Mannose-Presenting “Glyco-Colicins” Convert the Bacterial Cell Surface into a Multivalent Adsorption Site for Adherent Bacteria
Natasha E. Hatton, Joe Nabarro, Nicholas D. J. Yates, Alison Parkin, Laurence G. Wilson, Christoph G. Baumann, Martin A. Fascione

TL;DR
Researchers developed a new method using 'glyco-colicins' to prevent bacteria from sticking to surfaces, which could help combat antibiotic resistance.
Contribution
The study introduces glyco-colicins as a novel antiadhesion strategy leveraging multivalent interactions.
Findings
Glyco-colicins bind to E. coli and induce aggregation by presenting mannose on the bacterial surface.
This approach uses bacteria as a platform to block adherence via FimH adhesin interactions.
The method offers a nonbactericidal way to inhibit bacterial adhesion and potentially reduce infection risks.
Abstract
Biofilm formation is integral to the pathogenesis of numerous adherent bacteria and contributes to antimicrobial resistance (AMR). The rising threat of AMR means the need to develop novel nonbactericidal antiadhesion approaches against such bacteria is more urgent than ever. Both adherent-invasive Escherichia coli (AIEC, implicated in inflammatory bowel disease) and uropathogenic E. coli (UPEC, responsible for ∼80% of urinary tract infections) adhere to terminal mannose sugars on epithelial glycoproteins through the FimH adhesin on their type 1 pilus. Although mannose-based inhibitors have previously been explored to inhibit binding of adherent bacteria to epithelial cells, this approach has been limited by monovalent carbohydrate–protein interactions. Herein, we pioneer a novel approach to this problem through the preparation of colicin E9 bioconjugates that bind to the abundant BtuB…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
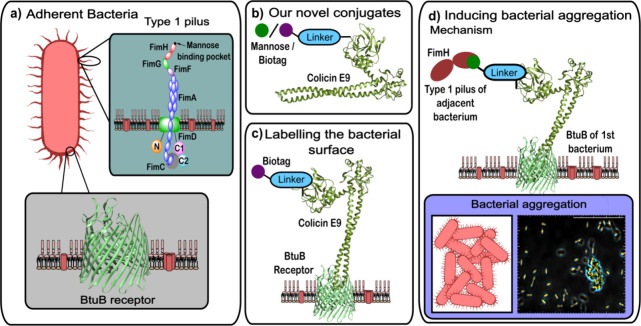 Figure 1
Figure 1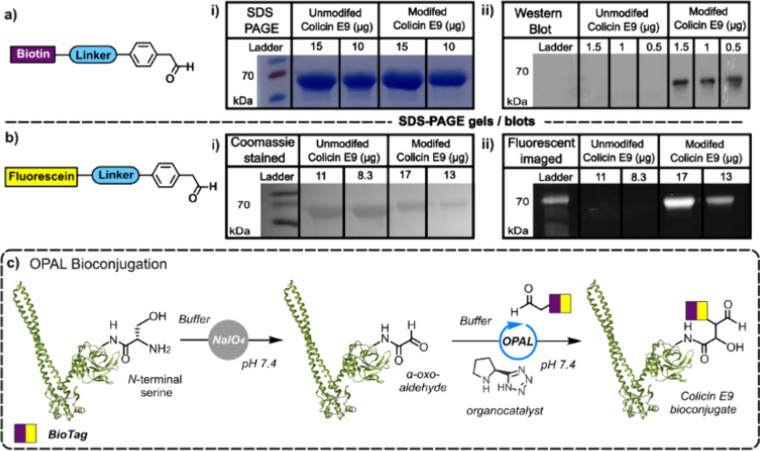 Figure 2
Figure 2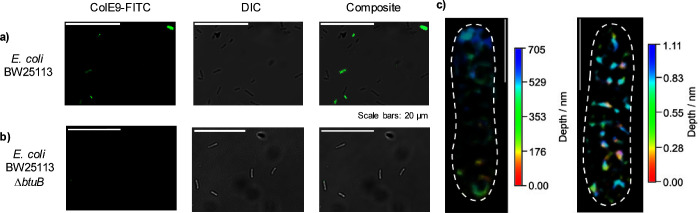 Figure 3
Figure 3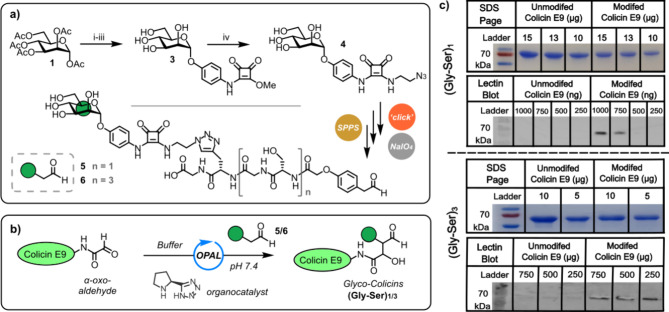 Figure 4
Figure 4 Figure 5
Figure 5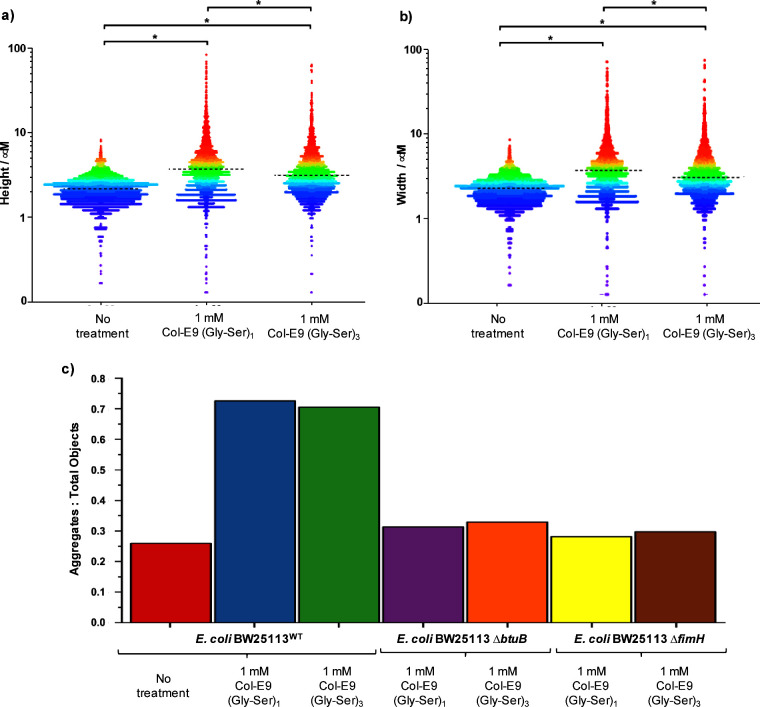 Figure 6
Figure 6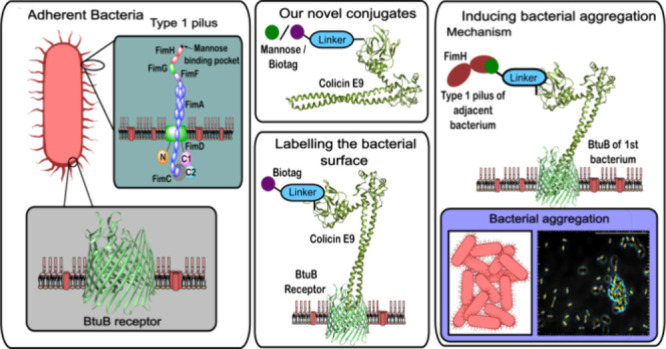 Figure 7
