Molecular, morphological, and morphometric evidence reveal a new, critically endangered rattlepod (Crotalaria, Fabaceae/Leguminosae, Papilionoideae) from tropical China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
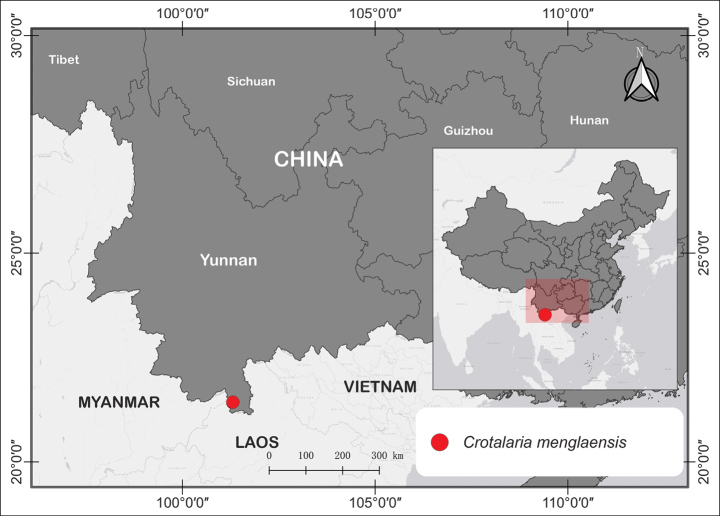 Figure 1
Figure 1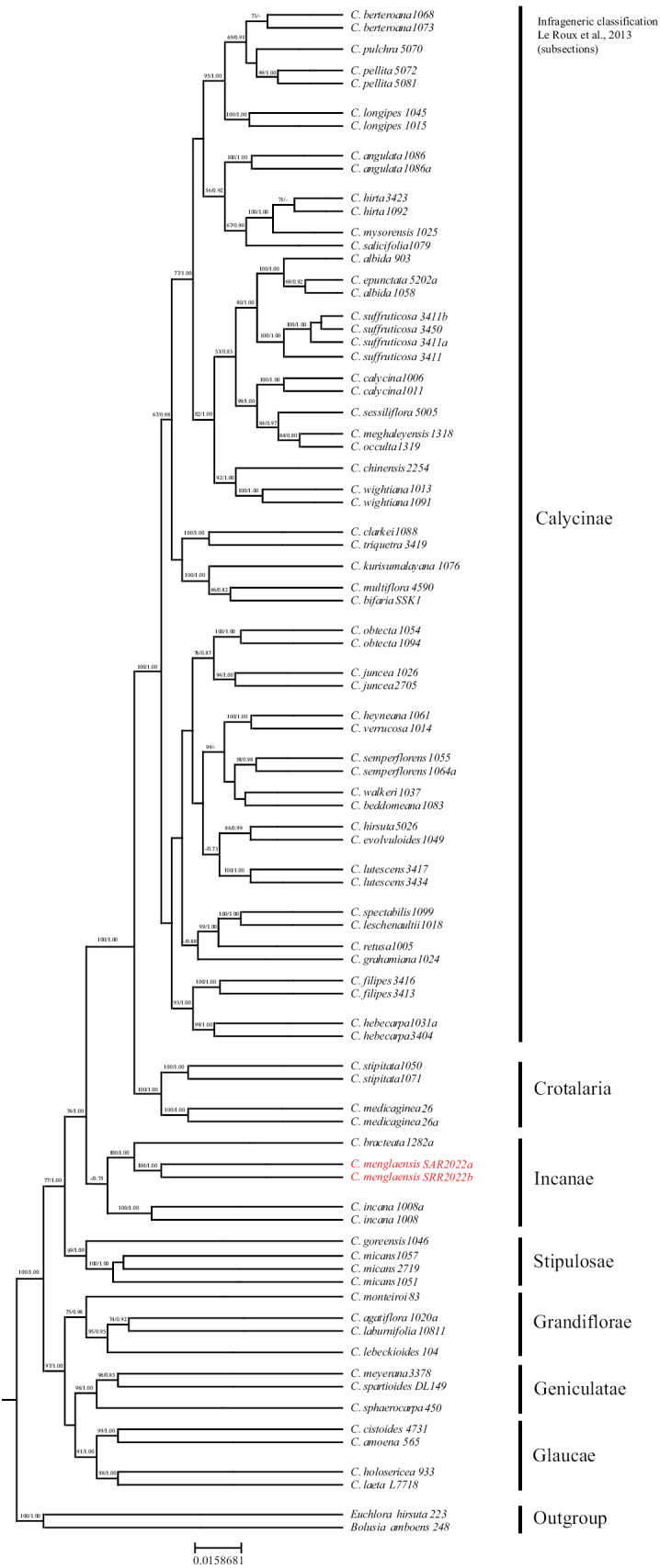 Figure 2
Figure 2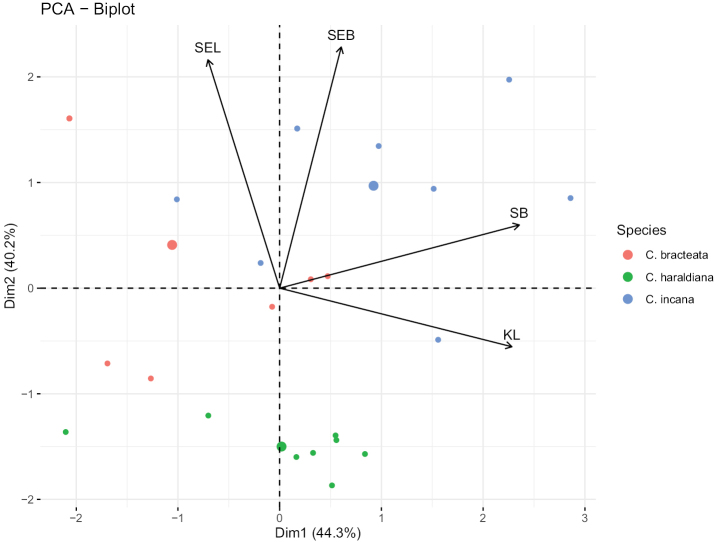 Figure 3
Figure 3 Figure 4
Figure 4| Morphological characters | |||
|---|---|---|---|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Petiole | 23–39 mm | 30–50 mm | 30–50 mm |
|
|
|
|
|
| Stipule | Acicular | Acicular | Acicular |
|
|
|
|
|
|
|
|
|
|
| Leaflet apex | Acute | Obtuse and mucronate | Acuminate |
| Leaflet base | Attenuate | Rounded to broadly cuneate | Attenuate |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Bract size | 1.2–2.0 × 0.6–0.7 mm | 1.5–2.2 × 0.4–0.7 mm | 1–1.5 × 0.1–1 mm |
|
|
|
|
|
|
|
|
|
|
| Bracteole size | 2.7–3.1 × 1.6–1.8 mm | 2–3 mm | 1–2 mm |
| Bracteole surface | Hirsute | Pubescent | Pubescent |
| Bracteole margin | Entire | Slightly involute | Slightly involute |
|
|
|
|
|
| Inflorescence | Terminal or axillary raceme | Terminal or axillary raceme | Axillary raceme or rarely terminal |
|
|
|
|
|
|
|
|
|
|
| Flower colour | Primrose or Strong pale yellow | Yellow | Yellow |
| Flower size | 10–11.9 × 3.3–4 mm | 10 × 5 mm | 7–10 × 9 mm |
| Pedicel length | 0.47 mm | 0.3–0.4 mm | 0.3–0.7 mm |
| Calyx length | 5 mm | 6–8 mm | 5–6 mm |
| Calyx tube | 2.4 mm | 8.1 mm | 7.6 mm |
|
|
|
|
|
| Standard dorsal surface | Hispid at the middle and tomentose at the base | Pubescent | Pilose on the back at the apex |
| Standard size | 88 × 74 mm | 8–1 mm | 9 mm |
|
|
|
|
|
| Standard Claw size | 1.4 mm | 6 mm | 2.4 mm |
|
|
|
|
|
| Wing size | 7.1–7.3 × 2.3–2.9 mm | Staminal sheath ca. 1.5 mm long | 8 mm |
