Promising New Methods Based on the SOD Enzyme and SAUR36 Gene to Screen for Canola Materials with Heavy Metal Resistance
Yue Dai, Hao Chen, Yufang Li, Rongkui Hui, Zhenqian Zhang

TL;DR
This study identifies SOD enzyme and SAUR36 gene as potential tools to screen canola plants resistant to heavy metal stress, improving breeding efforts.
Contribution
The study introduces SOD and SAUR36 as novel indicators for screening canola materials with heavy metal resistance.
Findings
F335 showed higher SOD and POD activities than F338 under heavy metal stress.
SAUR36 and SAUR32 were identified as key differential genes in heavy metal tolerance.
SAUR36 expression was significantly higher in field-grown canola, with FY958 showing the largest increase.
Abstract
Canola is the largest self-produced vegetable oil source in China; however, excessive levels of cadmium, lead, and arsenic seriously affect its yield. In this study, canola near-isogenic lines with different oil contents (F338 (40.62%) and F335 (46.68%) as the control) were used as the experimental materials, and heavy metal stress experiments and omics analysis were carried out. The results show that superoxide dismutase and SAUR36 were closely related to heavy metal stress tolerance. Therefore, they may be used to screen for new canola materials with good heavy metal stress tolerance for canola breeding. Canola is the largest self-produced vegetable oil source in China, although excessive levels of cadmium, lead, and arsenic seriously affect its yield. Therefore, developing methods to identify canola materials with good heavy metal tolerance is a hot topic for canola breeding. In…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1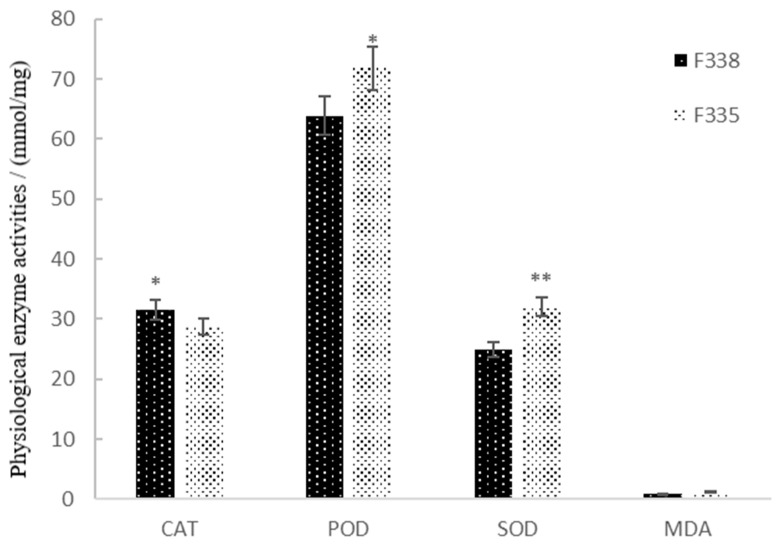 Figure 2
Figure 2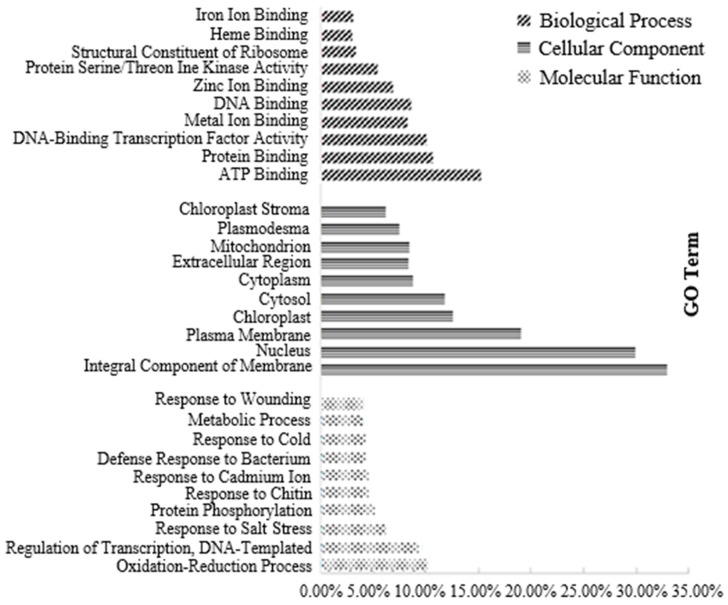 Figure 3
Figure 3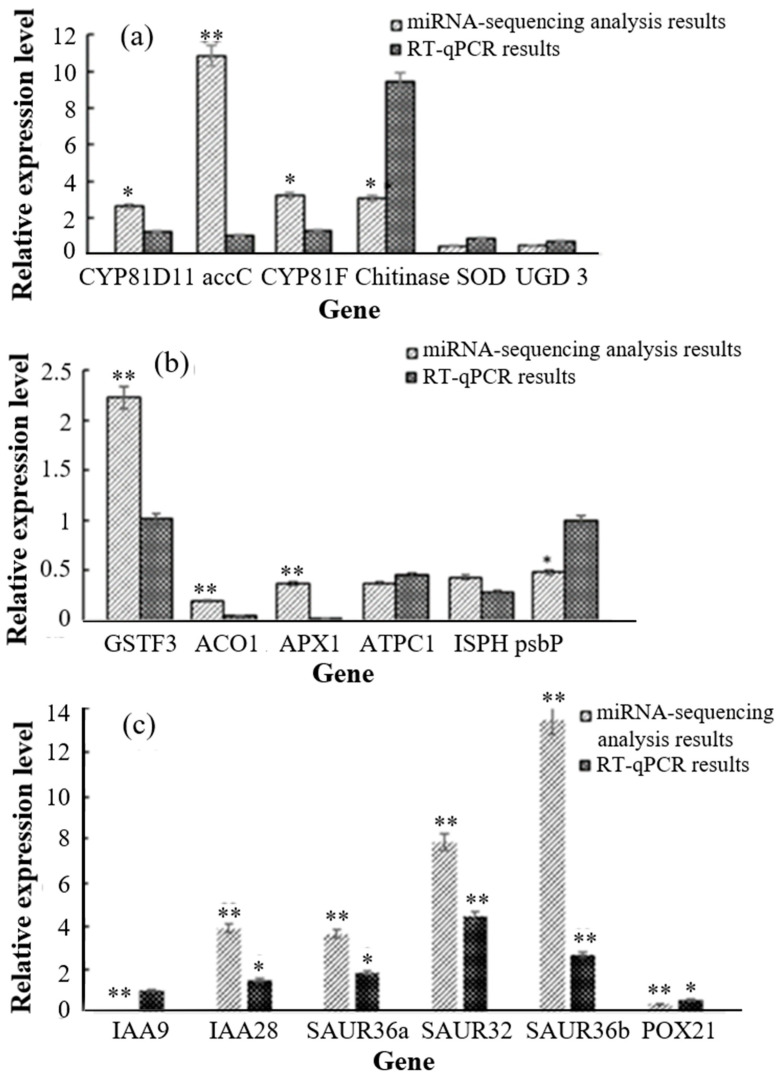 Figure 4
Figure 4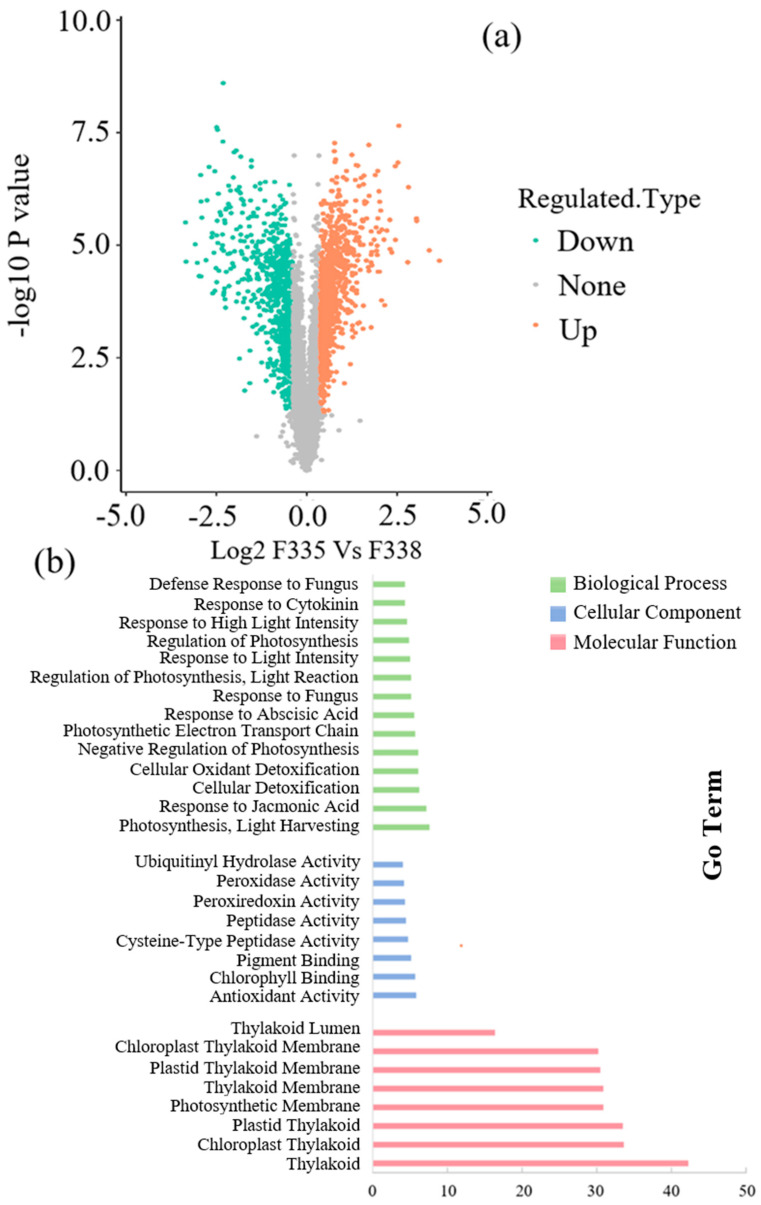 Figure 5
Figure 5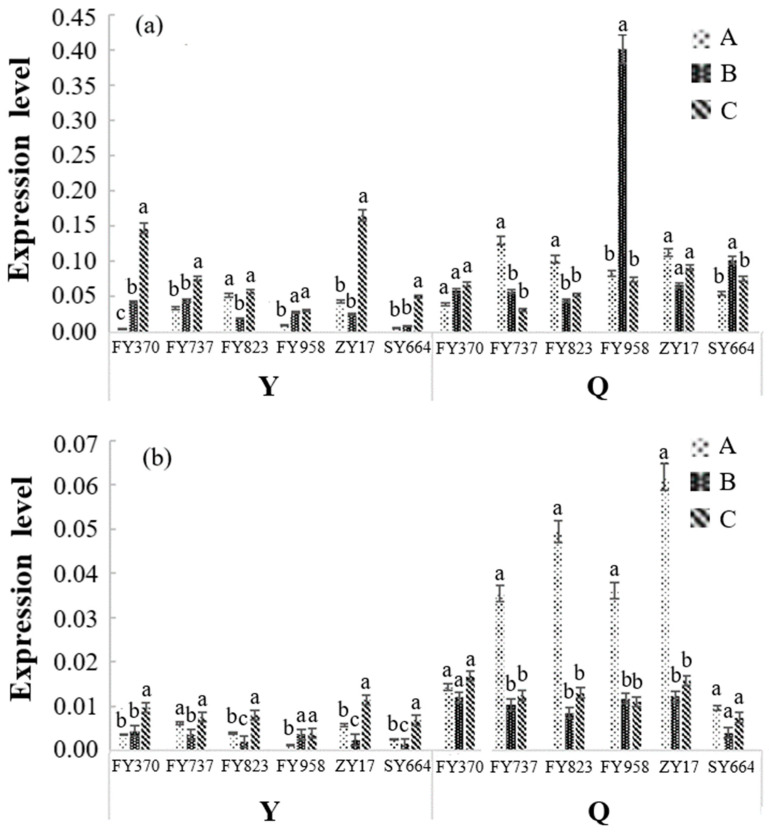 Figure 6
Figure 6- —National Science and Technology Innovation 2030 Major Projects
- —National Key Research and Development Plan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTannin, Tannase and Anticancer Activities · Chromium effects and bioremediation · Genomics, phytochemicals, and oxidative stress
1. Introduction
In China, edible oil is in short supply [1,2], and canola is the largest source of edible vegetable oil. Thus, it is important to increase the planting area and yield of canola [3]. However, the point overshooting rate of Chinese arable land has reached 19.4% [4]. As the largest canola planting area in China, Hunan Province is deeply affected by excessive heavy metals [5], among which the levels of Cd, Pb, Hg, and As were measured as 7.0%, 1.5%, 1.6%, and 2.7%, respectively. Rice is difficult to sell due to excessive cadmium levels.
Heavy metals in soil lead to the production of secondary metabolites in plants and serious phytotoxicity [6], which seriously affects the growth, metabolism, physiology, and aging of plants [7]. Cd accumulation affects plants’ absorption of mineral elements [8], alters photosynthesis [9] and antioxidant enzyme activities [10], and even leads to death [11]. Pb affects the transport of plant materials [12] and negatively affects metabolic processes [13], causing delays in growth and germination [14]. As inhibits plant root growth and causes plant death [15]. Poor arable land quality significantly impacts the growth and development of canola [16], resulting in a poor canola yield. Therefore, screening materials with good heavy metal tolerance and studying their internal molecular mechanisms is a key topic of canola breeding research at present [17].