Figure 7- —UK Research and Innovation10.13039/100014013
- —Engineering and Physical Sciences Research Council10.13039/501100000266
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMedieval Iberian Studies
Many pathogenic bacteria, including Escherichia coli, Pseudomonas aeruginosa, and Staphylococcus aureus, have the ability to form biofilms,^1^ which can provide increased protection from antibiotics^2^ and the host immune system^3^ and are thus implicated in pathogenesis.^4^ Adhesion is one of the first steps in biofilm formation^5^ and is mediated by adhesins located on pili or fimbriae on the surface of bacteria. These hairlike appendages are able to recognize and bind to specific receptor moieties on the host cell.^6^ FimH, a two-domain protein located at the terminus of the type 1 pili,^7^ is an adhesin with a mannose-binding pocket located on the end of its lectin domain (Figure 1a).^8,9^ The FimH gene is detectable in 90% of E. coli strains,^10^ and FimH is used by both adherent-invasive E. coli (AIEC)^11^ and uropathogenic E. coli (UPEC)^12^ to adhere to host cells via terminal mannose units on epithelial glycoproteins.^7^ These E. coli strains are well-known pathogens; UPEC is responsible for over 80% of uncomplicated urinary tract infections (UTIs)^13^ with an estimated yearly health care cost greater than $1 billion,^14^ and AIEC has been implicated in inflammatory bowel disease in Crohn’s disease patients.^11^ Antibiotics remain the frontline treatment for these infections;^15^ however, increasing antibiotic resistance rates coupled with international guidelines advising against the use of antibiotics in Crohn’s sufferers^16^ means there is an urgent need to develop nonbactericidal treatments against these adhesive bacteria. This has led to a wealth of elegant research into the use of monomeric α-d-mannopyranoside glycans as antiadhesion therapies that bind to FimH.^17,18^ However, a fundamental limitation of many of these molecules is that they only participate in monovalent binding, whereas in nature, carbohydrate ligand presentation is predominately multivalent,^21−25^ and although impressively constructed mannose-based dendrimers typically have higher potencies than their monovalent counterparts,^23,26−28^ they can also be more challenging to access.^29,30^
