Ace Deficiency Induces Intestinal Inflammation in Zebrafish
Mingxia Wei, Qinqing Yu, Enguang Li, Yibing Zhao, Chen Sun, Hongyan Li, Zhenhui Liu, Guangdong Ji

TL;DR
This study shows that a deficiency in the ace gene causes intestinal inflammation in zebrafish, offering new insights into inflammatory bowel disease.
Contribution
The study identifies ace as a novel gene involved in intestinal inflammation and IBD pathogenesis in zebrafish.
Findings
Ace deletion in zebrafish increases inflammatory marker gene expression in the intestine.
Ace−/− mutants show higher mucus secretion and are more susceptible to DSS-induced enteritis.
Ace plays a critical role in maintaining intestinal homeostasis.
Abstract
Inflammatory bowel disease (IBD) is a nonspecific chronic inflammatory disease resulting from an immune disorder in the intestine that is prone to relapse and incurable. The understanding of the pathogenesis of IBD remains unclear. In this study, we found that ace (angiotensin-converting enzyme), expressed abundantly in the intestine, plays an important role in IBD. The deletion of ace in zebrafish caused intestinal inflammation with increased expression of the inflammatory marker genes interleukin 1 beta (il1b), matrix metallopeptidase 9 (mmp9), myeloid-specific peroxidase (mpx), leukocyte cell-derived chemotaxin-2-like (lect2l), and chemokine (C-X-C motif) ligand 8b (cxcl8b). Moreover, the secretion of mucus in the ace−/− mutants was significantly higher than that in the wild-type zebrafish, validating the phenotype of intestinal inflammation. This was further confirmed by the IBD…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
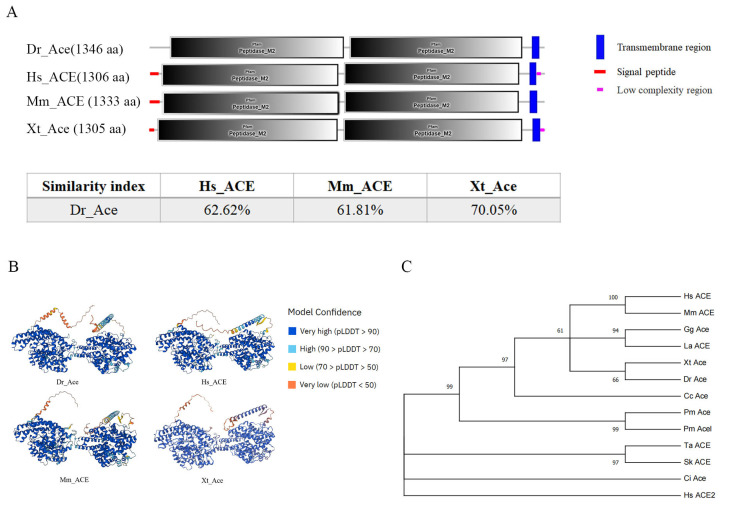 Figure 1