Heavy metals in soil may be easily absorbed by plant roots and transported to other organs and tissues, followed by oxidative stress and the production of related proteins and hormones, such as amino acids, antioxidants, and signaling molecules; compounds such as glutathione, plant chelate peptides, and metallothionein; and enzymes such as superoxide dismutase and peroxide [18]. Plants under metal stress are stimulated by antioxidant enzymes and related metabolic proteins, which play a vital role in signal transduction pathways [19]. Increases in ROS (reactive oxygen species) are considered to be the main phytotoxic effects of heavy metal stress [20,21]. With the development of molecular research technology, sequencing technologies, such as transcriptomics, metabolomics, proteomics, and genomics, and the combination of multiple analysis methods have been widely used [22,23]. For example, transcriptomics and genomics association analyses under different Cd tolerance levels revealed that the Nramp family genes were related to the transport of heavy metal ions in Arabidopsis thaliana, of which BnNramp2; 1 and BnNramp4; 2 were related to Cd transport [23]. Most of the previous studies were limited to a single molecular or physiological level [21,24]. However, the molecular mechanisms of canola’s heavy metal stress tolerance may be related to genes, proteins, and enzymes [25,26]; thus, there is an urgent need for comprehensive research on different aspects, such as the genome, the proteome, physiological characteristics, and field phenotypes [27,28,29].
In this study, canola near-isogenic lines (NILs) with significant phenotypic differences under heavy metal stress were used as the experimental materials, and transcriptomics and proteomics association analyses, verified using RT-qPCR, were used to examine the agronomic traits to identify the key genes or enzymes related to heavy metal stress tolerance and provide a reference for canola breeding.
2. Materials and Methods
2.1. Plant Materials
The canola NILs F335 and F338 were used as the raw materials and were provided by the College of Agriculture, Hunan Agricultural University. The materials had stable conventional propagation characteristics (Table S1).
2.2. Experimental Method
2.2.1. Treatment Methods for Heavy Metal Stress
A mixture of three heavy metal standard solutions of Cd, Pb, and As was prepared for the heavy metal stress experiment, and the water culture germination method was adopted. The concentration of heavy metals was set as 10 times, 50 times, and 100 times the safe standard values of cadmium, lead, and arsenic in water (5 µg/kg, 20 µg/kg, and 10 µg/kg), referred to as 10×, 50×, and 100× in this study (Table 1). The canola in the indoor experiment was placed in a germination box for heavy metal stress growth. Fifty full and disease-free seeds were selected, soaked in 75% alcohol for 30 s for disinfection and washed with sterile deionized water, then soaked in the mixed solution of heavy metals (As, Cd, and Pb) for 12 h and placed in the germination box, and supplemented with heavy metal solution with the same concentration every day for 7 days. The experimental design is 16 h a day of illumination, 8 h of darkness, a temperature of 25 °C, and an illumination intensity of 2455 lux. The experimental method referred to Soares et al. [30] and Kania et al. [31].
For indoor experimental materials, we mainly recorded germination potential, germination rate, emergence rate, and biomass of seeds on the seventh day after germination.
The formula for germination potential is: germination potential (%) = number of seeds germinated on the 3rd day/total number of seeds × 100%.
The formula for germination rate is: germination rate (%) = number of seeds germinated on the 7th day/total number of seeds × 100%
The formula for emergence rate (%) = number of seeds emerged on the 7th day/total number of seeds × 100%.
2.2.2. Physiological Indexes under Heavy Metal Stress
The seedlings on the 7th day of the 100× experiment were taken as samples to detect four enzymes, namely, superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and malondialdehyde (MDA). The SOD, POD, and CAT activities were determined according to Yang et al. [32] and Shi et al. [33], and the MDA content according to Draper et al. [34].
2.2.3. Omics Analysis
The seedlings on the 7th day of the 100× experiment were selected as samples, washed with pure water, frozen in liquid nitrogen, stored at −80 °C, and then sent to BGI Gene and Hangzhou Jingjie Biotechnology Co., Ltd. (Hangzhou, China) for transcriptome and iTRAQ analysis.