We hypothesized that colicin proteins labeled with mannose sugars could form the basis of novel nonbactericidal agents against adhesive bacteria. Colicins are small proteins produced by some E. coli strains, which are toxic to others,^31^ and bind to their target receptors with high (i.e., nanomolar) affinity.^32^ They are promising agents for use in therapeutic contexts as their narrow targeting range could potentially facilitate single-strain targeting,^33^ while the lack of their target receptor on mammalian cells means off-target interactions are unlikely.^34^ Notably, while native colicins are used as offensive weapons against competing E. coli strains,^35^ Penfold et al. demonstrated that the installation of a “top lock” in the R-domain of colicin E9 inhibits toxicity while not adversely affecting the binding of colicin E9 to its target receptor (the BtuB outer membrane protein receptor),^32^ thereby allowing colicin E9 to serve as a nonbactericidal agent for labeling E. coli.^35,36^ Herein, we present the design and synthesis of colicin conjugates presenting functional motifs, such as (i) biotin, (ii) fluorescein, and (iii) α-d-mannopyranosides. We demonstrate that the conjugation of functional motifs to colicin E9 can be achieved by an organocatalyst-mediated protein aldol ligation (OPAL) (Figure 1b) and that appending functional motifs to colicin E9 does not prevent the binding of the protein to its target BtuB receptor (Figure 1c). Furthermore, we demonstrate the premise of using α-d-mannose-presenting colicin E9 neoglycoproteins as heterobiselective binding agents that can induce aggregation in samples of E. coli K12 substr. BW25113 via the simultaneous binding of FimH and BtuB motifs. We propose that the initial high-affinity binding of colicin units to constitutively expressed (∼500 per cell) BtuB receptors (Figure 1d) results in the labeling of E. coli surfaces with multiple α-d-mannose motifs, which are thereafter capable of multivalent binding to FimH adhesins on neighboring bacterial surfaces.
