Impacts of Millipedes on Acari and Collembola Communities—A Microcosm Experiment
Wenjin Chang, Peng Zhang, Jianwei Li, Nonillon M. Aspe, Jiahua Hao, Siyuan Lu, Zhuoma Wan, Donghui Wu

TL;DR
Millipedes affect soil microarthropods by reducing Collembola in soil while increasing them in litter, and boosting Acari in soil, altering community structures.
Contribution
This study reveals millipedes' role as ecosystem engineers affecting soil microarthropod communities through non-trophic interactions.
Findings
Millipede activity reduced Collembola abundance and diversity in soil but increased them in the litter layer.
Acari abundance and diversity increased in soil due to millipede activity.
Millipedes altered community structures and strengthened correlations between Acari and Collembola.
Abstract
Interactions among soil organisms are crucial for food webs and ecological functions. However, the effects of millipedes, which play a key role as decomposers in the soil, on soil microarthropods remain unclear. This study demonstrates that millipede activity in the soil decreased the abundance and diversity of Collembola in the soil while increasing their abundance and diversity in the litter layer. Consequently, this led to an increase in the abundance and diversity of Acari in the soil. The dominant species of both Acari and Collembola were most notably affected by the experiment. Millipede activity significantly altered the community structure of Acari and Collembola and strengthened the correlation between these two groups. These findings enhance our understanding of the ecological role of millipedes. Ecosystem engineers influence the structure and function of soil food webs…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
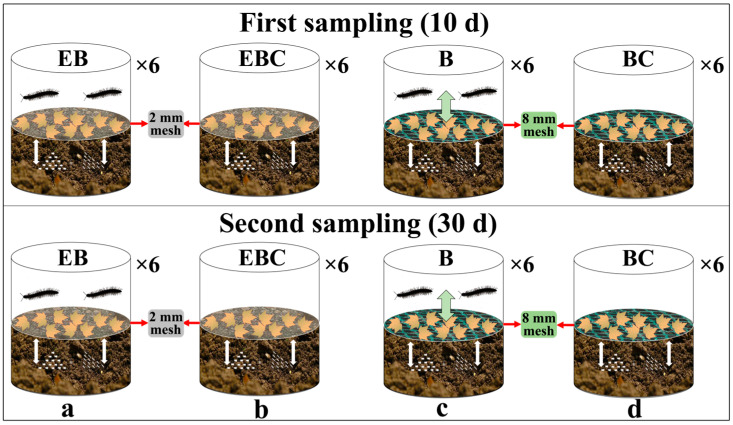 Figure 1
Figure 1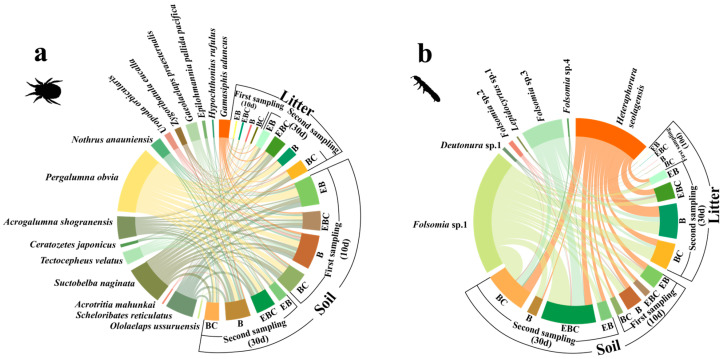 Figure 2
Figure 2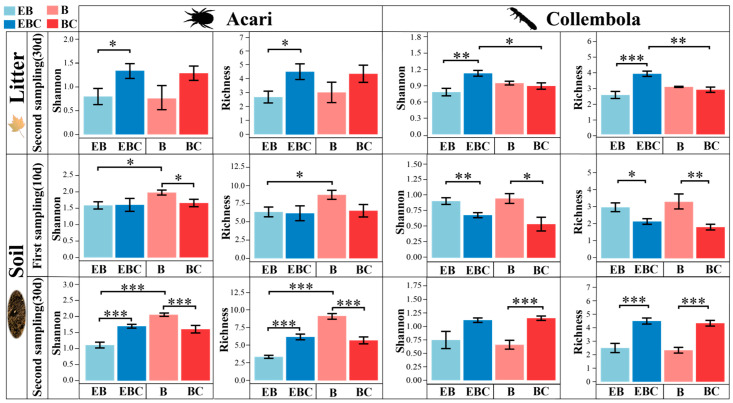 Figure 3
Figure 3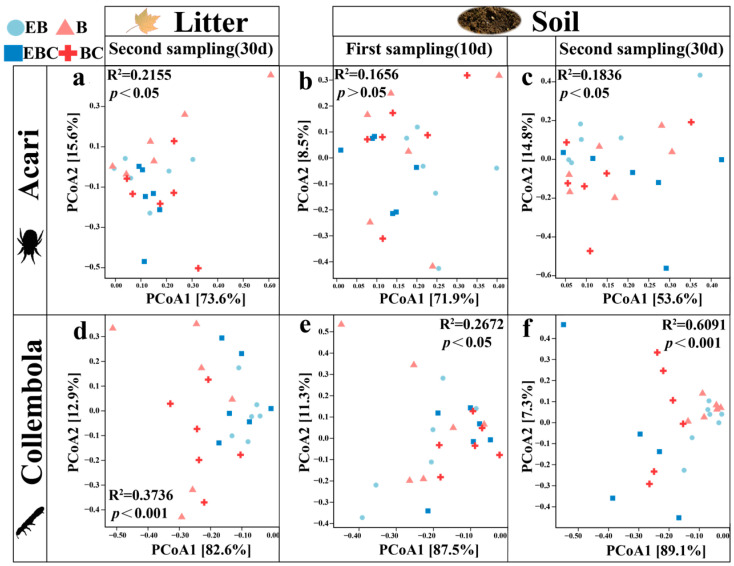 Figure 4
Figure 4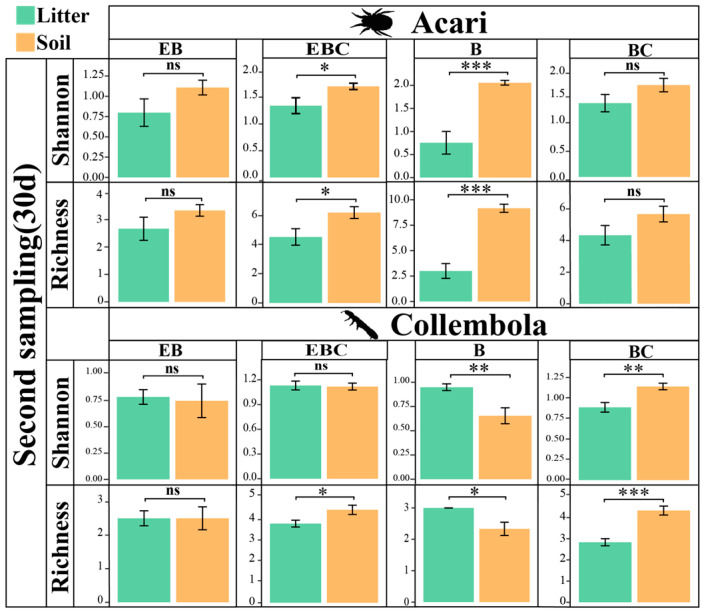 Figure 5
