Genetic detection and analysis of porcine norovirus in pigs farmed in north Vietnam
Hieu Van Dong, Giang Thi Huong Tran, Amonpun Rattanasrisomporn, Oumaporn Rungsuriyawiboon, Witsanu Rapichai, Jatuporn Rattanasrisomporn

TL;DR
This study identifies and genetically characterizes porcine norovirus in pigs from northern Vietnam, finding it closely related to Chinese strains.
Contribution
The study reports the detection and genetic characterization of porcine norovirus Genotype II.19 in Vietnam.
Findings
Five (4.9%) of 102 fecal samples tested positive for porcine norovirus.
The detected viruses were Genotype II.19 with high nucleotide identity (97.77–99.62%).
The Vietnamese strains were closely related to Chinese porcine norovirus strains from 2009.
Abstract
Norovirus (NoV) causing gastroenteritis symptoms, which has been reported in several hosts, including humans, pigs, and rats. This study was conducted to identify porcine viral infection and to characterize NoV strains from pigs in some provinces in north Vietnam. Totally, 102 fecal samples from diarrheal pigs on farms in six cities and provinces in northern Vietnam during July 2022 to March 2023 were collected. Polymerase chain reaction was used to identify the viral genome. Positive samples were used for nucleotide sequencing of the partial RNA-dependent RNA polymerase gene sequence. Five (4.9 %) positive stool samples were detected from animals farmed in five different farms, with one positive animal identified in each farm. Genetic analysis indicated that nucleotide identity was in the range 97.77–99.62 % among the 5 NoVs in this study. Phylogenetic analysis pointed out that the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
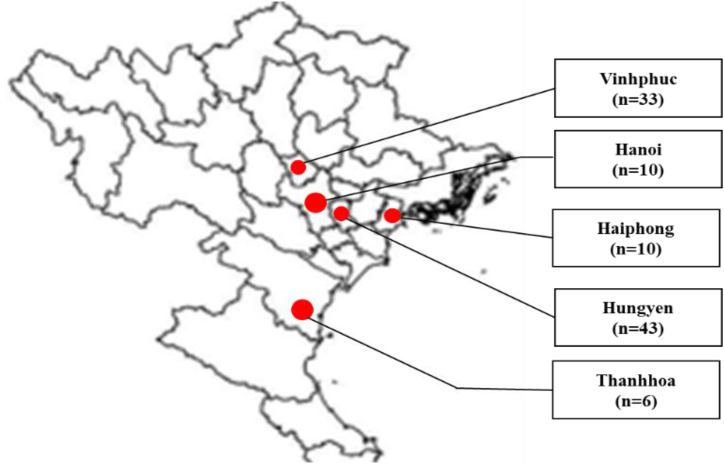 Figure 1
Figure 1 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral gastroenteritis research and epidemiology · Animal Virus Infections Studies · Viral Infections and Immunology Research
Introduction
1
Norovirus (NoV) has been identified as a main agent causing gastroenteritis symptoms in both humans and pigs [1]. In addition, NoVs have been reported as a leading pathogen of food-borne diseases [2]. Transmission routes of NoV can be fecal-oral, air-borne, water-borne, etc. [3]. Recovery of NoVs was successed in samples collected in both humans and animals, consisting of bovine, canine, pig, lion, and murine species [[4], [5], [6]]. NoV strains can infect all ages and cause moderate-to-severe diarrhea.
Analysis of the major capsid protein sequences, VP1, NoV belongs to the family Caliciviridae which forms into 10 genogroups, GI to GX. In addition, within the Caliciviridae family, 49 genotypes of NoV were classified [7]. NoVs are small, round, structured, non-enveloped viruses. The viral particle is approximately 27–38 nm in diameter. Their genomic composition encompasses a positive-sense and single-stranded genomic RNA. The length of viral genome ranges from 7.3 to 7.7 kb. The viral genetic material consists of three open reading frames (ORFs) [8], each responsible for encoding a polyprotein. ORF1 (∼5100 bp) is autocatalytically cleaved by viral protease resulting in the generation of 7 non-structural proteins (NS1–NS7). Among these, NS7 has been encoded for RNA-dependent RNA-polymerase (RdRp). Additionally, ORF2 (1600 bp) is responsible for the synthesis of viral pivotal capsid protein 1 (VP1). VP1 exhibits a structural organization comprising the shell (S) domain. The S singular the virus genome and the P domain contains the P2 domain. The P2 domain indicated as a highly genetic variability. The domain may contribute in inducing neutralization antibodies in the host [9]. In its capsid configuration, the S singular envelops the viral RNA and establishes a connection with the P1 and P2 domains through a hinge. ORF3 (∼720 bp) is responsible for encoding the minor capsid protein VP2, which contributes in viral assembly [10]. The genetics of NoVs are classified based on analysis of the full-length VP1 protein sequences and the ORF1 NS7 region nucleotide sequence [11].