| Wing claw size | 1.52–1.84 × 6.3–0.77 mm | 6–7.5 × 1–2.2 mm | 2–2.7 × 1.1–0.9 mm |
| Keel size | 10.1–15.1 × 4.8 mm | 5.5–6.5 × 2.5–6 mm | 8 mm |
|
|
|
|
|
| Keel alae | Present | Absent | Absent |
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
| Keel claw | 3.4–3.6 × 1.2–1.4 mm | 4.5–4.7 × 1.5–1.7 mm | 3.6–3.9 × 1.1–1.5 mm |
| Androecium size | |||
| Staminal sheath, 7.78 mm | Staminal sheath ca. 4.5 mm | Staminal sheath ca. 3.2–3.7 mm | |
| Filaments 1–1.3 mm long | Filaments 1.7–2.7 mm long | Filaments 5–2.9 mm long | |
| Longer anther, 1.2–1.5 mm | Longer anther, 1–1.4 mm | Longer anther, 1.2–1.7 mm | |
| Shorter anther, 0.5–0.6 mm | Shorter anther, 0.7–0.9 mm | Shorter anther, 0.8–0.9 mm | |
| Gynoecium | Sub sessile | Sessile | Sessile |
| Gynoecium size | 3.3 × 1.5 mm | 2.1 × 1.5 mm | 4.2 × 1.1–6 mm |
|
|
|
|
|
|
|
|
|
|
| Pod stalk | 4.63 mm | 2 mm | 7 mm |
|
|
|
|
|
| Pod size | 14.2–15 × 6–7.7 mm | 20–30 × 6–7.7 mm | 20 × 5–10 mm |
|
|
|
|
|
|
|
|
|
|
| Seed size | 2.2–2.5 × 0.9–1.2 mm | 1.2–5 × 0.32–2 mm | 1.8 × 5 mm |
| Seed colour | Bright citrine | Brown | Brownish black |
| Dimensions | Eigenvalue | Variance | Cumulative Variance Percent (%) |
|---|---|---|---|
| 1 | 1.77 | 44.28 | 44.28 |
| 2 | 1.61 | 40.20 | 84.48 |
| 3 | 0.43 | 10.66 | 95.14 |
| 4 | 0.19 | 4.86 | 100.00 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBotanical Research and Chemistry · Genetic diversity and population structure · Plant Parasitism and Resistance
Introduction
Xishuangbanna, located in the most southwestern part of Yunnan Province and sharing borders with Myanmar and Laos, is well recognized for its rich biodiversity. Its tropical forests play a vital role in global terrestrial biodiversity conservation efforts (Feng et al. 2018). Unfortunately, many plants in