Figure 5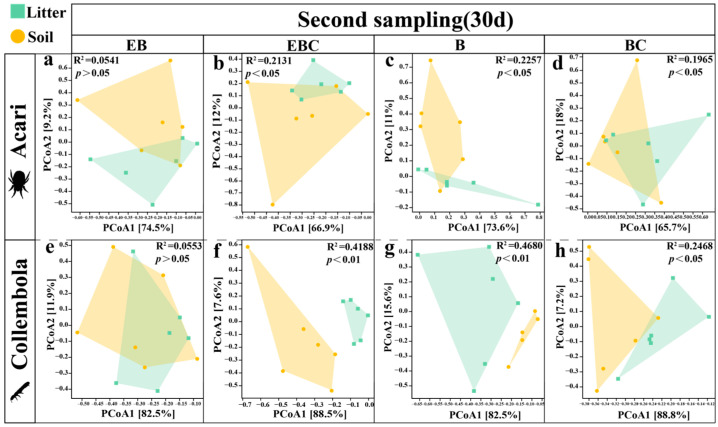 Figure 6
Figure 6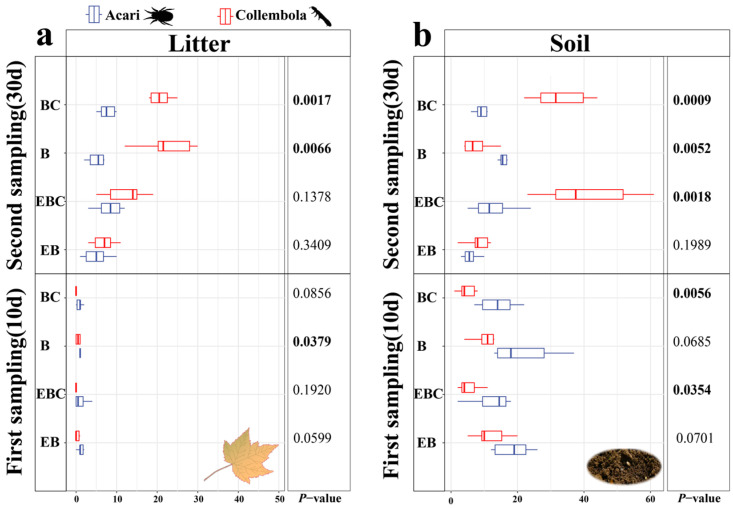 Figure 7
Figure 7- —National Natural Sciences Foundation of China
- —Program of Introducing Talents of Discipline to Universities
- —“One belt and one road” international scholarship from Ministry of Science and Technology of China
- —President’s International Fellowship Initiative from the Chinese Academy of Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStudy of Mite Species · Collembola Taxonomy and Ecology Studies · Fire effects on ecosystems
1. Introduction
Interactions between organisms are major determinants of biodiversity, species distribution, and abundance [1,2,3], profoundly affecting ecosystem stability [4]. Soil is among the most species-rich habitats on Earth [5], and the animals living within it play a crucial role in nutrient cycling, material utilization, and assessing soil health [6,7]. Most current studies focus on aboveground–belowground interactions, animal–plant relationships, and microbial interactions [8,9,10]. However, the mechanisms of interactions among soil organisms and their impacts on ecosystem processes are still not well understood. Soil invertebrates play crucial roles in key ecological interactions [11], and their function as ecosystem engineers significantly enhances species richness [12].
Forty-five percent (45%) of interactions in the soil are generated by burrowing ecosystem engineers [8]. Larger invertebrates influence the diversity of smaller organisms by facilitating their dispersal and modifying soil habitats [13,14]. Millipedes, considered important soil ecosystem engineers, are abundant, diverse, and widely distributed [11,15,16]. They impact soil fauna habitats to varying degrees [17] and enhance soil aeration by creating a more porous soil structure through the accumulation of excavated soil or manure [18,19,20]. Some species have been shown to have significant effects on microbial biomass, enzymatic activity, and soil aggregates [21,22,23,24]. Their feces has been used as an indicator of organic matter turnover, altering organic matter and C and N stabilization [25,26]. Compared to earthworms, millipedes have a greater ability to break down litter and alter the state of the litter, thereby altering food resources [18].