In the field, porcine NoVs (PNoV) are formed within GII. GII NoVs were first reported among American pigs [12]. Subsequently, several countries have recorded the appearance of GII PNoVs in both sick and normal pigs [13]. The PNoV strains circulating in pigs were identified as GII.11, II.18, and II.19 genotypes in many countries in the world [[14], [15], [16], [17], [18], [19]]. Wang et al. reported that the Chinese PNoV strains belonged to three genotypes [12]. Among the three genotypes obtained, one genotype was antigenically and genetically related to the human norovirus [12]. However, the information on GII circulation and the characterization of NoV strains in pigs is still limited.
In Vietnam, NoV infection in humans was reported first in 1999 [20]. The NoV-positive rate in diarrheal children was 5.4 % (72/1339). Genetic analysis of the viral strains indicated that the obtained strains belonging to GII accounted for 73 %, with the remaining strains being GI.4, I.8, II.1, II.3, and II.7 [20]. In 2013, it was reported that 96.5 % (304/315) of the GII virus had been detected from May 2009 to December 2010 [21]. Until now, no data are available on the NoVs present in pigs. Hence, this study was conducted to investigate the presence of NoVs in pigs in north Vietnam.
Materials and methods
2
Ethics statement
2.1
Human participants were not involved in this study. Feces from pigs farmed in north Vietnam were sampled under the acceptance of the Vietnam National University of Agriculture (VNUA). The Committee on Animal Research and Ethics of the VNUA approved the sampling protocol (CARE-2022/08). In order collect samples, permissions of all the pig farm owners were given.
Samples
2.2
Sows, fattening pigs, and piglets farmed in Haiphong (n = 10), Hanoi (n = 10), Vinhphuc (n = 33) Thanhhoa (n = 6), and Hung Yen (n = 43) were identified for sampling (Fig. 1). In total, three to six pigs with clinical sign of diarrhea and dehydration were sampled in each farm. Totally, 102 feces from pigs were sampled and immediately placed in separate sterile tubes, containing 1X phosphate-buffered saline (PBS) between July 2022 and March 2023. Samples were preserved in dry ice and sent to the Faculty of Veterinary Medicine of the Vietnam National University of Agriculture for further investigation. Next, the fecal samples were homogenized in PBS to prepare 10 % homonogenates and preserved at −80 °C.Fig. 1. Geographical locations of sampling in north Vietnam. Locations of provinces and cities for sampling are indicated as red circles. Feces were sampled from pigs with diarrhea and dehydration farmed in Haiphong (n = 10), Hanoi (n = 10), Vinhphuc (n = 33) Thanhhoa (n = 6), and Hung Yen (n = 43).Fig. 1
Extraction of total RNA
2.3
Extraction of RNA was performed by using commercial GeneAll® Ribospin vRD II Kits (GeneAll Biotechnology; Gyeonggi-do, South Korea). The protocol was performed according to instructions of the manufacturer. A volume of 50 μl of distilled water was used to elute RNA. The tube containing total RNA was preserved at −80 °C.
Synthesis of cDNA
2.4
Performance of the cDNA synthesis was carried out by using a M-MLV reverse transcriptase (Invitrogen; Carlsbad, CA, USA). Reactions were conditioned thermally at 25 °C/10 min, 37 °C/60 min, 65 °C/10 min. The cDNA product was preserved at −30 °C.