Use of colicin E9 to label E. coli with biologically relevant motifs necessitates the conjugation of the colicin to the desired motifs using a site-specific bioconjugation approach. The N-terminal T domain of colicin E9 has been shown to be a structurally disordered region with a high degree of flexibility and distal to the colicin E9 binding site.^32,37^ We therefore hypothesized that modification at this site would not preclude binding of colicin E9 to its receptor. Conveniently, the N-terminal residue of colicin E9 is a serine, and thus, the installation of a bio-orthogonal α-oxo aldehyde motif at this site was easily achieved through a mild treatment with NaIO_4_ (Figure 2c). This α-oxo aldehyde could subsequently be targeted through OPAL,^38^ a site-specific bioconjugation method previously developed in our lab that uses proline (tetrazole)-based secondary amines as organocatalysts to mediate cross-aldol ligation between an enolizable aldehyde-functionalized probe bearing a biologically relevant motif and a nonenolizable electrophilic α-oxo aldehyde, in this example enabling the desired chemical motif to be appended to the N-terminus of colicin E9 (Figure 2c). To demonstrate that OPAL conjugation to the N-terminus of colicin E9 was possible, a sample of α-oxo aldehyde-functionalized colicin E9 was subjected to OPAL with a biotin-presenting OPAL probe.^39,40^ Successful ligation was validated using both SDS-PAGE gel and Western blot analysis (Figure 2a). Because of the size of colicin E9 (61 kDa), the addition of the biotinylated OPAL probe only results in a subtle observable increase in the mass of the colicin E9 band in SDS-PAGE (Figure 2a,i), yet Western blot visualization using a streptavidin horseradish peroxidase-conjugate that binds biotin noncovalently with femtomolar affinity unequivocally demonstrated that the biotin had been successfully attached (see Figure 2a,ii). To prove that conjugation via OPAL does not prevent binding of colicin E9 to the BtuB receptor, a fluorescein-labeled colicin E9 conjugate was also prepared enabling BtuB binding to be visualized via fluorescence microscopy (Figure 2b). The successful labeling of colicin E9 with fluorescein was also confirmed via SDS-PAGE gel analysis (Figure 2b,i) in which a subtle mass increase can be observed after bioconjugation of the fluorescein-labeled OPAL probe confirmed by fluorescent visualization (Figure 2b,ii).
The fluorescein-labeled conjugate was subsequently incubated with samples of wild-type E. coli BW25113 and BW25113 ΔbtuB. These samples were then imaged via 3D-structured illumination microscopy (3D-SIM) and confocal fluorescence microscopy. Fluorescence signal was observed in wt cells (Figure 3a,c), while no significant fluorescence was observed in ΔbtuB (Figure 3b). This shows specific fluorescein-labeled conjugate binding to BtuB outer membrane protein on the surface of E. coli cells while providing vital evidence that OPAL ligation does not preclude binding to surface BtuB receptor.
Following successful demonstration of colicin E9 OPAL labeling as a reliable method for appending biologically relevant molecules to the surface of E. coli, we hypothesized that a mannose-presenting colicin E9 conjugate could be used to label the surface of E. coli with multiple glycans, thereby effectively turning each bacterium into a mannose-presenting “glyco-dendrimer.” Surface-exposed mannose units may then bind FimH units on nearby E. coli cells in a trans-fashion, thereby resulting in cell-to-cell adhesion and potentially autoaggregation. To test this hypothesis a mannose-bearing OPAL probe first had to be synthesized. Many different mannose scaffolds have previously been investigated for use against UPEC, including biphenyl mannosides,^41−45^ (neo)thiazolylaminomannosides,^46,47^ septanose,^48^ and squarate mannosides.^19,49^ An α-mannose-presenting squarate mannoside scaffold was identified as suitable for appending to an OPAL probe as (i) mannose azide 4 could be synthesized relatively simply in four steps (Figure 4a), (ii) squarate mannosides have been shown in literature to have high affinity for FimH with analogues achieving IC_50_ values in the micromolar range,^50^ and (iii) the squarate ester scaffold allows for easy installation of bioconjugation handles through the substitution of the terminal squarate methoxy group with an appropriate amine.