Acari and Collembola, key constituents of soil microarthropods, are among the most abundant and diverse soil fauna [27,28]. Serving as indicator organisms of soil quality [29,30], they occupy various soil and litter layers [31]. These organisms rely on soil pore space and food resources such as litter and microorganisms for survival [32,33,34,35]. Acari and Collembola constitute a significant portion of decomposer communities, with their dominant species being particularly sensitive to environmental changes [36,37,38]. Larger ecosystem engineers, like earthworms, affect Acari and Collembola differently by altering soil structure and food resources, thereby changing community composition and relative abundance [32,39,40].
Research on the interactions between millipedes and other organisms has primarily focused on their relationships with other soil macrofauna, such as earthworms, and how these associations influence litter decomposition, alter the soil environment, and regulate microorganisms [41,42]. Currently, there is very limited information on the effects of millipedes on soil microarthropod communities. Therefore, further experiments are needed to explore the role of millipedes on microarthropods [43]. In this work, we investigated the effects of millipedes on Acari and Collembola communities in the soil. Our hypotheses revolve around three key points: (a) millipedes differentially affect the structure and composition of Acari and Collembola communities while altering their distribution in soil and litter; (b) the presence and activity of millipedes contribute to alterations in the relationship between Acari and Collembola; and (c) the most pronounced changes within the Acari and Collembola communities will be observed through their dominant species.