Performance of PCR and nucleotide sequencing
2.5
PCR was performed to determine the PNoV, transmissible gastroenteritis virus (TGEV), porcine epidemic diarrhea virus (PEDV), porcine rotavirus (PoRoV), and porcine sapovirus genomes in the collected pig feces samples. The specific information of primers for each virus are indicated in Table 1. A partial RNA-dependent RNA polymerase (RdRp) gene region was amplified, previously described elsewhere [22]. PCR reagents were performed using GoTaq® Green Master Mix (Promega) and specifice primers (Table 1), followed by thermal conditions: of 94 °C/5 min, 35 cycles of 94 °C/30 s, 53 °C/60 s, and 72 °C/2.5 min, extension at 72 °C/10 min. The target PCR product of about 319 bp was electrophoresized on a 1.2 % agarose gel. Gel pictures were captured under ultraviolet radiation. Purification of the target PCR products was performed using a GeneClean® II Kit (MP Biomedicals; Santa Ana, CA, USA). The nucleotide sequencing was performed by the 1st BASE Company (Malaysia).Table 1. Primers used for PCR in this study.Table 1. Target virusPrimer nameNucleotide sequence (5' - 3′)PCR product (bp)ReferenceNorovirusP289TGACAATGTAATCATCACCATA319[18]P290GATTACTCCAAGTGGGACTCCACPorcine rotavirusRot3AAAGATGCTAGG GACAAAATTG308[23,24]Rot5TTCAGATTGTGGAGCTATTCCAPorcine epidemic diarrhea virusP1TTCTGAGTCACGAACAGCCA650[24]P2CATATGCAGCCTGCTCTG AATransmissible gastroenteritis virusT1FGTGGTTTTGGTYRTAAATGC859[24]T1RCACTAACCAACGTGGARCTAPorcine SapovirusPSaV-FCTCATCAACCCTTTTGAAAC757[25]PSaV-RAAAGCATGATGTTGTTAGGC
Data analysis
2.6
Clustal W [26] in the BioEdit software [27] was applied to perform alignment and analysis of the nucleotide sequences in this study. The nucleotide homology among obtained sequences and other sequences from GenBank was performed using the Basic Local Alignment Search Tool (BLAST; https://blast.ncbi.nlm.nih.gov/) and GENETYX version 10 software (GENETYX Corp.; Tokyo, Japan) and nucleotide sequences of the Vietnamese PNoV strains in this study and other NoVs (43), Sapovirus (1), and Lagoviruse (2) (Table 2) from the GenBank database were used to construct a phylogenetic tree. A neighbor-joining method with the p-distance model was used to establish the tree in MEGA6 with 1000 replicates of bootstrapping [28]. Sequence data was available in GenBank with accession numbers OR835794–OR835798.Table 2. Description of Norovirus, Sapovirus, and Lagovirus strains used to conduct genetic and phylogenetic characterization in this study.Table 2. NumberGenBank accession No.StrainLocationSourceYearVirus1GU556171.1SII-2-13TaiwanPig2008Norovirus2GU556173.1SII-2-16TaiwanPig2008Norovirus3GU556169.1SII-2-32TaiwanPig2008Norovirus4GQ149616.1sw42ChinaPig2009Norovirus5GQ149615.1sw59ChinaPig2009Norovirus6HQ392821.1Ch6ChinaPig2009Norovirus7AF194184.134NetherlandsPig1998Norovirus8JN644277.1sw50NetherlandsPig2009Norovirus9JN644280.1sw86NetherlandsPig2010Norovirus10MN605619.190186–1ItalyPig2018Norovirus11MN605620.190186–2ItalyPig2018Norovirus12FJ843083.1Vet10-S08108DenmarkPig2009Norovirus13FJ843084.1Vet14-S08257DenmarkPig2009Norovirus14FJ843087.1Vet53-S08140DenmarkPig2009Norovirus15EU448332.1F12-8CanadaPig2005Norovirus16EU448322.1F12-8CanadaPig2009Norovirus17EU448323.1F18-10CanadaPig2009Norovirus18EU448325.1F16-8CanadaPig2009Norovirus19FJ498782.1AB276CanadaPig2009Norovirus20FJ715807.131SloveniaPig2004Norovirus21AB039775.1Saitama