Mannose azide 4 was synthesized via a BF_3_·OEt_2_-mediated glycosylation of d-mannose pentaacetate 1 with 4-nitrophenol to afford α-aryl mannoside 2 in a 53% yield following purification by recrystallization (Figure 4a). Subsequent deacetylation of this protected sugar was followed by the hydrogenation of the nitro group to an amine group, which was followed by squarate ester coupling to afford mannoside 3. As the installation of a sugar into an OPAL probe had not previously been attempted, we chose to make our α-mannose-presenting squarate mannoside amenable to derivatization through azide–alkyne “click” reactions via the addition of 1-amino-2-azidoethane to afford the novel azide containing mannoside squarate 4. Two OPAL probe precursor peptides with different serine-glycine Gly-Ser linker lengths (n = 1 or 3) were then synthesized via solid-phase peptide synthesis (SPPS) as we anticipated linker length and flexibility may influence the conformational entropy and, therefore, the change in free energy upon binding. The Gly-Ser spacer units were chosen as they enable subtle tailoring of linker length in SPPS while maintaining solubility. These precursor peptides featured an l-propargylglycine residue at their C-termini, which we selectively conjugated to our versatile mannose azide 4 prior to resin cleavage in an adaption of the on-resin Cu-catalyzed “click” method of Sewald and co-workers.^51^ Following cleavage from the resin, a stoichiometric quantity of NaIO_4_ was then used to cleanly and selectively convert the more reactive 1,2-amino alcohol motif present in the OPAL probe precursor peptides S8 and S9 to an aldehyde motif without oxidizing the 1,2-diol of the mannose motif,^52^ thereby affording two activated α-mannose-presenting OPAL probes 5 and 6 for bioconjugation (Figure 4a). After freshly installing N-terminal α-oxo aldehyde functionality into colicin E9, the OPAL probes 5 and 6 were used to construct two α-mannose-presenting glyco-colicins (Gly-Ser)1 and (Gly-Ser)3 with different length linkers (Figure 4b). The success of OPAL conjugation was confirmed through analysis by SDS-PAGE and lectin blotting with concanavalin A (a lectin that binds to terminal α-mannose units) (Figure 4c).^53^ OPAL bioconjugation of probes 5 and 6 to colicin E9 was evident in the lectin blots with no lectin binding to unmodified colicin E9 observed.
To investigate whether the glyco-colicins were capable of inducing aggregation of E. coli, the BW25113 wt strain was cultured in static, anaerobic conditions using glycerol as a primary carbon source in place of glucose to promote phase on transitions resulting in fimH expression. Incubation of the bacteria with increasing concentrations of both (Gly-Ser)1 and (Gly-Ser)3 mannose glyco-colicins was then performed with concentration-dependent bacterial aggregation observed by confocal microscopy for both linker lengths (Figure 5) with no aggregation observed when ΔfimH and ΔbtuB knockout strains were used (Figure S63). The extent of cell aggregation in each sample was quantified by measuring aggregate particle dimensions, surface area and two cross-sectional measurements (termed “width” and height”) across three experimental replicates in wt, ΔfimH, and ΔbtuB cells in the presence and absence of 1 mM of both mannose glyco-colicins (Figure 6a,b). Increasing levels of aggregation were observed in cells incubated with increasing concentrations (10 μM, 100 μM, and 1 mM) of both conjugates, in contrast to ΔfimH and ΔbtuB where no significant aggregation was observed irrespective of conjugate concentration (Figures S63 and S64). The extent and BtuB/FimH dependence of aggregation when using 1 mM glyco-colicin is also clearly evident when the ratio of number of aggregates/total objects (aggregates + single cells) is counted and plotted (Figure 6c). Quantitative assessment of aggregation further reinforced this observation demonstrating unequivocally the efficacy of the two conjugates trigger aggregation in cells presenting FimH and BtuB at their surface with statistically significant positive shifts quantified via application of a Mann–Whitney U test to subject populations in the aggregate surface width, height, and area distributions (Tables S1–S3) observed for wt cells in the presence of 1 mM of both conjugates (Gly-Ser)1. In contrast, no statistically significant change in ΔfimH and ΔbtuB aggregate surface area distributions was observed upon incubation with 1 mM glyco-colicin. Notably, a statistically significant larger aggregate width, height, and area size was also observed for the shorter (Gly-Ser)1 compared with the longer (Gly-Ser)3 linker, potentially indicative of a greater conformational entropy penalty when the more flexible (Gly-Ser)3 glyco-colicin is incorporated into an aggregate. Greater loss of entropic freedom might be anticipated for even longer linkers, which may also be able to bind BtuB and FimH on the same bacterium in a cis-fashion.