A total of 276,873 chromatograms were obtained from proteome by mass spectrometry analysis, and 71,762 effective chromatograms were matched by Maxquant (v1.6.15.0), and the data were filtered by search database analysis. The accuracy FDR of spectrum, peptide, and protein identification was set at 1%. The identification protein needed to contain at least one unique peptide segment. We compared the relative quantitative values of protein for t-tests, and calculated the corresponding p-value as the significance index. The default p-value is ≤0.05. When the p value was less than 0.05, the differential expression level was significantly upregulated when it exceeded 1.3, and significantly downregulated when it was less than 1/1.3.
Transcriptome sequencing used fold change ≥ 2 and false discovery rate < 0.01 as the screening criteria for differential genes. FDR was obtained by correcting the difference significance p-value. In the correlation analysis between transcription group and protein group, when Log2 FC > 1 and the verification p value was less than 0.01, it was a significant difference expression of the transcript, and when Log2 FC < −1 and the verification p value was less than 0.01, it was a significant difference expression of the transcript. When the ratio was greater than 1.3 and the p value was less than 0.05, the upregulated protein was significantly differentially expressed. When the ratio was less than 1/1.3 and the p value was less than 0.05, the downregulated protein was significantly differentially expressed. The screening criteria for the results were the same as those used by Ye et al. [35].
2.2.4. Quantitative Real-Time PCR (RT-qPCR) Detection
RNA was extracted and cDNA was reverse-transcribed using the TransZol Up Plus RNA kit (Beijing, China) and One-Step gDNA Removal (TRANS). The Hieff^®^ qPCR SYBR Green Master Mix (High Rox Plus) (Shanghai, China) was employed for the RT-qPCR. The parameter settings and gene expression calculations were the same as those used by Ye et al. [35].
2.2.5. Field Experiment
Six canola varieties (FY730; FY737; FY823; FY958; ZY17; and SY664) were planted in the same field. They were transplanted into the field with Cd > 0.3 mg/kg (Q) and the control with Cd < 0.1 mg/k (Y) at the 5–6 leaf stage and then sampled three times: once every 14 d, i.e., at 14 days (A), 28 days (B), and 42 days (C), respectively, for RT-qPCR. The contents of heavy metals in the two fields are different (Table S3). Urea, KCL and potassium dihydrogen phosphate are used to supplement the nutrients, so as to ensure that the nutrients in the two places are consistent with the field management methods.
2.3. Data Analysis
Each result in this study was the average value of three replicates. IBM SPSS Statistics 25 statistical software (25.0) was used for the correlation analysis of the experimental data.
3. Results
3.1. Performance of Canola NILs at Different Heavy Metal Concentrations
The two canola NILs were treated with different heavy metal contents to compare their tolerance levels (Figure 1). The results show that at the 100× heavy metal concentration, the germination rate and biomass of F338 were significantly higher than those of F335. However, in the 50× heavy metal concentration experiment, the biomass of both materials exceeded 2.00 g, and the germination rate exceeded 98.00% (Table 2). The results show that low concentrations of heavy metals can promote seed germination, but higher concentrations may have a significant toxic effect on seed germination and root growth. F338 was minimally affected by heavy metal stress, which may be because it has a lower oil content and higher protein content, whereby the heavy metal ions form metal–protein complexes with the functional side-chain groups and are fixed, which reduces the toxicity of the metal ions to cells [36,37].
3.2. Physiological Performance of Canola NILs in Resisting Heavy Metal Stress
The physiological traits of the canola NILs under high-concentration heavy metal stress were compared (Figure 2). F335 and F338 exhibited significant differences in SOD and POD activities: 32.02 mmol/mg and 24.85 mmol/mg for SOD and 71.84 mmol/mg and 63.86 mmol/mg for POD, respectively. Under 100× heavy metal stress, the MDA contents also showed a significant difference of 1.34 times, indicating that heavy metal stress can cause lipid peroxidation. SOD and POD may be used as the main detoxification enzymes for canola seed germination subjected to heavy metal stress [38,39].
3.3. Omics Association Analyses of the Canola NILs in Response to Heavy Metals
The materials under 100× heavy metal stress were used for genomics and proteomics analyses.