U1JapanHuman2002Norovirus22AB039776.1Saitama U3JapanHuman2000Norovirus23AB039777.1Saitama U4JapanHuman2000Norovirus24AB039781.1Saitama U18JapanHuman2002Norovirus25AB039780.1Saitama U25JapanHuman2002Norovirus26AB039782.1Saitama U201JapanHuman2002Norovirus27AY237415.2Mc37JapanHuman2004Norovirus29AB074892.1Sw43JapanPig1997Norovirus30AB126320.1swine43JapanPig2009Norovirus31AF145896.1Camberwell/101922AustraliaHuman1994Norovirus32X86557.1LordsdaleUKHuman2002Norovirus33AY772730.1Neustrelitz260GermanyHuman2000Norovirus34U07611.2Hawaii virusUSAHuman1971Norovirus35AY038599.2VA97207USAHuman1997Norovirus36AY823304.1OH-QW101USAPig2003Norovirus37AY823305.2OH-QW125USAPig2003Norovirus38AY823306.1OH-QW170USAPig2003Norovirus39AY823307.1OH-QW218USAPig2003Norovirus40AF097917.5Newbury2UKBovine1976Norovirus41AF182760.1Cowden virusUSAPig1999Sapovirus42M67473.1RHDV-FRGGermany1991Lagovirus43Z69620.1EBHSV-GDFrance1996Lagovirus
Statistical analysis
2.7
Comparison of rates of the PNoV genome detection according to sampling area, age, and scale of pig farms was performed using Fisher's exact test. A p-value less than 0.05 was used to test for statistically significant differences.
Results
3
Detection of PNoV genome in field samples
3.1
The fecal samples tested were obtained from pigs indicating clinical signs of diarrhea and dehydration. Of 102 pigs examined, the viral genome was detected in five (4.9 %) samples by PCR analysis. The porcine rotavirus, PEDV, TGEV, and porcine sapovirus genome were not detected in these five samples. The PNoV genome-positive rates were 10 %, 10 %, and 6.98 % (p > 0.05) in Hanoi, Haiphong, and Hungyen, respectively, while no positive samples were detected in the remaining two provinces (Thanhhoa and Vinhphuc). Of the 20 pig farms tested, 5 (20 %) were positive for at least one sample to the detection of norovirus. The genome-positive rates were 50 % and 50 % in Hanoi and Haiphong, respectively, and then for Hungyen (37.5 %) (p > 0.05) (Table 3).Table 3. Identification of porcine norovirus genome in fecal samples from pigs in North Vietnam using PCR.Table 3. Province/cityNo. of collected samplesNo. of gene-positive samples (%)No. of farmsNo. of gene-positive farms (%)Hanoi101 (10.00)21 (50.00)Haiphong101 (10.00)21 (50.00)Hungyen433 (6.98)83 (37.50)Thanhhoa60 (0.00)10 (0.00)Vinhphuc330 (0.00)70 (0.00)Total1025 (4.90)205 (25.00)
Based on age, 4 (12.12 %) samples were positive for the PNoV genome in finishing pigs, which was significantly higher (p = 0.02) than for a postweaning pig at ten weeks of age (3.03 %). There were no positive samples in the piglets (<21 days of age) or sows. Three levels of farm scale were classified: level 1 (<100 pigs), level 2 (100–300 pigs), and level 3 (>300 pigs). Three (15 %) samples were positive for the viral genome at farm level 2, which was significantly higher (p = 0.02) than for farm level 1 (3.85 %). No positive samples were detected in pig farms at level 3 (Table 4).Table 4. Identification of PNoV genome in fecal samples of pigs according to pig age and farm scale.Table 4. ParameterAge (Week)Number of tested samplesNumber of gene-positive samples(%)Pig age classNursing<360 (0.00)Postweaning3–12331 (3.03)^a^Finishers18–24334 (12.12)^b^Sows>50300 (0.00)Farm scale< 100522 (3.85)^x^100–300203 (15.00)^y^> 300300 (0.00)Different lowercase superscripts ^a, b, x, and y^ indicate significant differences between groups (p < 0.05).