Xishuangbanna have recently faced significant threats from deforestation and the establishment of artificial plantations, especially rubber plantations (Brinck et al. 2017; Liu et al. 2017; Yang et al. 2021; Yang et al. 2023). To enable effective protection of the unique and rich diversity of Xishuangbanna, efforts are needed to record species diversity, including that of many species still unknown to science (Chang et al. 2018; Chang et al. 2022; Chen et al. 2022).
Here, we focus on accessions belonging to the legume genus Crotalaria L., which comprises approximately 702 species worldwide (Rockinger et al. 2017; Rather et al. 2018). Its highest species diversity is found in Africa and Madagascar, with an estimated 540 species. It has also expanded to South America and North America, with 64 and 34 species, respectively (Flores et al. 2006; Pandey et al. 2010; Le Roux et al. 2013; Rather et al. 2018). India hosts the largest number of species in Asia (ca. 80 species), followed by Southeast Asian countries, which collectively host 105 species (Lock and Simpson 1991; Rather et al. 2018). Approximately 45 species have been recorded to occur in China, predominantly in Southwest China, including nine endemics and six introduced species (Li et al. 2010). The genus exhibits both annual and perennial life forms and various growth forms (prostrate to erect herbs, undershrubs, robust shrubs, and occasionally small trees) and occupies various habitats, such as open grasslands, roadsides, and forest edges (Polhill 1982; Rather et al. 2018). Crotalaria is characterized by papilionoid flowers, the presence of paired callosities on the standard petal, a rostrate keel, 5 + 5 dimorphic anthers, a hairy style, inflated pods and the presence of pyrrolizidine alkaloids (Le Roux et al. 2013; Rather et al. 2018).