2. Materials and Methods
2.1. Study Area
The materials used in the experiments were obtained from the Changchun Agricultural Positioning Experiment Station, Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences (44°59′ N, 125°23′ E), in Changchun City, Jilin Province, China. The area has a temperate continental climate with an average annual temperature of 4.6 °C–6.4 °C and average annual rainfall of 614 mm. The type of soil is black (Typic Hapludoll, USDA, 1993) with clay loam texture. Orthomorphella pekuensi (Karsch, 1881) is a millipede widely distributed in China [44] and is the dominant large soil animal in the study site, measuring about 20 to 35 mm in length. Like most millipedes, O. pekuensi is a scavenging species that feeds primarily on plant litter or organic residues in various stages of decomposition. The litter of Populus tomentosa Carr was selected as the study material as it is the dominant tree species in the study site. It is also one of the main food sources for millipedes and soil microarthropods. The experiment was conducted in mid-August, when the study site was in the highest number and it was the most active time for the millipedes.
2.2. Microcosmic Experimental Design
The experiment utilized four microcosmic treatments as follows: (a) Millipedes were introduced to the litter layer, with a 2 mm mesh barrier placed between the litter layer and the soil layer to prevent millipedes from burrowing into the soil while allowing the activities of Acari and Collembola. This treatment is abbreviated as exclusion burrowing (EB). (b) A 2 mm mesh barrier was added in the same manner as (a), but without millipedes, serving as a control for exclusion burrowing (EBC). (c) Millipedes were added to the litter layer, and to avoid interference from the mesh, an 8 mm mesh was added to both the litter and soil layers, allowing millipedes to burrow into the soil. This treatment is abbreviated as burrowing (B). (d) An 8 mm mesh was added without millipedes, serving as a control for burrowing (BC) (see Figure 1). The 2 mm mesh permitted the movement of Acari and Collembola between the soil and litter layers while preventing millipedes from passing through the mesh [45]. Conversely, the 8 mm mesh did not impede the movement of millipedes along with Acari and Collembola. The experiment spanned a duration of 30 days, during which, two destructive sampling sessions were conducted on days 10 and 30, taking into account the impact of millipedes on feces production and soil structure alteration within this timeframe. Each microcosm treatment was replicated six times, resulting in a total of 48 microcosms for the experiment. Each microcosm comprised a cylindrical PVC plastic cup measuring 130 mm in height, with a bottom diameter of 83 mm and a mouth diameter of 95 mm. Small 0.1 mm filters were fitted onto the lids of the plastic cups to prevent soil animals from escaping while maintaining air permeability (see Figure 1).
2.3. Experimental Material Collection and Pre-Processing
2.3.1. Millipede Collection and Pre-Processing
All millipedes used in the experiment were collected manually from the Positioning Experimental Station. Samples were taken two days after the rain, when the soil had the optimum moisture content (between 17% and 28%). Millipedes and soil microarthropods were more active at this time. The millipedes were incubated in four boxes before being used for the experiment. The rearing conditions were as follows: temperature of 20 °C, photoperiod 12 h/12 h, and air humidity of 40% (referring to the same conditions as the sampling site). The microcosms were misted with water on a weekly basis to keep the millipedes in a moist environment. The millipedes were cultured for one month using soil (0–10 cm) and aboveground litter (moderately broken) from the collection site as the medium to acclimatize them to the laboratory culture environment. The larger adult millipedes were selected from the pre-cultured millipedes prior to the start of the experiment (body width greater than 2 mm and less than 8 mm). These millipedes were starved for 48 h to void their intestine.
2.3.2. Soil Collection and Pre-Processing
A square sample plot measuring 20 × 20 m was designated at the millipede collection site. Within this plot, five points were selected: one at the center and four at the corners, each measuring 1 × 1 m. From these points, soil samples from the 0–5 cm depth were collected. The soil obtained from the five points was combined and manually sorted to remove larger roots, stones, and plant debris, as well as soil macrofauna visible to the naked eye [46]. These unfiltered soils retained their natural microarthropod communities. Prior to being introduced into the experiment, the soil was homogenized to ensure consistency in the Acari and Collembola communities and population numbers across each microcosm [47].
2.3.3. Litter Collection and Pre-Processing
Litter at the site was collected at the same time soil samples were collected. Intact leaves were chosen and then cut into pieces approximately 0.8–2 cm^2^ in size [41]. To remove microarthropods and insect eggs from the litter while preserving the microbial community as effectively as possible, the leaves were dried in an oven at 70 °C for 24 h [48].