3.3.1. Transcriptome Analysis of Canola NILs under 100× Heavy Metal Stress
In total, 9665 DEGs were observed in the transcriptome analysis, of which 4820 genes were downregulated and 4845 were upregulated. The GO and COG analyses showed that the differential genes mainly participated in the redox process and the responses to cadmium ions, salt stress, and cold under high-concentration heavy metal stress, which indicated that the functional genes in canola that responded to heavy metal stress also participated in abiotic stress processes. The findings on protein, zinc ion, and iron ion binding from the molecular function (MF) and cellular component (CC) analysis indicate that heavy metal ions may enter through the inorganic salt absorption channels in canola and then selectively penetrate the cell membrane, which transfers and fixes them to vacuoles and the cytoplasm, thus reducing the toxicity of the heavy metals to the plant. Canola binds related proteins via activation, thus fixing the metal ions and reducing their toxicity (Figure 3). A total of 322 KEGG pathways were significantly enriched in KEGG, among which the most significant difference was in plant hormone signal transduction pathways [40]. The metabolism of starch and sucrose, protein processing in the endoplasmic reticulum, phenylpropionic acid biosynthesis, and plant–pathogen interactions were significantly different, which may explain the decrease in the germination rate and biomass of canola under heavy metal stress [41,42,43,44,45].
Six DEGs were selected and used to verify the results of the transcriptome analysis via RT-qPCR. The results were consistent with the transcriptomics data (Figure 4a), indicating that the transcriptome analysis results were reliable.
3.3.2. iTRAQ Analysis of Canola NILs under 100× Heavy Metal Stress
A total of 276,873 chromatograms were obtained from the iTRAQ analysis, 71,762 effective chromatograms were matched via MaxQuant analysis (v1.6.15.0), and 8925 proteins and 1787 differential proteins were identified (Figure 5a). The GO analysis showed that most DAPs in BP and MF were related to the cell interpretation of hormone antioxidant activity, and most were upregulated DAPs (Figure 5b). The downregulated DAPs mainly manifested as effects on seed and seedling development, further proving that heavy metal stress can cause membrane lipid peroxidation, affect life metabolism, and reduce biomass in plants. Twenty-six differential pathways were enriched in the KEGG pathway analysis. Compared with F335, F338’s downregulation of adenosine was mainly reflected in the degradation and synthesis of oil, indicating that heavy metal stress affects the substance synthesis of canola [41]. The KEGG pathways showed that the upregulated expression of DAPs was mainly focused on the pathways related to photosynthesis and plant hormones.
Six DAPs were selected, and their corresponding genes were used to verify the results of the proteomics analysis via RT-qPCR. The RT-qPCR results were consistent with the corresponding proteomics data (Figure 4b), indicating that the proteomics analysis results were reliable.
3.4. Validation of Transcription and Proteomics Analyses Using Real-Time Quantitative PCR (RT-qPCR)
In this study, 9665 DEGs and 1787 DAPs were identified by transcriptome and proteome, among which 183 DEGs and 11 DAPs were involved in plant hormone signal transduction and the MAPK signal pathway, and most of the related differential genes were members of the auxin response protein and peroxidase family. It has been found that KEGG pathways such as plant hormone signal transduction and the MAPK signal are highly correlated with plant resistance to heavy metals [40,46,47,48]. Comparing the differential genes in KEGG pathway, the expression of differential genes in the starch and sucrose metabolism pathway is greater, which shows that heavy metal stress can inhibit crop substance synthesis, destroy cell infiltration regulation, degrade protein hydrolysis activity, and finally, inhibit seed germination and seedling development [41,44,45]. In this study, under the stress of high concentration of heavy metals, seed germination was inhibited, and the time to enter the seedling stage after germination was prolonged, or even directly died, resulting in a decrease in biomass, which was consistent with the research results of Seneviratne and others [49]. In the early stage of seed germination, heavy metals inhibited the hydrolysis of carbohydrates and the transfer of hydrolyzed sugars, resulting in slow seedling growth [49].
Comprehensive analysis showed that genes with consistent expression trends in the transcriptome and proteome were mainly involved in pathways such as peroxisome metabolism, enzymatic activity, amino sugar and sugar metabolism, and startup and sugar metabolism. This indicates that crops can resist heavy metal stress and reduce its toxic effects by regulating the expression of key genes involved in metabolic processes, enzymatic activity, and signal transduction [50]. Heavy metal stress during plant growth affects related pathways such as biosynthesis, substance metabolism, and signal transduction (Table S2). In total, 23 DEGs and 27 DAPs were involved in phenylpropanoid biosynthesis, while 16 DEGs and 22 DAPs were involved in starch and sucrose metabolism. Plants respond to heavy metal stress via plant hormone signal transduction and the MAPK signaling pathway [50,51,52]. In the high-concentration heavy metal stress experiments, 56 DEGs in F338 and F335 were involved in plant hormone signal transduction, while 5 DEGs and 10 DAPs were involved in the MAPK signaling pathway (Table S2). The DEGs and DAPs in the related pathways were mainly members of the auxin-responsive protein and peroxidase families. Studies have shown that auxin-responsive protein and peroxidase are associated with plant resistance to heavy metal toxicity [53,54]. In the transcriptome analysis, 45 genes in the peroxidase (POX) family and 24 genes in the auxin-responsive protein ARF family were differentially expressed. Most of the genes in the auxin-responsive protein family were related to the expression of indole acetic acid (IAA) and SAUR. The SAUR-related genes and most of the 12 IAA-related genes were downregulated compared with the control group.