In the present study, several interesting specimens of Crotalaria were collected during field trips to Mengla County in Yunnan Province, China. Initially, some plants observed in the Mengpengzhen area of the Xishuangbanna Dai Autonomous Prefecture could not be assigned to any known taxa. Thus, we considered three priority explanations. The first explanation considered interpreted that these accessions are natural hybrids formed between two sympatrically occurring Crotalaria species, namely, C.bracteata Roxb. ex DC. and C.incana L. However, upon closer examination, the newly discovered species did not match either of these taxa. The plants exhibited differences in numerous characteristics, including plant height, leaflet shape, inflorescence, flower, pod shape, indumentum, and number of seeds per pod, among others. The subsequent discovery of numerous plants during further surveys, which included nearly 50 mature individuals and several immature plants spread over an area of 0.1 km^2^, eliminated the possibility that these plants were hybrids. The second explanation interpreted these accessions as a new distributional record of a known species within the genus Crotalaria L. However, there have been no documented new records for the genus Crotalaria L. This possibility was ruled out after unsuccessful attempts to identify the plants using existing identification keys (Brach and Song 2006; Li et al. 2010). Additionally, we consulted taxonomists at various institutes in China who were unable to recognize the taxa collected. Furthermore, comparisons with verified images of other Crotalaria L. taxa available in the Plant Image Library of China (PPBC; https://ppbc.iplant.cn/) also failed to yield any proper matches. The final explanation considered these plants to represent a new, previously undescribed taxon. This study was designed to confirm this hypothesis by focusing on three lines of evidence, namely, traditional diagnostic morphological character identification, morphometric studies using principal component analyses, and phylogenetic analyses using both plastid matK and nrITS DNA sequences. Finally, we present a comprehensive taxonomic description of this newly discovered Crotalaria L. species, supplemented with taxonomic comments and accompanying photographs.
Materials and methods
Ethics statement
The geographic sites where the newly identified species was found do not coincide with any designated natural conservation areas. Therefore, specific permission for access to these locations was not needed.
Morphological observations
The morphological analysis and description of the newly discovered species were prepared using freshly collected samples. The flowers were preserved in FAA solution (formaldehyde–glacial acetic acid–alcohol) for further studies. They were rehydrated using a mixture of water and detergent to observe the corolla in detail, followed by dissection. Minute corolla features were examined using a Stemi 305 binocular microscope. Morphological terminology adhered to the standards set by Harris and Harris (2001), Hickey and King (2007) for vegetative characters, Hewson (1990) for indumentum description, and Endress (2010) for inflorescence morphology. A comparison of the significant morphological features of the new species with those of its allied species C.incana L. and C.bracteata Roxb. ex DC. was performed (Table 1).
The identification of the allied species C.incana L and C.bracteata Roxb. ex DC. was established through previous revisionary and systematic studies (Brach and Song 2006; Ansari 2008; Le Roux et al. 2013) and examinations of their types and other authentic specimens housed in large herbaria, such as PE, KIB, WUK, HITBC, CAL, DUH, FRLH, M, MH, SJC, and SKU. Additionally, virtual images of these species sourced from repositories such as the JSTOR Global Plants (JSTOR 2024), China Virtual Herbarium (Chinese Virtual Herbarium 2024), Flora of Pakistan (Eflora of Pakistan 2024), and several other prominent online herbaria (B, BM, BR, E, FI, FOB, G-DC, K, L, LINN, NYBG, P, TUB) were also analysed.
A distribution map was constructed to visualize the geographical distribution of the newly identified species. This map was developed with a foundational base map constructed from Natural Earth (www.naturalearthdata.com) and generated using QGIS version 3.28.2 (QGIS 2021) (Fig. 1).
Map visualizing the only known occurrences of Crotalariamenglaensis S.A.Rather in Mengpeng village of Xishuangbanna Dai Autonomous Prefecture, Yunnan, China (red dot).