Genetic and phylogenetic analysis of PNoV strains
3.2
The five Vietnamese strains were named as Vietnam/PNoV/VNUA-06, -22, −35, −40, and −56. Among these Vietnamese viral strains, nucleotide identity was high, ranging from 97.77 % (Vietnam/PNoV/VNUA-35 vs. Vietnam/PNoV/VNUA-40 and VNUA-56) to 99.62 % (Vietnam/PNoV/VNUA-40 vs. Vietnam/PNoV/VNUA-56) (Table 5). Comparisons of the five Vietnamese PNoVs from the current study with other viruses downloaded from the GenBank database indicated that these viruses shared the highest nucleotide identities of 98.88 % (Vietnam/PNoV/VNUA-40 vs. Pig/China/GII/Ch6/2009 (HQ392821.1) and 99.25 % (Vietnam/PNoV/VNUA-06, Vietnam/PNoV/VNUA-35 vs. Pig/China/GII.19/Sw42-09-Ch (GQ149616.1); Vietnam/PNoV/VNUA-22 vs Pig/China/GII/Ch6/2009 (HQ392821.1); Vietnam/PNoV/VNUA-56 vs. Pig/China/GII/Sw59-09-Ch/2009 (GQ149615.1)) (Table 6).Table 5. Comparisons of nucleotide identity of partial RdRp gene (270 bp) among sequences of five Vietnamese PNoV strains.Table 5. Strain nameNucleotide identity (%)Vietnam/PNoV/VNUA-06Vietnam/PNoV/VNUA-22Vietnam/PNoV/VNUA-35Vietnam/PNoV/VNUA-40Vietnam/PNoV/VNUA-56Vietnam/PNoV/VNUA-06100Vietnam/PNoV/VNUA-2298.51100Vietnam/PNoV/VNUA-3599.2599.25100Vietnam/PNoV/VNUA-4098.1498.5197.77100Vietnam/PNoV/VNUA-5698.5198.5197.7799.62100Table 6Comparisons of nucleotide identity of partial RdRp gene (270 bp) of sequences of five Vietnamese PNoV strains with downloaded sequences from GenBank database.Table 6. NumberStrain nameVirus with highest nucleotide identityStrainCountryAccession numberYearIdentity (%)1Vietnam/PNoV/VNUA-06Sw42ChinaGQ149616.1200999.252Vietnam/PNoV/VNUA-22Ch6ChinaHQ392821.1200999.253Vietnam/PNoV/VNUA-35Sw42ChinaGQ149616.1200999.254Vietnam/PNoV/VNUA-40Ch6ChinaHQ392821.1200998.885Vietnam/PNoV/VNUA-56Sw59ChinaGQ149615.1200999.25
In order to construct a phylogenetic tree, the partial RdRp gene sequence (270 bp) of the five viral strains obtained in the current study and 34 other viruses were used as reported previously [29,30]. The phylogenetic tree showed that the Vietnamese strains were PNoVs that could be divided into 2 sub-clusters. These five Vietnamese PNoVs obtained were clustered with Chinese PNoV strains Pig/China/GII.19/sw42/2009 (GQ149616.1), Pig/China/GII/sw59/2009 (GQ149615.1), and Pig/China/GII/Ch6/2009 (HQ392821.1) (Fig. 2), which belonged to Genotype II.19.Fig. 2. Neighbor-joining phylogenetic tree of RdRp gene (270 bp) sequences of current Vietnamese porcine norovirus strains and sequences from GenBank database, where MEGA 6 software [28] was used with 1000 bootstrap replicates and the five Vietnamese viral strains are indicated with filled black circles.Fig. 2
Discussion
4
PNoV circulates among several hosts, including humans and animals (such as pigs, mice, dogs, and sheep) [31]. PNoV infection was reported in several countries in the world, including Vietnam [21]. There are currently only limited Vietnamese PNoV sequences in GenBank. To our knowledge, this study was the first report on PNoV infection in north Vietnam in pigs and on the molecular characteristics of the viral strains in the country.