Figure 1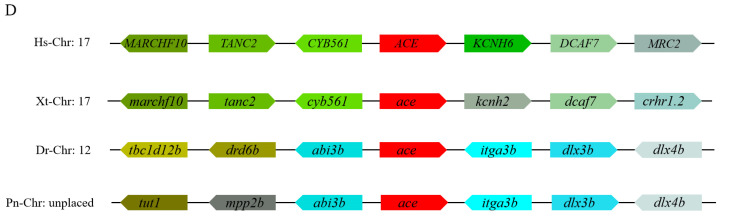 Figure 2
Figure 2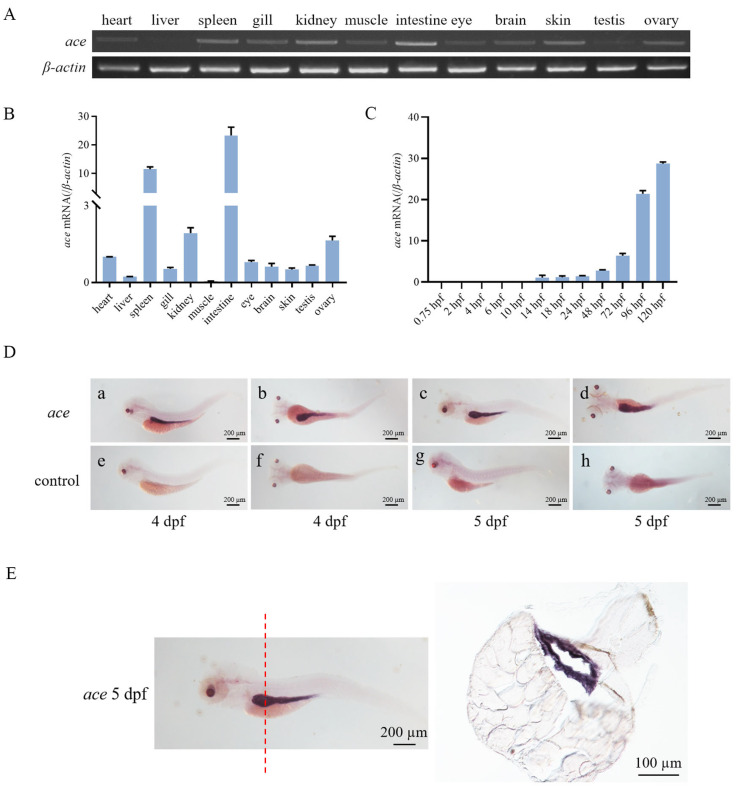 Figure 3
Figure 3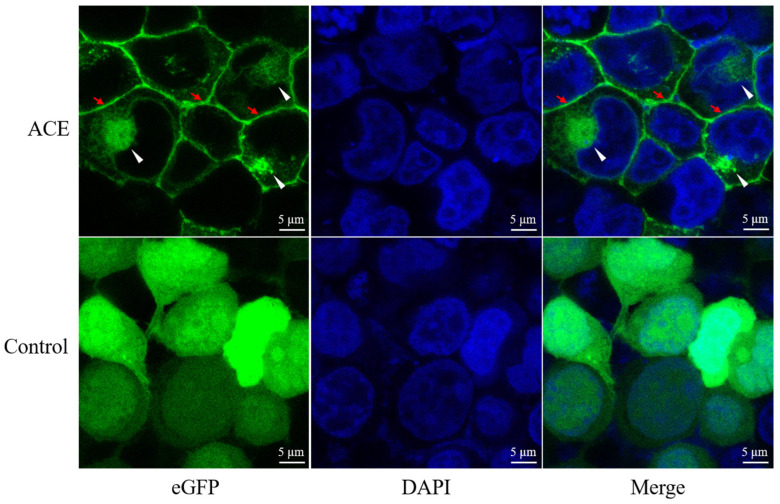 Figure 4
Figure 4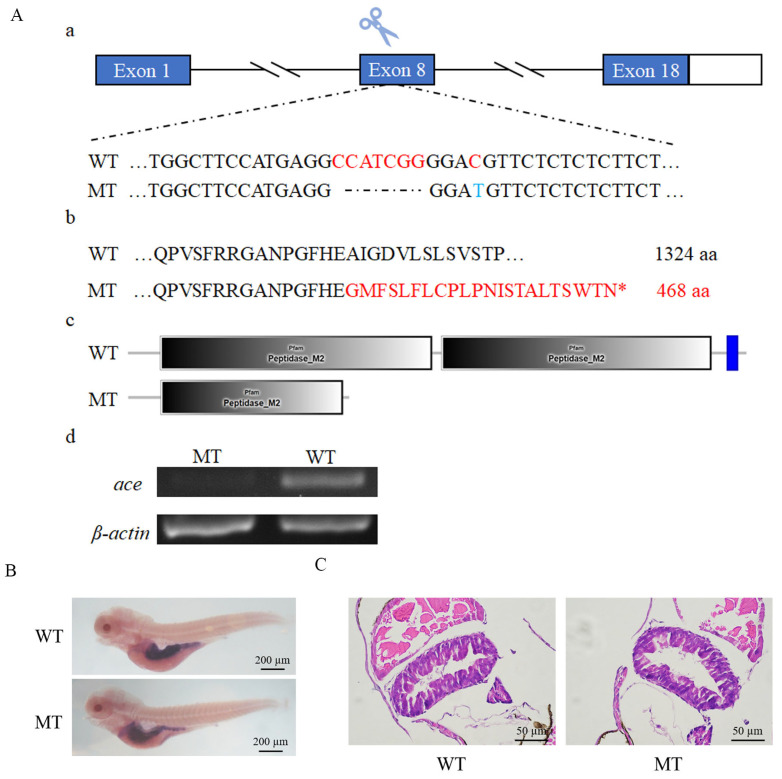 Figure 5
Figure 5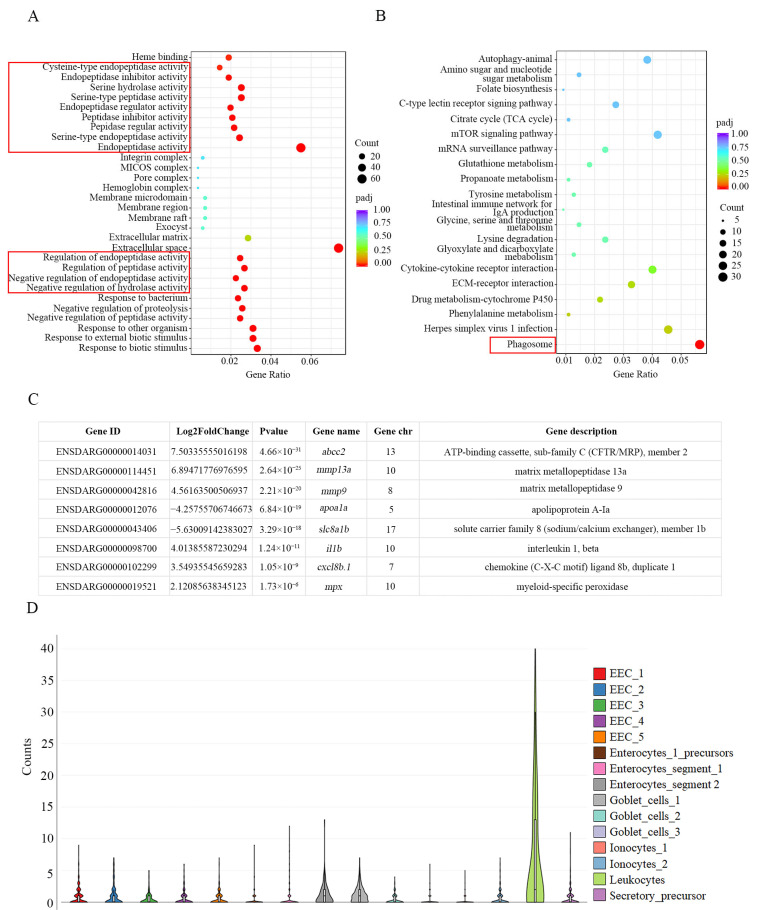 Figure 6
Figure 6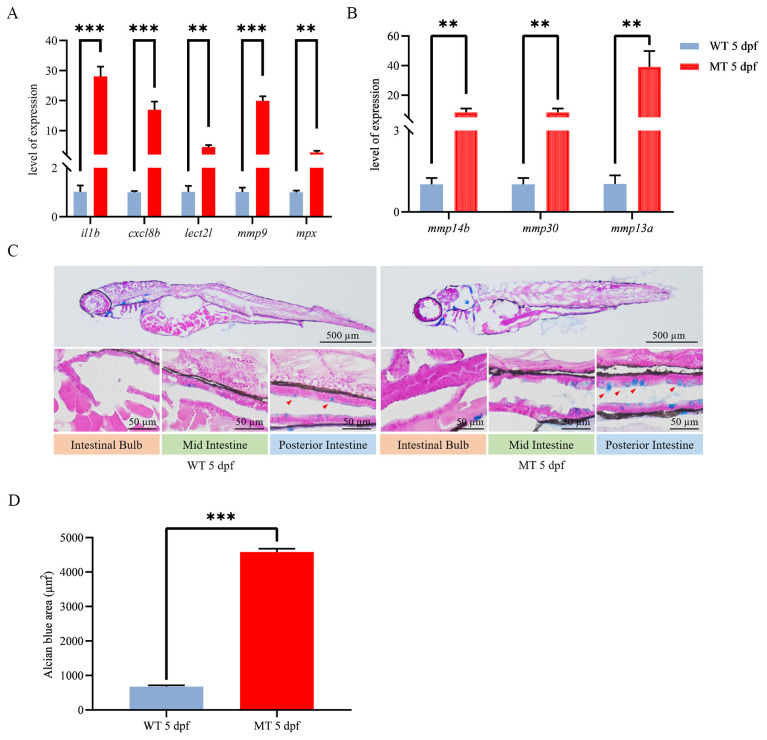 Figure 7
Figure 7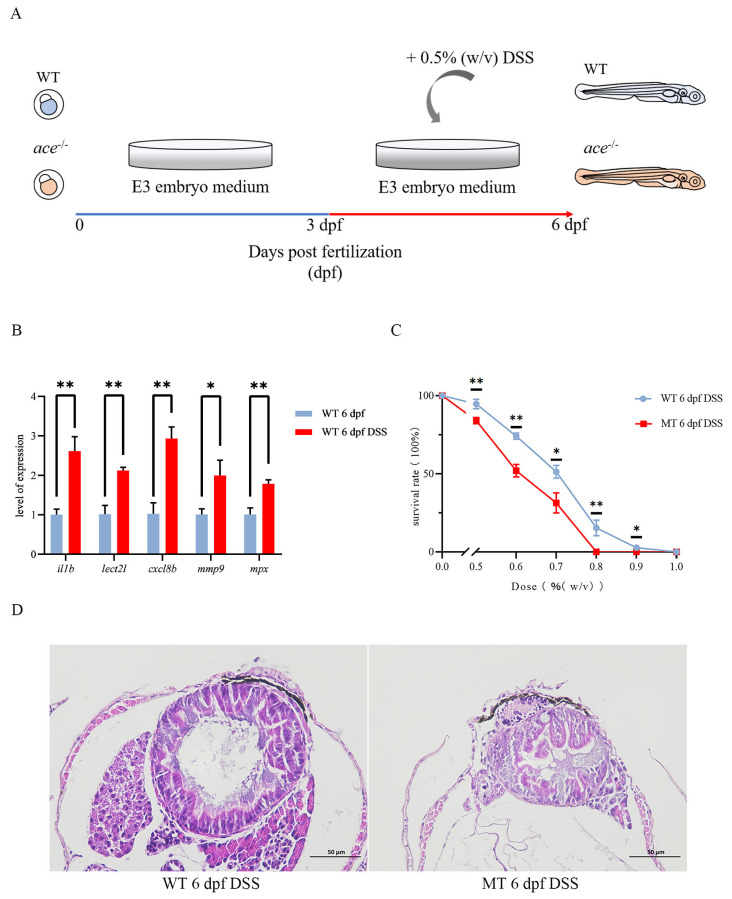 Figure 8
Figure 8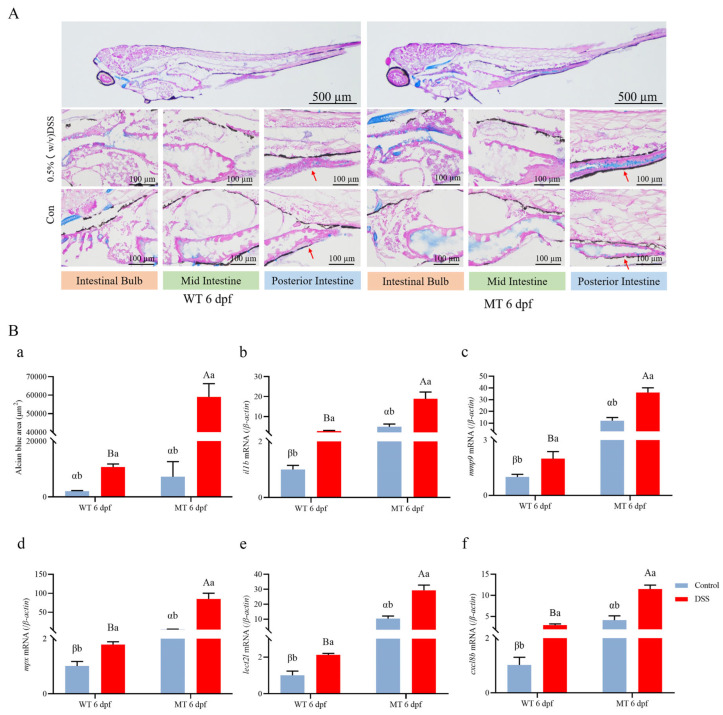 Figure 9
Figure 9- —Natural Science Foundation of China
- —Science & Technology Innovation Project of Laoshan Laboratory
- —National Key Research and Development Project of the Ministry of Science and Technology
- —Shandong Provincial Natural Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEngineering Applied Research
1. Introduction
Chronic intestinal inflammation is a result of a breakdown in intestinal homeostasis. Intestinal inflammation occurs with many gastrointestinal diseases, among which inflammatory bowel disease (IBD) is typical. IBD includes Crohn’s disease (CD) and ulcerative colitis (UC) [1,2]. IBD occurs primarily in the mucosa of the large intestine, leading to debilitating conditions including diarrhea, rectal bleeding, and weight loss [3,4,5]. Animals with IBD have abnormal expressions of proinflammatory molecules such as IL-6, IL-1b, and TNFa and immunoregulatory cytokines such as TGFb, IL-10, and IL-35 [6,7]. Various undetermined environmental and genetic factors, even an inappropriate immune response to gut bacteria, contribute to the disease [8,9,10,11]. However, there is currently no clear pathogenesis of IBD. A widely used model to investigate the pathogenesis of IBD is the dextran sulfate sodium (DSS)-induced colitis model in mice [12,13,14,15].