Taxa sampling for molecular study
Fresh and disease-free leaves were collected from specimens in the field and promptly dried using silica gel to facilitate subsequent DNA extraction. The voucher specimens were preserved at the Herbarium of Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences (HITBC), and detailed information about each sample is provided in Suppl. material 1. The analysis included a total of 81 accessions, covering both the ITS region and the plastid marker matK. Additionally, this dataset included two outgroup sequences from Bolusiaamboensis and Euchlorahirsuta, as well as nine publicly available sequences of African Crotalaria sourced from the NCBI databases (https://www.ncbi.nlm.nih.gov). Overall, our dataset comprised 81 individuals, representing 56 distinct Crotalaria species (Suppl. material 1).
DNA Extraction, PCR Amplification and Sequencing
Genomic DNA was isolated using a DNeasy Plant Mini Kit (Qiagen, Amsterdam, The Netherlands) following the manufacturer’s protocol. The DNA quantity was confirmed via 0.8% agarose gel electrophoresis, and its concentration was determined using a SmartSpec^TM^ Plus Spectrophotometer (Bio-Rad, Hercules, CA, United States). Before amplification, the DNA samples were stored at -20 °C. Polymerase chain reactions (PCR) were performed in a 25 µL reaction volume comprising 2.5 µL of 10× buffer with 2 mM MgCl2, 1 U of Taq DNA polymerase, 1 µL of dNTPs (0.125 mM), 1 µL of each primer (5 pM), and 30–50 ng of total DNA. Nuclease-free water was added to reach the final volume. The optimal PCR conditions and detailed primer information are listed in Suppl. material 2. PCR products were visualized by electrophoresis on 0.8% agarose gels, followed by purification using BioMed multifunctional DNA fragment purification recovery kits (Beijing, China). The purified products were sequenced using the same primers used for PCR amplification. Sequencing was conducted on an ABI 3730 automated sequencer at Sangon Biotech, Shanghai, China.
Sequence alignment and data analysis
To ensure the accuracy and authenticity of the sequences, the original trace files were subjected to rigorous validation through web-based BLASTn searches on the NCBI platform. We conducted sequence alignment in Geneious version 8.1.7, which included trimming, visual inspection, and manual adjustments (Kearse et al. 2012). The trimming parameters were set to an error probability of 0.1 per base and a quality threshold of 20, allowing the removal of any low-quality base calls at the 5’ and 3’ ends of the sequenced PCR products. Each gene was aligned separately using MUSCLE (Edgar 2004) within Geneious. To improve alignment quality and accuracy, ambiguous regions were trimmed using Gblocks v0.91b (Castresana 2000). Individual alignments were then concatenated to create a two-gene alignment for all 81 samples. Microsatellite repeats were excluded, and gaps were considered as missing data. Phylogenetic analyses were performed using the standard-maximum-likelihood (ML) method with IQ-TREE (Nguyen et al. 2015), Bayesian analysis with MrBayes (Ronquist et al. 2012), and the optimal nucleotide substitution model was determined with ModelFinder (Kalyaanamoorthy et al. 2017). Branch support in the ML tree was assessed through 10,000 ultrafast bootstrap replicates (Minh et al. 2013). All procedures were executed using PhyloSuite v1.2.2 (Zhang et al. 2020). The nrITS region of the newly discovered species yielded a 715 bp sequence, while the matK region produced a 775 bp sequence. Concatenating these sequences generated a 1490 bp sequence-aligned matrix, with a total of 1690 characters across 81 accessions. The new species, C.menglaensis, differed from C.incana and C.bracteata by 12 nucleotide substitutions and one inversion at site 601 in both the ITS and matK regions. Additionally, it showed three insertions, with lengths of 3 and 12 at sites 420–422 and 795–806, respectively, compared to its closest relatives. The multiple sequence alignment was submitted to TreeBASE with ID 31180.
Morphometric analyses
To assess potential differences between the new species and their closest relatives and to determine which traits were most relevant for their identification, we conducted a principal component analysis (PCA) using the “factoextra” package in R version 4.3.0 (Kassambara and Mundt 2020; R Core Team 2023) with a significance level set at 5%. We examined three to five specimens of C.menglaensis S.A.Rather, C.incana L., and C.bracteata Roxb. ex DC. The length and width of leaflets, flowers, standards, wings, keels, seeds and pods were measured (Suppl. material 3). Correlation analysis was performed to eliminate highly correlated traits (r > 0.71) using the “corrplot” package in R version 4.3.0 (Wei and Simko 2021; R Core Team 2023). In total, four traits were retained for the PCA biplot analysis: keel length (KL), standard width (SW), seed width (SEW), and seed length (SEL) (Suppl. material 3).