In this study, the PNoV genome was detected in 5 (4.9 %) out of 102 fecal samples from pigs with signs of diarrhea and dehydration. This rate was somewhat low, being similar to that in several Asian countries, Korea (0.5 %) [32], Taiwan (7.2 %) [33], and Japan (10 %) [34], and in European countries, Slovenia (1.2 %) [35] and Belgium (4.6 %), but lower than in the USA (20 %) [36] and Canada (25 %) [5]. These differences in the rates of PNoV-positive samples could be due to variations in location, time, sensitivity of the identification method, and the number of tested samples. Additionally, of the 20 pig farms tested, the viral genome was detected in 5 (25 %) in north Vietnam, suggesting that PNoV strains may have been affecting the porcine production industry in the country. The present result found that only one fecal sample from each farm was detected to be positive. As mentioned previously, several ways of norovirus transmission were recorded, including fecal-oral or via air-borne routes, etc. [3]. This was due to the fact that sample size, sampling scheme may affect the result number of positive samples in each pig farm. Further study would be perform to expand numbers of samples in each farm, numbers of farms to clarify this point. Previous studies reported that PNoV could be detected in both sick and healthy pigs [33,37]. In experimental study, gnotobiotic piglets infected with PNoV developed mild diarrhea at five day-post inoculation [38]. Affected piglets continued shedding virus during viral infection [29,38]. PNoV infection was detected in asymptomatic swine [33,39]. Until now, no direct evident showed that PNoV plays critical roles on raising of gastroenteritis in natural cases of infection in pigs. Clinical signs of affected pigs were non-specific and it was hard to diagnose. The current study has reported PNoV in diarrheal pigs for the first time in Vietnam. Although PNoV-positive samples were tested and negative for the porcine rotavirus, PEDV, TGEV, and PSaV genomes, other pathogens including viruses, bacteria, etc. would be involved to be examined to clarify the role of PNoV strains. Additional studies should be conducted to isolate viral strains and accessed pathogenicity of the current Vietnamese PNoV strains in this study.
Regarding age of infection, Wang et al. (2006) reported that PNoV infection was detected in fecal samples in 43 (40.95 %) out of 105 samples collected from finishers in the U.S., whereas they could not detect PNoV-positive samples from nursing and postweaning pigs during 2003–2005 [36]. In this study, the PNoV genome was identified in a postweaning pig at ten weeks of age (3.03 %), Finishers (12.12 %), whereas no positive samples were found in piglets (<21 days-old) and sows. Taken together, young piglets could not be susceptible to natural PNoV infection. However, Chao et al. noted that suckling (9.2 %), weaning (5.4 %) and finishing (7 %) pigs farmed in central and southern Taiwan in 2011 were positive for the PNoV genome [33]. Further studies should be conducted to expand sampling numbers to better estimate the age-related percentages of PNoVs among Vietnamese pigs. In addition, current results noted that PNoV-positive rates were different between farm scale, suggesting that management of animals may affect rates of PNoV infection in pig farms. No direct evidence indicated the presence of NoV to be associated with the management of animals in the investigated farms more than to their age in this study.
For genetic characterization, previous studies classified NoVs using the partial or complete ORF1 NS7 region that codes for the partial RdRp protein [30,[23], [40], [41]]. Among genotypes, GII genotype viruses were detected as circulating widely in Asian countries, with strains detected broadly across pig farms [12,24]. Genetic and phylogenetic analyses pointed out that the five Vietnamese PNoVs and other Chinese viruses (Pig/China/GII.19/sw42/2009, Pig/China/GII/sw59/2009, and Pig/China/GII/Ch6/2009) were clustered in a similar genotype and belonged to porcine norovirus genotype II.19 (Fig. 2). The current results obtained suggested that PNoV genotype II continues to be widely spread among Asian countries.
Norovirus spreads and circulates in both humans and animals, which raises a potential risk of a zoonotic disease due to direct close contact. Genotype II strains have been widely identified in humans and several animal species [12,24], with genotypes II.11, II.18, and II.19 being specific for pigs. In the current study, all five PNoV strains were genotype II.19 viruses. We found no relationships between the obtained Vietnamese PNoV strains to human NoVs. Further study should continue to determine any associations among the NoVs circulating in pigs and humans in Vietnam.
PNoV infection was detected not only in diseased pigs but also healthy pigs [33,37]. In this study, only pigs with diarrheal symptoms were studied, but not healthy pigs. These current data may not indicated an overlook of PNoV among pigs in Vietnam. More samples from healthy pigs would be collected and conducted in the future. In term of genetic and phylogenetic characterization, previous studies used the partial and full-length RdRp sequences of PNoV strains characterize the viral strains [29,30,33]. Meanwhile, the full-length VP1 and complete genome were also used to conduct genetic characteristics of the PNoVs [[25], [42], [43]]. In this study, only a short RdRp gene (270 bp) of Vietnamese PNoVs were characterized. Further studies should be developed to get the complete VP1 gene and genome of the Vietnamese PNoV strains to perform genetic analysis deeply.