2.3.4. Assembly of Microcosmic Experimental Setup
An average weight of 160 g of moist soil with its natural microarthropod community was introduced into each microcosm. The initial moisture content of the soil in each experimental unit was measured. Distilled water was added to reach 20% moisture content. The dried leaves were mixed and weighed, and 2 g of litter was added to each microcosm. Meanwhile, 2 mm and 8 mm mesh were inserted, respectively, between the litter and the soil to serve as barriers for the movement of soil fauna. The leaves were lightly sprayed with water to maintain moisture. Treatment microcosms were placed in an incubator at 20 °C for 48 h until the leaves were completely moist [43,49] (Figure 1).
2.4. Cultivation and Sample Collection for Microcosm Experiments
Once the microcosm was fully assembled, millipedes of similar weight and length were introduced into the litter layer of the microcosm after they were starved. Two millipedes were added to each microcosm (consistent with the density of millipedes at the collection site). The survival rate of millipedes was assessed daily. The millipedes were kept in a moist environment by lightly spraying water every 2–3 days. If a millipede died before the end of the experiment, it was removed and replaced with a millipede of similar size and weight. The microcosm was maintained in an incubator subjected to constant temperature and soil moisture (20 °C and 20%, respectively) for 30 days. The activity and consumption of food resources by millipedes within this period was sufficient enough to have an impact on the soil [43].
The litter in each microcosm was carefully pinched out with tweezers, and then, all the soil in each microcosm was poured out. The litter and soil were placed on separate labeled sieves using the modified Tullgren funnel method. After 10 days, samples were collected and stored in 95% ethanol at −20 °C and were prepared for identification (with the first sampling event randomly selecting 24 microcosms). All Acari and collembolans were separated and counted using a stereomicroscope. The adults were identified at the species level [50,51,52,53].
2.5. Statistical Analysis
The data were processed and analyzed using R 4.1.1. Analysis of variance (ANOVA) was used to compare the abundance of Acari and Collembola and community alpha diversity between different treatments in the litter and soil layers. If the data did not follow a normal distribution, the Kruskal–Wallis test was used. In order to compare the abundance and community alpha diversity of Acari and Collembola between litter layer and soil layer under the same treatment, a T-test was used, and a non-parametric test was used to test the data with great deviation. Principal Coordinate Analysis (PCoA) was utilized to elucidate the distribution patterns of Acari and Collembola under different treatments. Furthermore, differences in community composition under different treatments were examined through the permutational multivariate analysis of variance (PERMANOVA). These analyses were conducted using the “vegan” package [54]. In order to compare the relationship between different species of Acari and Collembola, correlation heatmaps were constructed using Pearson correlation coefficients. The creation of these heatmaps relied on the implementation of the “pheatmap” package [55]. The classification of dominant, common, and rare taxa was based on the relative abundance table of Acari and Collembola, utilizing the following criteria: taxa with an individual count exceeding 10% of the total number were classified as dominant, taxa with a count between 1% and 10% were classified as common, and taxa below 1% were classified as rare [56]. Significance levels were established as follows: correlations and differences were deemed significant at p < 0.05, particularly significant at p < 0.01, and highly significant at p < 0.001.
3. Results
3.1. Abundance and Composition of Acari and Collembola Communities
In this experiment, a total of 2195 Acari and Collembola individuals were collected. Among these, there were 15 species of Acari from 15 genera and 13 families and 7 species of Collembola from 4 genera and 4 families. In the initial sampling event, no significant impact on the abundance of Acari and Collembola in the litter was observed in the EBC and BC treatments. However, the presence of millipedes (EB and B) significantly increased the abundance of Acari (p < 0.001) and Collembola (p < 0.01) in the soil (refer to Figure 2 and Table S1). In the subsequent sampling event, a notable increase was observed in the abundance of Acari in the soil in treatment B (p < 0.05), while there was a decrease in the abundance of both Acari and Collembola in the EB treatment (p < 0.001) (Figure 2, Table S2).
The same species were clearly growing more in the soil than in the litter at the time of the first sampling event (Figure 2, Table S3). At the time of the second sampling event, there was a significant increase in Collembola abundance in the litter of the B treatment (p < 0.001), while there was a significant decrease in Collembola abundance in the litter layer vs. the soil layer in the BE treatment (p < 0.001) (Figure 2, Table S4). There was significantly higher Acari and Collembola abundance in the soil than in the litter in the control groups (EBC and BC) (p < 0.05) (Table S4).