Results and discussion
The proposed new species, Crotalariamenglaensis S.A.Rather, resembles C.incana L. and C.bracteata Roxb. Ex DC. However, it differs from the former in several aspects. It has an ovate to oblanceolate leaflet shape with a pubescent leaf surface, an obovate-orbicular standard shape, a straight keel beak, and an elliptical to oblong pod shape. It differs from the latter in having a stem surface covered with white hairs, a pilose bract surface, a notched standard apex, planar callosity, an angled keel shape, and a tomentose pod indumentum. A comprehensive morphological comparison is presented in Table 1 to elucidate the distinctions between the new taxon and its closest relatives.
The maximum likelihood (ML) and Bayesian tree phylogenies showed congruent topologies (Fig. 2). The phylogenetic tree identified seven major clades, corresponding to seven of the 11 sections proposed by Le Roux et al. (2013). These seven major clades (i.e., Calycinae, Crotalaria, Incanae, Stipulosae, Grandiflorae, Geniculatae and Glaucae) had bootstrap values greater than 80%. These clades are consistent with previous phylogenetic analyses (Subramaniam et al. 2013; Rather et al. 2018). Furthermore, phylogenetic analysis strongly supported the monophyletic status of the genus (100% BS). The phylogeny places the newly discovered species C.menglaensis S.A.Rather within a separate clade, supporting its distinction from other allied species included (Fig. 2). C.menglaensis S.A.Rather forms a distinct clade with C.bracteata Roxb. Ex DC. And C.incana L. (100% BS). Additionally, C.menglaensis S.A.Rather and C.bracteata Roxb. ex DC. form a sister clade with strong support (100% BS), and in turn, they are sisters of C.incana L. (100% BS) (Fig. 2).
Phylogenetic hypothesis of the genus Crotalaria based on the concatenated matrix including matK and nrITS sequences constructed via maximum likelihood as implemented in IQ Tree. Bootstrap values are printed above the branches. Since Bayesian analyses resulted in almost the same topology, only the ML tree made is presented here. The new species Crotalariamenglaensis S.A.Rather was marked in red. The names on the right side of the phylogeny correspond to the infrageneric classification of the genus Crotalaria by Le Roux et al. (2013).
Morphometric analyses based on principal component analysis using Pearson’s coefficient were employed to identify significant morphological characteristics that facilitated differentiation between the new species and its closest relatives based on gross morphology (Fig. 3, Suppl. material 4, Table 2). These morphometric analyses have proven highly valuable for elucidating correlations among variables or distances among groups and assessing the significance of each character. Fig. 3 illustrates the significant characteristic ratios that contribute to the uniqueness of the new species. Following Pearson’s correlation analysis, highly correlated traits were excluded, and four traits were used for statistical analysis (Fig. 3, Suppl. material 4, Table 2). The results showed significant differences in morphological characters, including keel length (KL), standard width (SW), seed width (SEW), and seed length (SEL), compared to those of allied species (Fig. 3). Dimension 1 of the PCA explained the greatest variance, followed by dimensions 2, 3, and 4, which collectively accounted for 84.48% of the variation, a substantial proportion (Fig. 3, Suppl. material 4, Table 2).
Scatter plot visualizing Dim1 and Dim2 from the principal component analyses based on the assembled morphological trait variables and accessions of the three species nested in the Incanae clade (see Fig. 2), namely, C.incana L., C.bracteata Roxb. ex DC. and the new species Crotalariamenglaensis S.A.Rather. Dim1 explained 44.3% of the variation, whereas DIM2 explained 40.2%. The vectors corresponded to KL = keel length, SW - standard width, SEW - seed width, and SEL - seed length.
Taxonomic treatment
Crotalaria
menglaensis
Taxon classificationPlantaeFabalesFabaceae
S.A. Rather sp. nov.
9C06759F-2FE8-581B-9FBA-ADED8DAAA956
urn:lsid:ipni.org:names:77343398-1
Type.