Five peroxidase genes were co-expressed in the transcriptome and proteome with significant differences. Two SAUR-related genes, two IAA-related genes, and one POX gene were examined using RT-qPCR. The results showed that the expression trends of the two SAUR-related genes BnaC04g00740D (SAUR32) and BnaC08g30850D (SAUR36) were the same in the transcriptome and proteome, and there were significant differences between F335 and F338, which indicated that they may be the key genes for plant resistance to heavy metal stress (Figure 4c). The small auxin-upregulated gene (SAUR) family is one of the main early auxin-responsive gene families found in higher plants and plays a central role in auxin-induced acidic growth. It can also be independently regulated by various other hormonal pathways and tissue-specific transcriptional factors [55,56]. SAUR36 is associated with senescence in plants [57,58], and its overexpression may cause slower hypocotyl growth and the disappearance of apical hooklet formation [59].
3.5. The Functional Verification of SAUR Genes
The RT-qPCR analysis showed that in the first three periods, the expression level of SAUR36 in the sample from Field Q in Period A and the other periods was 27.82 times higher and more than 2.5 times higher than that in Field Y, respectively, especially in the canola of FY958 (Figure 6). A difference in SAUR36 expression of 1.07–13.20 times was observed in the other materials. This indicates that canola can tolerate heavy metal stress and maintain life activities by enhancing the expression of SAUR36 [55].
4. Discussion
4.1. Effects of Heavy Metal Stress on Physiological Performance of Canola
Upon entering plants, heavy metals promote the production of reactive oxygen species (ROS). The increase in the level of ROS leads to membrane lipid peroxidation and a large amount of O^2−^ accumulation in the cells, thus destroying the redox steady state of the cells [60,61] and plant metabolism and physiological responses [62,63]. This study found significant differences in SOD activity, POD activity, and MDA content between F335 and F338 under high-concentration heavy metal stress. The difference in SOD was the largest; therefore, SOD may be the key enzyme related to heavy metal stress. Plants can convert a large amount of O^2−^ produced via peroxidation into H_2_O_2_, which is then decomposed into H_2_O and O_2_, thus reducing the toxicity of heavy metals in the plants by enhancing SOD activity [38]. Studies have shown that SOD plays an important role in plant growth and development under stress [39,64,65,66,67]. In this study, SOD was the main heavy metal stress-resistant enzyme at the early growth stage of canola [68], and it mainly cooperated with POD in eliminating excessive ROS caused by heavy metal stress. Gokul et al. [68] found that SOD activity affected the heavy metal stress tolerance of Brassica napus. Yu et al. [69] revealed the function of SOD in the interaction between plants and abiotic stress, and SOD played an important role in plants’ tolerance to Cd stress. At present, SOD has been widely studied in the neighborhood where plants tolerate abiotic stress. For example, a whole-genome study identified that the SOD gene in canola was significantly expressed under abiotic stress [70]. Various stress-tolerant crops have been developed by modifying the SOD gene using transgenic methods [71]. SOD may be helpful for screening crops with heavy metal stress tolerance.
4.2. The Key Genes for Heavy Metal Stress Tolerance in Canola
A total of 183 DEGs and 11 DAPs involved in plant hormone signal transduction were found in this study. The MAPK signaling pathway was mostly related to the auxin-responsive protein and peroxidase family. Studies have shown that IAA can reduce the toxicity of heavy metals by reducing their absorption and increasing plant antioxidant capacity [54,72,73,74]. Overexpression of SAUR regulates cell wall acidification to induce plant growth [75,76]. Some histidine-rich regions at the N- and C-terminals of SAURs can also bind to metals [77] and may thus enhance plant environmental adaptability [45]. The expression of the SAUR gene is associated with tolerance to cold and salt stress [78]. SAURs are involved in the regulation of adaptive growth under abiotic stress and play an important role in plant adaptation to drought stress [79]. Current research shows the SAUR gene’s importance in regulating dynamic adaptive growth [55]. Qiu et al. [80] studied the function of the SAUR gene in Arabidopsis thaliana with CRISPR/Cas9 SAUR gene-editing technology, and the results showed that SAUR is an abscisic acid (ABA)-induced gene that regulates cell amplification, ion homeostasis, and plant salt tolerance. Many hormones and stress response elements exist in the promoter region of SAURs. The expression of SAURs may be induced by abiotic stress and exogenous hormones, which participate in the complex physiological processes in plants resisting abiotic stress [81]. SAUR36 has been found to play a vital role in plant senescence [58,82], regulating seed germination [83], promoting plant root growth [84], and enhancing plant resistance, such as salinity tolerance [84,85], waterlogging tolerance [86], etc. In this study, the expression of SAUR36 in the field was more significant than that of SAUR32, and the gene expression increased with the higher toxicity of heavy metals to canola, indicating that SAUR36 may play an important role in protecting plants from heavy metal stress. There is little research on the SAUR gene’s function in tolerating abiotic stress, and the research on SAUR36 mostly focuses on plant antiaging [87]. The results of this study show that SAUR36 may be a key gene for heavy metal stress tolerance in canola. Comparing the expression of SAUR36 in different materials may be a promising method to identify new materials with heavy metal stress tolerance for canola breeding.