In conclusion, we demonstrate that OPAL bioconjugation can be successfully used to append functional handles, such as biotin and fluorescein, to colicin proteins. Furthermore, OPAL ligation does not preclude the binding of colicin E9 to its target BtuB receptor on the surface of E. coli, as evidenced by fluorescent labeling of living bacterial surfaces characterized using fluorescence confocal microscopy and 3D-SIM. This work also establishes that mannose-presenting glyco-colicins can induce autoaggregation in an adherent E. coli strain, which was validated using fluorescence confocal microscopy and quantified statistical analyses of aggregate sizes. Bacterial autoaggregation is a complex process that occurs primarily through either a depletion aggregation mechanism, an entropy-driven process that results in characteristically ordered lateral alignment of bacteria, or a bridging aggregation mechanism where adhesins or other molecules bind neighboring cells together.^54,55^ The latter mechanism results in less-ordered aggregates composed of multiple layers of cells resembling a “stacked-brick” pattern and is characteristic of enteroaggregative E. coli,^55^ a diarrheagenic pathogenic human strain in which neighboring cells are held together by aggregative adhesive fimbriae.^56^ In this study, when using glyco-colicins, similar fimbriae-dependent “stacked-brick”-like disordered aggregation is observed, which we hypothesize is a result of simultaneous binding of conjugates to the BtuB outer membrane receptor on one E. coli bacterium and a FimH adhesin on an adjacent bacterium in a trans-fashion. Thus, highlighting how glyco-colicins can potentially mimic native bacterial autoaggregation mechanisms. Additionally, although formation of large aggregates was observed when using 1 mM glyco-colicins, microscopy studies provide evidence of aggregation at micromolar concentration, which are below the concentration required to reduce bacterial loads in mouse bladder studies using monovalent mannosides,^11^ thereby underlining potential future application of glyco-colicins as antiadhesion therapies. However, pharmacokinetics would need to be carefully considered if oral administration was envisioned, with mannose glycoside containing conjugates likely susceptible to the action of glycosidases.^57,58^ An alternative application for glyco-colicins may instead be as a catheter coating or pretreatment to combat catheter-associated infections, which currently account for 75% of all UTIs.^59^
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bai X.; Nakatsu C. H.; Bhunia A. K. Bacterial Biofilms and Their Implications in Pathogenesis and Food Safety. Foods 2021, 10 (9), 211710.3390/foods 10092117.34574227 PMC 8472614 · doi ↗ · pubmed ↗
- 2Høiby N.; Bjarnsholt T.; Givskov M.; Molin S.; Ciofu O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35 (4), 322–332. 10.1016/j.ijantimicag.2009.12.011.20149602 · doi ↗ · pubmed ↗
- 3Jefferson K. K. What drives bacteria to produce a biofilm?. FEMS Microbiol. Lett. 2004, 236 (2), 163–73. 10.1111/j.1574-6968.2004.tb 09643.x.15251193 · doi ↗ · pubmed ↗
- 4Vestby L. K.; Grønseth T.; Simm R.; Nesse L. L. Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics 2020, 9 (2), 5910.3390/antibiotics 9020059.32028684 PMC 7167820 · doi ↗ · pubmed ↗
- 5Kreve S.; Reis A. C. D. Bacterial adhesion to biomaterials: What regulates this attachment? A review. Jpn. Dent. Sci. Rev. 2021, 57, 85–96. 10.1016/j.jdsr.2021.05.003.34188729 PMC 8215285 · doi ↗ · pubmed ↗
- 6Soto G. E.; Hultgren S. J. Bacterial adhesins: common themes and variations in architecture and assembly. J. Bacteriol. 1999, 181 (4), 1059–71. 10.1128/JB.181.4.1059-1071.1999.9973330 PMC 93481 · doi ↗ · pubmed ↗
- 7Sauer M. M.; Jakob R. P.; Eras J.; Baday S.; ErişD.; Navarra G.; Bernèche S.; Ernst B.; Maier T.; Glockshuber R. Catch-bond mechanism of the bacterial adhesin Fim H. Nat. Commun. 2016, 7 (1), 1073810.1038/ncomms 10738.26948702 PMC 4786642 · doi ↗ · pubmed ↗
- 8Qin J.; Wilson K. A.; Sarkar S.; Heras B.; O’Mara M. L.; Totsika M. Conserved Fim H mutations in the global Escherichia coli ST 131 multi-drug resistant lineage weaken interdomain interactions and alter adhesin function. Comput. Struct. Biotechnol. J. 2022, 20, 4532–4541. 10.1016/j.csbj.2022.08.040.36090810 PMC 9428848 · doi ↗ · pubmed ↗