China. Yunnan: Xishuangbanna Dai Autonomous Prefecture, Mengla County, Mengpengzhen., 21°26'57.42"N, 101°18' 31.49"E, alt. 577 m, 23 November 2022, SAR 202305 (holotype HITBC! isotypes KIB! PE! DUH! CAL!).
Diagnosis.
The new species is similar to two sympatrically occurring species, C.incana L. and C.bracteata Roxb. ex DC. However, C.menglaensis S.A.Rather differs from the former and latter in its height, 0.5 m (vs 1 vs 60–1.20); stem surface, pubescent with white hairs (vs pubescent brownish vs densely brownish yellow); bract surface, pilose (vs glabrous vs glabrous); leaflet shape, ovate to oblanceolate (vs elliptic obovate, or suborbicular vs narrowly elliptic) ; leaflet surface, pubescent (vs glabrous vs sparsely pilose); standard shape, obovate-orbicular (vs elliptic vs oblong); planar callosities (vs ridge vs ridge); keel shape, angled (vs subangled vs subangled); keel beak, straight (vs spirally twisted up to 90° vs slightly incurved); pod shape, elliptic to oblong (vs fusiform vs ellipsoid-fusiform); and pod indumentum tomentose (vs rusty pilose vs densely rusty pubescent).
Crotalariamenglaensis S.A.Rather A habit B plant twigs with leaves and flowers C inflorescence with flowers D inflorescences with flowers and fruits E flower in dorsal, lateral, and ventral views F calyx showing the dorsal and ventral surfaces G standard adaxial surface H standard abaxial surface with paired planar callosity pairs at the base with white silky pubescence I wing petals with prominent cavae and a distinct claw J adaxial and abaxial surfaces of keel petals, beak not twisted, pubescence along the margins from the middle to the base of the keel petal K anthers monodelphous, 10 dimorphic anthers (common to all the species within the genus) L gynoecium showing the ovary, style, and stigma M pod in ventral, dorsal, and lateral views N pod splitted longitudinally with young seeds.
Description.
Stiff and erect herbs, ca. 0.5 m tall. Stems terete and densely pubescent. Stipules acicular. Leaves trifoliolate, alternate, petiole up to 30 mm long, lamina ovate to oblanceolate, 30–80 × 21–31 mm, terminal leaflet larger than the lateral ones, attenuated at the base, acute at apex, margin entire with puberulent indumentum, adaxial surface glabrescent, abaxial surface pubescent. Inflorescence a terminal or axillary raceme, a terminal raceme 80–120 mm bearing up to 12 flowers, and an axillary raceme 110–170 mm bearing up to 47 florets. Flower 10–11.9 × 3.3–4 mm. Bract lanceolate, 1.2–2 × 0. 6–0.7 mm covered with white pilose hairs inserted at the base of a pedicel. Pedicel ca. 4.7 mm, pubescent, reflexed downwards; bracteole ovate to obovate with an asymmetric base, 2.7–3.1 × 1.6–1.8 mm, hirsute, margin entire. Calyx 5-lobed, calyx tube ca. 2.4 × 2.9 mm, oblong-lanceolate, 2.2–2.9 × 0.4–0.71 mm, apex attenuate, densely ciliate along margins. Corolla primrose or strongly pale yellow, exserted beyond calyx, obovate-orbicular, ca. 8.8 × 7.4 mm, claw ca. 1.4 mm, with paired planar callosites at the base, ca. 0.6–0. 7 × 0. 7–0.8 mm; wing petals 7.1–7.3 × 2.3–2.9 mm, claw 1.52–1.84 × 6.3–0.77 mm, cavae 4.2–4.4 mm; keel angled, curvature below the middle, claw 3.4–3.6 × 1.2–1.4 mm, glabrous, beak straight. Staminal sheath 7.8 mm; filaments free, glabrous, shorter filament 3.7–6.7 mm, longer filament 7.7–8.0 mm; anthers dimorphic, basifixed ones longer, ensiform, ca. 1.2–1.5 mm, dorsifixed ones shorter, orbicular ca. 0.5–0.6 mm. Ovary sessile, linear, ca. 3.3 × 1.5 mm, inflated, style 8.2 mm long, geniculate, trichomes in a single row; stigma brush-like and contracted, ca. 0. 21 mm long, hairy. Pods elliptic to oblong, 14.2–15 × 6–7.7 mm, tomentose, with persistent style. Seeds 2.2–2.5 × 0.9–1.2 mm, bright citrine, smooth and glossy.