5. Conclusions
In this study, the difference in the germination rate and biomass of the near-isogenic canola lines F335 and F338 under 100× heavy metal stress reached 1.69 times and 3.34 times. SOD activity was significantly different under high-concentration heavy metal stress, reaching 1.29 times. SOD might be the key enzyme for the early growth of canola while tolerating heavy metal toxicity. A total of 9665 EDGs and 1787 DAPs were obtained with transcriptome and proteome association analyses, respectively. The expressed DEGs and DAPs were mainly involved in the pathways related to photosynthesis, plant hormones, and plant hormone signal transduction, and most of them were members of the auxin-reactive protein and peroxidase family, especially the SAUR32 and SAUR36 genes. The expression level of SAUR36 in different canola materials was significantly different between the experimental field and the control, and the highest expression level difference was 27.82 times. The expression level of SAUR36 and activity of SOD may be useful for screening canola materials with heavy metal stress tolerance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Meilke K.D. Griffith G.R. An application of the Market Share Approach to the demand for soyabean and rapeseed oil Eur. Rev. Agric. Econ.19818859710.1093/erae/8.1.85 · doi ↗
- 2Shahid M. Cai G. Zu F. Zhao Q. Qasim M.U. Hong Y. Fan C. Zhou Y. Comparative Transcriptome Analysis of Developing Seeds and Silique Wall Reveals Dynamic Transcription Networks for Effective Oil Production in Brassica napus L.Int. J. Mol. Sci.201920198210.3390/ijms 2008198231018533 PMC 6515390 · doi ↗ · pubmed ↗
- 3Yan L. Shah T. Cheng Y. LÜY. Zhang X.K. Zou X.L. Physiological and molecular responses to cold stress in rapeseed (Brassica napus L.)J. Integr. Agric.2019182742275210.1016/s 2095-3119(18)62147-1 · doi ↗
- 4Deng J. Li W. Xu W. He Z. Tan X. Correlation and the concentrations of Pb, Cd, Hg and As in vegetables and soils of Chongqing, China Environ. Geochem. Health 2021432357237610.1007/s 10653-020-00606-232524341 · doi ↗ · pubmed ↗
- 5Guo Z.H. Song J. Xiao X.Y. Ming H. Miao X.F. Wang F.Y. Spatial distribution and environmental characterization of sediment-associated metals from middle-downstream of Xiangjiang River, southern China J. Cent. South Univ. Technol.201017687810.1007/s 11771-010-0013-7 · doi ↗
- 6Asad S.A. Farooq M. Afzal A. West H. Integrated phytobial heavy metal remediation strategies for a sustainable clean environment-A review Chemosphere 201921792594110.1016/j.chemosphere.2018.11.02130586789 · doi ↗ · pubmed ↗
- 7Munir O. Ersin Y. Salih G. Serdal Aksoy A. Plants as Biomonitors of Trace Elements Pollution in Soil Trace Elements as Contaminants and Nutrients: Consequences in Ecosystems and Human Health John Wiley & Sons Hoboken, NJ, USA 200872374410.1002/9780470370124.ch 28 · doi ↗
- 8Liu J.G. Liang J.S. Li K.Q. Zhang Z.J. Yu B.Y. Lu X.L. Yang J.C. Zhu Q.S. Correlations between cadmium and mineral nutrients in absorption and accumulation in various genotypes of rice under cadmium stress Chemosphere 2003521467147310.1016/S 0045-6535(03)00484-312867177 · doi ↗ · pubmed ↗