The dominant Acari taxa in the soil at the time of the first sampling event were Scheloribates reticulatus of Scheloribatidae, Suctobelba naginata of Suctobelbidae, and Pergalumna obvia of Galumnidae, which accounted for 49.33–71.17% of the total Acari species. Heteraphorura seolagensis and Folsomia sp.1 accounted for 88.73–100% of the overall Collembola population. The addition of millipedes increased the common taxa of Collembola with rare taxa of Acari (Figures S1 and S2). The dominant Acari taxa in the second sampling event of litter and soil were Scheloribates reticulatus, Suctobelba naginata, and Pergalumna obvia and Acrogalumna shogranensis of Galumnidae, which accounted for 52.12–73.34% of the total Acari species. Meanwhile, Heteraphorura seolagensis, Folsomia sp.1, and Folsomia sp.3 accounted for 86.49%–100% of the total Collembola species. The addition of millipedes reduced the dominant taxa of Acari in litter and soil (Figures S1 and S2).
3.2. Changes in Acari and Collembola Communities in Litter and Soil under Different Treatments
The number of Acari and Collembola collected in litter layer during the first sampling event was very small. At the time of the first sampling event of the soil, the diversity of Collembola increased in the EB and B treatments (p < 0.01), while the diversity of Acari significantly increased in the B treatment (p < 0.05) (Figure 3). In the second sampling event, the diversity of Acari and Collembola in the litter and soil significantly decreased in the EB treatment (p < 0.001 and p < 0.001, respectively). Compared with EB treatment, B treatment increased the diversity of Acari in soil (p < 0.001) and decreased the diversity of Collembola in soil (p < 0.001) but had no significant effect on Acari and Collembola in litter (Figure 3).
The number of Acari and Collembola colonizing the litter for the first time was small and insufficient to analyze the community structure. However, the change in the community structure of Collembola in the soil was more pronounced (R^2^ = 0.2672; p < 0.05) (Figure 4e). In the second sampling event, the presence of millipedes (EB and B) caused a significant change in the Acari and Collembola communities and a more pronounced change in the Collembola community structure (R^2^ = 0.6091; p < 0.001) (Figure 4). The EB treatments had a greater effect on Acari in the soil, and the B treatments had a greater effect on the Collembola community structure in the soil (Figure 4).
At the time of the second sampling event, the diversity of Acari in the soil and Collembola in the litter significantly increased in the B treatment (p < 0.001 and p < 0.01, respectively). Exclusion burrowing (EB) significantly reduced the diversity of Acari and Collembola in the litter (p < 0.05) (Figure 5).
Comparing the litter and soil layers, the community structure of Acari and Collembola was significantly different in the control groups (EBC and BC) and burrowing (B) treatments (p < 0.05) (Figure 6). The EB treatments did not have a significant effect on Acari and Collembola community structures (Figure 6a,e).
3.3. Relationship between Acari and Collembola
In the first sampling event, the dominant Acari and Collembola species in the litter and soil showed a significant positive correlation (p < 0.05) (Figure S3), which were made more pronounced with the addition of millipedes (EB and B) (p < 0.001) (Figure S3). In the second sampling event, burrowing (B) intensified the positive correlation (p < 0.05) (Figure S3).
With regard to the overall abundance of Acari and Collembola, significant changes were observed in the control groups (EBC and BC) (Figure 7). Acari abundance was significantly higher than Collembola abundance in the soil in the B treatment in the second sampling event (p < 0.01) (Figure 7b), while Collembola abundance was significantly higher than Acari in the litter layer (p < 0.01) (Figure 7a).
4. Discussion
4.1. Effect of Mechanical Disturbance of Millipedes on Acari and Collembola
The presence and burrowing activities of millipedes had a strong influence on Acari and Collembola in the soil and litter (Figures S1 and S2; and Tables S3 and S4), supporting Hypothesis a. Millipedes burrowed in the soil (B), creating numerous visible pores and a softer texture at the end of the experiment, while the large number of different shapes and sizes of feces produced by millipedes changed the soil structure (Figure S5a,b). Millipedes affect small soil animals and microorganisms by altering the physical environment and creating microhabitats [14]. Soil quality is heavily influenced by the activity of soil macrofauna. Similar to other ecosystem engineers like earthworms and ants, millipedes enhance soil pore space by their burrowing activities, consequently improving soil permeability [20,57]. Various species of Collembola are adapted to different soil depths and microhabitat conditions [58]. Studies have suggested that the increase in Folsomia spp. populations may be linked to changes in soil porosity induced by earthworms [59,60]. Oribatid mites, a type of Acari, primarily feed on detritus and fungi, typically inhabiting organic and litter layers [61]. Although millipedes mostly dwell near the soil surface or beneath litter, their digging behavior, akin to earthworms, can transport more food resources deeper into the soil [62]. This behavior facilitates soil-dwelling microarthropods’ access to food, aiding in their survival. When facing millipedes in the soil, similar to the effects of earthworms on soil structure, Collembola find it easier to access the soil and evade predators [63]. Collembola, being more mobile than Acari, tend to survive better in the litter layer when predatory Acari populations increase in the soil. Additionally, frequent mechanical disturbance of the soil diminishes the density and abundance of Collembola [64]. This dynamic results in a higher diversity and abundance of Acari in the soil compared to the litter layer (refer to Figure 2 and Figure 4; and Tables S3 and S4). It has been proposed that Acari may exhibit a stronger symbiotic relationship with millipedes [15]. However, different millipede species, with varying body sizes and behaviors, may exert differing impacts on soil microarthropods, necessitating further research on the ecological implications of different soil fauna.
