Figure 2).
Research plot III. Birch–spruce–larch forest. The field layer is dominated by horsetail ferns (mostly, Equisetum pratense). The research plot is located 1.5 km southeast of Talnakh and 15 km northeast of Norilsk: on the watershed of the Talnakh and Listvyanka rivers. There are both hills and lowlands on the research plot. Surface depressions are swampy or filled with thermokarst lakes. The research plot III is located 46 m above sea level (Figure 3).
Research plot IV. Tundra covered by shrubs, subshrubs, and sedges. The research plot is located 6 km north of Talnakh and 23 km northeast of Norilsk: on the second above-floodplain terrace of the Kharaelakh River valley. The relief is represented by a gentle (3–4°) northwestern slope. The research plot IV is located 163 m above sea level (Figure 4).
Table 1 shows GPS coordinates for the research plots and brief species composition of studied plant communities.
2.2. Data Collection and Analysis
The material was collected in July–August 2018–2021 in the basin of the Bolgakhton and Norilsk rivers. The study was conducted using cruise method. Macromycetes were identified by their fruit bodies. The fruit bodies (basidiomas) were collected over a month at 10-day intervals. Common and easily identified fungi were recorded in a field diary. Specimens, that were difficult or impossible to identify in the field, were collected for the herbarium. Preparation of fungi specimens for deposit as herbarium followed standard methods [34,35,36]. Each fungi specimen was placed in a separate container and was assigned a unique number. For each herbarium specimen, we also recorded the following information: collection location, research plot number, habitat, frequency, and cover classes. A total of 104 herbarium specimens were collected.
Identification of species was carried out in the Laboratory of Forest Cultures, Mycology, and Phytopathology of Sukachev Institute of Forest of Siberian Branch of the Russian Academy of Sciences. Microstructure was studied on dried material using a Mikmed 2 microscope and a standard set of reagents (KOH 5%, Melzer’s reagent for determining the amyloid reaction). The studied samples are stored in the herbarium of Sukachev Institute of Forest SB RAS.
The trophic level and habitat [12,16,17,18] of the identified macromycetes were determined according to Kovalenko classification, information about belonging to a trophic group is given according to [37]:
I. Decomposers:
- -On plant litter—Fd (folia desecta);
- -On forest floor—St (stramentum);
- -On humus—Hu (humus);
- -On wood—Le (lignum epigaeum);
- -On undamaged wood—Lei (lignum epigaeum integrum);
- -On rotten wood—Lep (lignum epigaeum putridum);
- -On roots and wood buried in soil—Lh (lignum hupogaeum);
- -On mosses—M (muscu);
- -On fungi fruit bodies—Mm (macromycetes);
- -On excrements—E (excrementum).
II. Simbionts:
- -Mycorrhizal fungi—Mr (mycorrhiza).
III. Parasites:
- -Facultative parasites on trees and shrubs—P (parasitum).
To assess the distribution of macromycetes within ecotopes, generally accepted scales of frequency and cover were used [38]. The scales characterize the spatial arrangement of fruit bodies and provide a quantitative and qualitative assessment of the proportion that species take in a fungi community composition in various ecological and trophic groups.
Frequency classes:
- Very rare (rarissime): 1–2 sporocarps (localities).
- Rare (raro): from 3 to 10 localities.
- Rather frequent (saepe): more than 11 localities.
- Very frequent (saepissime): more or less evenly over the entire area.
Cover classes:
- Fruit bodies occur singly—1.
- Fruit bodies occur in small groups. The number of fruit bodies in a group varies from 10 to 50—2.
- Fruit bodies occur in large groups (from 50 to 100 or more) or distributed evenly over the entire area—3.
Fungal taxonomy and their scientific names are given in accordance with the system adopted in the 10th edition of the Ainsworth and Bisbee Dictionary of the Fung [39], as well as the Index Fungorum databases (http://www.indexfungorum.org accessed on 16 June 2024) [40] and Mycobank (http://www.mycobank.org accessed on 16 June 2024) [41].
3. Results
The present research resulted in identifying 70 species of macromycetes in the study area (Table 2). Most species (about 85%) were studied during the growing season of 2018 due to intensive basidioma formation.
Table 3 shows a list of fungi species, identified in the study area, indicating their trophic level, habitat, and frequency/cover classes.
Figure 5 shows a relative distribution of macromycete species (identified near Norilsk) by trophic-level groups. We also suggest using data on the trophic structure of macromycetes in the southern taiga forests (near Krasnoyarsk) for comparative analysis [42].
Table 4 shows a quantitative assessment of the species composition and trophic structure of macromycete communities in the study area.
4. Discussion
The findings of the study permit the formulation of generalizations regarding the basidiomycete flora of the study area and their ecological role in the tree and shrub communities of the Arctic.