Phenology.
The plants were observed to bear flowers and fruits from October to January.
Etymology.
The specific epithet of the new species “menglaensis” is derived from the type locality of this species.
Distribution and habitat.
Crotalariamenglaensis S.A. Rather is found in grasslands and exposed areas of Mengpeng, Mengla County, within the Xishuangbanna Dai Autonomous Prefecture, Yunnan, China.
Uses.
Locals use the pods of this species as a food source. Additionally, its roots and seeds are utilized in traditional medicine to treat various digestive disorders.
IUCN Red List Category.
This species is exclusively documented in a single location where clustered populations of fewer than 100 mature individuals have been observed. Its habitat is adjacent to roads and agricultural land and is consistently affected by anthropogenic activities such as grazing, deforestation, cultivation, and landscape management. The potential degradation of its natural habitat and restricted geographical range significantly threatens its survival. Therefore, according to the IUCN Standards and Petitions Committee (2019), this species should be considered critically endangered under criteria A4, B2a, C2a, and D1. These criteria denote species facing a very high risk of extinction in the wild.
Additional specimens examined
(paratypes). China, Yunnan. Mengla, in forest, alt. 1600 m, 12 June 2012, Y.M. Shi & W.S. Chen 254655 (KUN). Xishuangbanna, Mengla, in the forest, 1650 m, 16 July 2014, Y.M. Shui & W.S. Chen 245266 (KUN). Xishuangbanna, Mengla, in the forest, 1450 m, 14 August 2016, Z.Y Wen & Z.A Wang 524694 (KUN). Hekou, on forest edges, 1459 m, 25 November 2005, Z.Y. Chang et al. 162458 (KUN). Xishuangbanna, Mengla, in grasslands, 1200 m, 3 August 2007, Z. Y. Chang 445123 (KUN). Xishuangbanna, Mengla, 1180 m, 25 August 2010, Z.Y. Chiang 2005387 (HITBC).
Supplementary Material
XML Treatment for Crotalaria menglaensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ansari AA (2008) Crotalaria L. in India. Bishen Singh Mahendra Pal Singh.
- 2Brach AR Song H (2006) e Floras: New directions for online floras exemplified by the Flora of China Project.Taxon 55(1): 188–192. 10.2307/25065540 · doi ↗
- 3Brinck K Fischer R Groeneveld J Lehmann S Dantas De Paula MPütz S Sexton JO Song D Huth A (2017) High resolution analysis of tropical forest fragmentation and its impact on the global carbon cycle.Nature Communications 8(1): 14855. 10.1038/ncomms 14855 PMC 535786328303883 · doi ↗ · pubmed ↗
- 4Castresana J (2000) Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis.Molecular Biology and Evolution 17(4): 540–552. 10.1093/oxfordjournals.molbev.a 02633410742046 · doi ↗ · pubmed ↗
- 5Chang Y Hori K Murakami N Cao L Lu S Schneider H (2018) Validation of Hymenaspleniumlaterepens (Aspleniaceae): Evidence from morphology and molecular analyses.Phytotaxa 374(4): 277–290. 10.11646/phytotaxa.374.4.1 · doi ↗
- 6Chang Y Zhang G Wang Z Cao L (2022) Molecular and morphological evidence reveals a new fern species of Hymenasplenium (Aspleniaceae) from south and southwestern China.Phyto Keys 211: 93–106. 10.3897/phytokeys.211.9036336760732 PMC 9836484 · doi ↗ · pubmed ↗
- 7Chen K Khine PK Yang Z Schneider H (2022) Historical plant records enlighten the conservation efforts of ferns and lycophytes’ diversity in tropical China. Journal for Nature Conservation 68: 126197. 10.1016/j.jnc.2022.126197 · doi ↗
- 8Chinese Virtual Herbarium (2024) Chinese Virtual Herbarium. https://primulaworld.blogspot.in/n/2015/12/the-Chinese-virtual-herbarium- cvh.html [accessed 05.01. 2024]