4.2. The Relationship between Acari and Collembola
The presence and activity of millipedes changed the relationship between Acari and Collembola (Hypothesis b) and had a positive effect on the population of some species of both groups (Figures S3 and S4). Acari abundance decreased and community structure was disturbed during the second sampling event, but the diversity was not negatively affected. Collembola reduced in abundance and diversity when confronted with millipedes (Figure 2 and Figure 3 and Tables S1 and S2). Mutualistic symbiosis may be the most variable species relationship among multiple interactions relative to others [65]. Positive effects at the time of the first sampling may be due to the formation of microhabitats, which provide more space and resources for their survival. Reciprocal interactions, as opposed to competitive interactions, depend largely on abiotic factors, and they become more important under extreme conditions according to the stress gradient hypothesis (SGH) [66]. An example is the instability of microbial communities in response to environmental stresses [67]. For Acari and Collembola to respond differently, it is possible that the stress due to environmental change leads to differences between communities. Oribatid mites have long been recognized as k-selective organisms with slow metabolisms, low fecundity, and little ability to respond quickly to environmental changes [68,69]. Consequently, their populations decline when their favorable habitat is disturbed. The low reproductive rates of most oribatid species may be slow to recover from these disturbances [70]. In contrast, several predatory mites, such as the Mesostigmata, were essentially unchanged in the experiments (Tables S1 and S2), probably because they are usually predators of other microarthropods, larvae, and nematodes [71,72]. In contrast to oribatids, mite-predatory microarthropods in a mesocosm exert predatory pressure on soil microarthropods, while their resistance to environmental change may be high due to a wider range of feeding options [73,74]. Among the soil fauna, Collembola are considered as a rapid response group (r-strategist) to environmental changes. They are more prolific and react more quickly in the face of disturbances [64,75]. Therefore, there was a sharp increase in the number of Collembola species during the second sampling event, and this greater density reduced the space available for Acari, leading to a decrease in their abundance. When the Collembola community showed a decreasing trend, the competition for food resources may have been reduced, so the abundance and diversity of the Acari community increased significantly.
4.3. Main Mechanisms Driving the Relationship of Millipedes with Acari and Collembola
Overall, the presence of millipedes in this experiment had a significant impact on the abundance, diversity, and community structure of Acari and Collembola (Figure 2, Figure 3 and Figure 4, and Tables S1 and S2). The presence of soil macrofauna has a strong influence on the soil microarthropods by altering the soil structure, food resources, and their interactions such as predation and competition [40,76]. Millipedes not only provide litter resources directly to soil microorganisms, but also alter the physical state of the litter and soil [21]. Millipedes differ from earthworms in that they have more developed mouthparts, which break up the litter to produce finer debris during the feeding process, increasing the range of food resources available to the microarthropods by providing easier access to them [77]. The genus Folsomia has a small percentage of fungi in its gut [34], and fungi are not a major food resource for this Collembola genus [78]. Members of this genus are thought to feed on litter and detritus [79,80]. The distribution of Acari is somewhat related to resource constraints [33]. Some Acari of the genus Scheloribates are categorized as pan-phytophagous mites, which have numerous distributions in grassland soils [81], and plant litter is an important food source for this species [82]. Certain Acari species, such as Tectocepheus, exhibit broad dietary preferences, consuming various resources from lichens to fungi [83]. However, there were no notable changes in abundance observed across the treatments (Tables S1 and S2). Pergalumna spp. of the Galumnidae family, under the isotope test, were shown to feed on a wide range of resources, including live nematodes and collembolans [84]. They have a wider range of food choices and have been observed to be more stable to millipede disturbance, and thus, were not significantly affected across the treatments.