Conclusions
5
To our knowledge, this was the first study on PNoV infection in north Vietnam. The norovirus genome-positive rate was low (4.9 %), while 25 % of the sampled farms produced positive results in the present study. The five sequences obtained belonged to porcine norovirus GII.19, which has spread widely in pigs. The Vietnamese PNoVs were closely related to the Chinese PNoV strains. This study identified no evidence of any human norovirus among the Vietnamese pigs farmed in north Vietnam.
Funding
The research was supported by the Vietnam National University of Agriculture, Vietnam under grant number T2023-09-36, and the Faculty of Veterinary Medicine, 10.13039/501100004539Kasetsart University, Thailand and by the 10.13039/501100004704National Research Council of Thailand (10.13039/501100004704NRCT) through an 10.13039/501100004704NRCT Senior Scholarship to R. Thanawongnuwech (2022 #N42A650553).
CRediT authorship contribution statement
Hieu Van Dong: Writing – original draft, Software, Methodology, Investigation, Funding acquisition, Formal analysis, Data curation, Conceptualization. Giang Thi Huong Tran: Software, Methodology, Investigation. Amonpun Rattanasrisomporn: Writing – review & editing, Resources. Oumaporn Rungsuriyawiboon: Writing – review & editing. Witsanu Rapichai: Writing – review & editing, Writing – original draft. Jatuporn Rattanasrisomporn: Writing – review & editing, Writing – original draft, Visualization, Supervision, Resources, Project administration, Investigation, Formal analysis, Data curation, Conceptualization.
Declaration of competing interest
The authors declare that there was no conflict of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mead P.S.Slutsker L.Dietz V.Mc Caig L.F.Bresee J.S.Shapiro C.Griffin P.M.Tauxe R.V.Food-related illness and death in the United States Emerg. Infect. Dis.55199960762510.3201/eid 0505.99050210511517 PMC 2627714 · doi ↗ · pubmed ↗
- 2Todd E.C.D.Greig J.D.Viruses of foodborne origin: a review Virus Adapt. Treat.720152545
- 3De Graaf M.Van Beek J.Koopmans M.P.Human norovirus transmission and evolution in a changing world Nat. Rev. Microbiol.147201642143310.1038/nrmicro.2016.4827211790 · doi ↗ · pubmed ↗
- 4Martella V.Lorusso E.Decaro N.Elia G.Radogna A.D'Abramo M.Desario C.Cavalli A.Corrente M.Camero M.Germinario C.A.Banyai K.Di Martino B.Marsilio F.Carmichael L.E.Buonavoglia C.Detection and molecular characterization of a canine norovirus Emerg. Infect. Dis.14820081306130810.3201/eid 1408.08006218680664 PMC 2600395 · doi ↗ · pubmed ↗
- 5Mattison K.Shukla A.Cook A.Pollari F.Friendship R.Kelton D.Bidawid S.Farber J.M.Human noroviruses in swine and cattle Emerg. Infect. Dis.13820071184118810.3201/eid 1308.07000517953089 PMC 2828081 · doi ↗ · pubmed ↗
- 6Martella V.Campolo M.Lorusso E.Cavicchio P.Camero M.Bellacicco A.L.Decaro N.Elia G.Greco G.Corrente M.Desario C.Arista S.Banyai K.Koopmans M.Buonavoglia C.Norovirus in captive lion cub (Panthera leo)Emerg. Infect. Dis.13720071071107310.3201/eid 1307.07026818214183 PMC 2878247 · doi ↗ · pubmed ↗
- 7Chhabra P.de Graaf M.Parra G.I.Chan M.C.Green K.Martella V.Wang Q.White P.A.Katayama K.Vennema H.Koopmans M.P.G.Vinje J.Updated classification of norovirus genogroups and genotypes J. Gen. Virol.1001020191393140610.1099/jgv.0.00131831483239 PMC 7011714 · doi ↗ · pubmed ↗
- 8Jiang X.Wang M.Wang K.Estes M.K.Sequence and genomic organization of Norwalk virus Virology 19511993516110.1006/viro.1993.13458391187 · doi ↗ · pubmed ↗