The identified macromycetes (70 species) belong to 44 genera, 25 families, and 8 orders included in the subclass Agaricomycetidae (Table 2 and Table 3). The order Agaricales dominates by the number of species (34 species), followed by Russulales (14 species), Polyporales (9 species), and Boletales (8 species), making up a total of 93% of the species in the identified mycobiota. The remaining orders are represented by a few species of macromycetes (1–3 species). The six leading families, uniting 52% of the species of the studied biota, include Russulaceae (with the largest number of species—14), followed by Polyporaceae (6 species), Suillaceae, and Tricholomataceae (5 species each), Cortinariaceae and Strophariaceae (4 species each). The remaining families are represented by a smaller number of species, including 11 families represented by one species only.
The species of fungi we discovered is not endemic to the Arctic [42]. Our research indicates that they can thrive in cold climates, including in the northern regions of Central Siberia.
Macromycetes are trophically associated with vegetation: some directly interact with trees and shrubs, and some inhabit detritus, which is dominated by plant organic matter [43]. Thus, the species diversity of macromycetes largely depends on the vegetation diversity. Therefore, the largest number of fungi species were found within forest ecosystems (RP I and III), characterized by a richer species composition of plant communities (Table 4).
Mycorrhizal fungi and wood decay fungi dominate in the study area (Figure 5), which is also typical for the trophic structure of the macromycetes in the southern taiga [42] and temperate [23] forests. The relative number of species of these trophic levels in the northern latitudes (study area) is similar (37% and 34%, respectively), with a slight predominance of mycorrhizal fungi. In the southern taiga forests, wood decay fungi significantly exceed mycorrhizal fungi by the number of species [42]: 48% and 27%, respectively. The lower abundance of wood-associated fungal species compared to taiga forests (Figure 5) can be attributed to the smaller role of tree species in the communities under study and the simplified structure of the tree layer. Southern taiga forest communities, being of higher species diversity, provide a greater variety of wood habitat, compared to northern taiga forests and woodlands. Mycorrhizal and wood decay fungi play a crucial role in forest ecosystems. Mycorrhizal fungi form symbiotic relationships with the roots of trees, increasing their growth and productivity and contributing to an increased nutrient uptake (primarily carbon) by tree biomass. Wood decay fungi, on the other hand, contribute to carbon emissions by decomposing woody debris.
Mycorrhizal fungi occurred in all research plots (Table 4). In the larch woodland (RP II), they take 60% of the species composition of macromycetes. In typical northern taiga forest communities (RP I and III), mycorrhizal fungi take 39% and 31%, respectively. The most numerous among mycorrhizal fungi were species of the family Russulaceae (14 species), and the genus Lactarius (9 species) (Table 2 and Table 3). L. vietus (Fr.) Fr. is common, while L. repraesentaneus Britzelm. Pers. and L. pubescens Fr. occur less frequently. The number of species of the genus Russula is relatively small, among which there are species associated exclusively with birch and species with a wide range of symbiont partners. R. paludosa Britzelm is the most common species in all research plots. Most species are ecologically flexible and capable of living in various habitats since they can form mycorrhizae with a wide range of woody plants.
Wood decay fungi occur in those plant communities, where a tree layer is present (Table 3). Wood decay fungi take the largest proportion in the species composition of the mycobiota in the birch–spruce–larch forest (RP III), which is explained by increased habitat diversity relative to other forests. Special mention should go to Armillaria mellea s.l, which prefers a parasitic lifestyle but is also able to feed saprotrophically (facultative decomposers). There were also sterile conks of Inonotus obliquus (Fr.) Pilát found on some living trees. Specialist fungi, that are able to live in one habitat only, are rare. For example, the host range of Fomitopsis betulina (Bull.) and I. obliquus is restricted exclusively to birch species. Fomitopsis pinicola (Sw.) P. Karst, on the other hand, colonizes wood of many deciduous and coniferous species (sometimes on weakened living trees). Fruit bodies of this species, along with Bjerkandera adusta and Fomes fomentarius, are quite common in forest communities.
Fungal decomposers inhabiting plant litter, forest floor, and humus together include about 30% of the species in the macromycete biota of the study area (Figure 5), which is consistent with previously obtained data for the southern taiga [42] and temperate [23] forests. They act as an important link in the nutrient cycle and improve soil fertility by decomposing plant residues. Fungal decomposers often exhibit broad trophic specialization, so their belonging to ecological and trophic groups cannot always be determined accurately. The proportion of species belonging to studied trophic-level groups is unequal within the research plots (Table 3). The share of fungal decomposers inhabiting plant litter, forest floor, and humus is 25–30% in forest ecosystems and woodlands (RP I–III); while in tundra it reaches 78% (RP IV) with the dominance of humus saprobionts (67%). Tundra biotopes are dominated by macromycetes inhabiting forest floor and humus, due to the presence of plant litter and the absence of woody substrate necessary for wood-decaying fungi (so no wood-decaying fungi was found there).