It was observed in the experiment that a large amount of millipede feces accumulated on the soil surface or in the litter layer (Figure S5). The persistence of millipede feces resulted in a longer accumulation of soil carbon [21]. Their crushing of the litter for partial digestion and repackaging into feces increased the instability of the organic matter [25]. The presence of this feces changes the composition of the microbial community in the soil [42,43], which in turn changes the allocation of its food resources, leading to changes in the Acari and Collembola communities. Acari and Collembola have a wide range of dietary habits [34,85], and the presence of leaf litter and the abundance of fungi provide abundant food resources for Acari and Collembola that feed on detritus and fungi [86]. The mite species Acrogalumna shogranensis, which was dominant throughout the experiments, had a strong susceptibility to millipede activity (Figure 2; and Tables S3 and S4). Most Acrogalumna species feed on fungi [87]. Changes in the dominant species of the Acari and Collembola communities were most pronounced when millipedes were present in the different treatments (Figure 2 and Figures S1, S3 and S4). The abovementioned changes in dominant species also proved to support Hypothesis c. As Acari and Collembola are the two dominant microarthropod groups, their abundance and diversity drive the overall microarthropod community [86]. The dominant species of these groups play an important role in maintaining biodiversity in the soil and in the stabilization of the ecosystem through the decomposition and recycling of materials, and the presence of millipedes may increase food resources and survival space for the microarthropod groups.
5. Conclusions
This study showed that millipedes had different effects on different species of Acari and Collembola. In the short term, millipedes significantly increased the abundance and diversity of soil Acari and Collembola, but the rapid growth of Collembola over time suppressed the Acari community. Millipede burrowing predisposes collembolans to colonize the litter layer, resulting in an increase in soil Acari abundance and diversity. Millipedes altered the structure and composition of the Acari and Collembola communities and intervened in their interactions. These changes were driven by the dominant species in the Acari and Collembola communities. These results provide support for the effects of soil macrofauna on soil microarthropod communities to better understand the interactions between soil organisms and the effects on soil biodiversity. Further studies are needed to investigate the main drivers of the relationship between millipedes and soil microarthropods.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wilson W.G. Lundberg P. Vázquez D.P. Shurin J.B. Smith M.D. Langford W. Gross K.L. Mittelbach G.G. Biodiversity and species interactions: Extending Lotka–Volterra community theory Ecol. Lett.2003694495210.1046/j.1461-0248.2003.00521.x · doi ↗
- 2Ricklefs R.E. A comprehensive framework for global patterns in biodiversity Ecol. Lett.2004711510.1046/j.1461-0248.2003.00554.x · doi ↗
- 3Brophy C. DooleyÁ. Kirwan L. Finn J.A. Mc Donnell J. Bell T. Cadotte M.W. Connolly J. Biodiversity and ecosystem function: Making sense of numerous species interactions in multi-species communities Ecology 2017981771177810.1002/ecy.187228444961 · doi ↗ · pubmed ↗
- 4Gorter F.A. Manhart M. Ackermann M. Understanding the evolution of interspecies interactions in microbial communities Philos. T. R. Soc. B.20203752019025610.1098/rstb.2019.0256 PMC 713353832200743 · doi ↗ · pubmed ↗
- 5Bardgett R.D. van der Putten W.H. Belowground biodiversity and ecosystem functioning Nature 201451550551110.1038/nature 1385525428498 · doi ↗ · pubmed ↗
- 6Swift M.J. Andrén O. Brussaard L. Briones M. Couteaux M.-M. Ekschmitt K. Kjoller A. Loiseau P. Smith P. Global change, soil biodiversity, and nitrogen cycling in terrestrial ecosystems: Three case studies Global Chang. Biol.1998472974310.1046/j.1365-2486.1998.00207.x · doi ↗
- 7Bagyaraj D.J. Nethravathi C.J. Nitin K.S. Soil Biodiversity and Arthropods: Role in Soil Fertility Economic and Ecological Significance of Arthropods in Diversified Ecosystems Springer Singapore 20161751
- 8Wurst S. Sonnemann I. Zaller J.G. Soil macro-invertebrates-their impact on plants and associated aboveground communities in temperate regions Aboveground-Belowground Community Ecology Ohgushi T. Wurst S. Johnson S.N. Springer Tokyo, Japan 2018 Volume 234175200