The advantages and disadvantages of metabarcoding (ITS sequencing) of fungi are described in [44,45]. One clear advantage is the ability to identify a larger number of species compared to classical methods for studying fungal communities. However, for high latitudes, the use of this approach is complicated by the lack of sufficiently complete reference databases. A more universal technical problem is the difficulty in recognizing taxa due to either too high or insufficient intraspecific variability in fungi [44].
Nevertheless, it seems reasonable to compare our results with data on fungal diversity obtained using DNA barcoding. It was unexpected that the taxonomic diversity of mycorrhiza-forming basidiomycetes established in a study of Arctic communities involving larch [46] was found to be lower than that obtained in our studies. This work revealed the presence of only nine operational taxonomic units (OTUs) in the communities under study, in contrast to the 37 species we identified (Figure 5). This discrepancy is likely due to the greater diversity of plant communities that were studied in this work compared to that of [46]. A total of 115 amplicon sequence variants (ASVs) belonging to the basidiomycetes were identified in the soils of Spitsbergen, representing 13.07% of the total fungal diversity [47]. This represents a notable increase compared to our findings, although to some extent it can be attributed to the more diverse conditions under which the soil samples were collected for analysis.
A comprehensive examination of the North American Arctic, conducted in tundra communities, led to the identification of 1834 OTUs, of which 486 were classified as basidiomycetes, belonging to 36 families [48]. A comparison of our results with data for zone E, which is the most similar in climatic conditions to Norilsk, showed a high degree of similarity between them in the level of alpha diversity (73 OTUs versus 70 species in our research plots). A detailed analysis of the similarities in the structure of trophic groups is difficult due to the lack of this information in [48]. However, the data we know about the taxa found in the North American Arctic allows us to conclude that there are no sharp differences. It should also be noted that the distribution of the most typical fungal taxa found in [48] is not limited to the Arctic. This conclusion is entirely consistent with the data obtained.
5. Conclusions
The diversity of macromycetes in the studied area is relatively limited at the species level but is comparable to that observed in more southern ecosystems at the genus-to-order level. The leading families in terms of the number of species are Russulaceae (14 species), Polyporaceae (6), Tricholomataceae (5), and Suillaceae (5). This represents the inaugural comprehensive data set on Basidiomycota in the Arctic zone of Central Siberia. The most common species are mycorrhizal fungi (37%), which form a symbiotic association with plant roots, and wood decay fungi (34%), which decompose woody debris. The rest (29%) are fungal decomposers inhabiting plant litter, the forest floor, and humus.
The number of species and their habitat and trophic spectrum depend on the diversity of plant substrates. The largest number of species occur in forest biotopes dominated by mycorrhizal and wood decay fungi (up to 70%), that are trophically associated with woody plants and woody debris. The fungal decomposers inhabiting plant litter, the forest floor, and humus dominate (about 80%) in the species composition of tundra, where, in the absence of woody substrate, wood decay fungi have not been found at all. The ecological structure of the arctic fungal community is similar to that of the taiga and temperate forests. The data obtained can be utilized for the purpose of identifying the biological processes occurring in the Arctic ecosystems.
The study of macromycetes in Northern Siberia revealed their species diversity and role in the unique high-latitude ecosystems. Since fungi form fruit bodies irregularly and their life cycle is short in such harsh conditions, it is impossible to identify all macromycete species, inhabiting northern ecosystems, at once [49]. Thus, further research on macromycetes in the study area is needed.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Utkin A.I. Forest biogeocenoses of the cryogenic region as specific systems Ecology 197631522
- 2Pozdnyakov L.K. Permafrost Forestry L.K. Nauka Novosibirsk, Russia 198396 p
- 3Pozdnyakov L.K. Permafrost Forestry L.K. Nauka Novosibirsk, Russia 1986192 p
- 4Abaimov A.P. Larch Forests and Woodlands of Northern Siberia: Diversity, Ecology and Forest Forming Process Ph.D. Thesis Sukachev Institute of Forests Krasnoyarsk, Novosibirsk, Russia 1997
- 5Abaimov A.P. Bondarev A.I. Semechkin I.V. Ecological and geographical features of tree stand dynamics in Siberia Proceedings of the Forest Dynamics and Its Mechanisms: Proceedings of International Workshop Tsukuba, Japan 6–9 September 19936768
- 6Abaimov A.P. Prokushkin S.G. Zyryanova O.A. Kaverzina L.N. Permafrost larch forests and features of their forming and functioning Lesoved 199751323
- 7Karatygin I.V. Nezdoiminogo E.G. Novozhilov Y.U.K. Zhurbenko M.P. Fungi of the Russian Arctic Marafon St. Petersburg, Russia 1999212 p
- 8Shiryaev A.G. Mukhin V.A. Clavarioid-type fungi of Svalbard: Spatial structure in the European High Arctic N. Am. Fungi 201056784