Zebrafish (Danio rerio) have been used as animal models for human intestinal inflammation diseases including IBD [14,16,17,18]. Many genes that function in inflammatory responses are conserved between zebrafish and mammals. In mammals, for example, NOD1 and NOD2 are both involved in the detection of bacteria and contribute to gastrointestinal inflammation. In zebrafish, Nod1 and Nod2, which are expressed in intestinal epithelial cells and neutrophils, are also IBD-susceptible genes [19,20]. Other genes such as Il-1β and Sst3 also have a similar function in the innate immune response between zebrafish and mammals [21,22,23]. Therefore, zebrafish have proven to be effective models for studying intestinal damage.
The angiotensin-converting enzyme (ACE) is an essential enzyme in the renin–angiotensin–aldosterone system (RAAS), which regulates blood pressure by the cascading of enzyme proteolysis [24]. Two isoforms of mammalian ACE are recognized: the somatic form (sACE) and the germinal form (gACE) [25,26,27,28]. ACE was implicated in the pathological processes of brain ischemic injury, cardiovascular disorders, lung injury, and other processes [29]. In mice lacking ace, manifestations of hypotension, renal vascular thickening, and impaired urine concentration were observed [30,31,32,33]. It was reported that the apoptosis and proliferation of intestinal epithelial cells within the intestinal epithelium are compromised in mice with ACE deficiency [34,35,36]. Thus, potential correlations of ACE with inflammation in the gastrointestinal tract were demonstrated [37,38,39]. Nevertheless, the exact mechanism by which ACE exerts its regulatory effects on the gastrointestinal tract remains unclear.
Based on this, the aim of this study is (1) to understand the evolutionary conservation of zebrafish Ace among vertebrates and its specific expression in the intestine; (2) to investigate the role of Ace in the development and progression of IBD; (3) to analyze the effects of ace deletion in zebrafish on intestinal inflammation; and (4) to confirm the role of Ace in intestinal inflammation using a dextran sodium sulfate (DSS)-induced IBD model.
2. Results
2.1. Ace Is Evolutionarily Conserved in Vertebrates
The zebrafish ace gene was cloned based on the gene information from Ensembl database (Ensembl ID: ENSDARG00000079166). The Ace protein in zebrafish was identified to have two metalloproteinase domains and a transmembrane region, which are conserved among vertebrates (Figure 1A). Also, they have a similar 3-D structure (Figure 1B). The phylogenetic analysis showed that zebrafish Ace proteins are clustered with those from other vertebrates (Figure 1C), and there is syntenic conservation of ace between zebrafish and flameback cichlids; however, no conservation was observed among zebrafish ace genomic neighborhoods and those of humans and xenopus (Figure 1D), suggesting that genomic rearrangements, such as inversions or translocations, may have occurred between ray-finned fishes and tetrapods over evolutionary time, leading to the difference in gene order and synteny.
2.2. Zebrafish ace Expressed in Intestines during Early Development
The relative transcript levels of ace were examined in different tissues of adult zebrafish. We observed that the highest expression of ace was detected in the intestinal tissue of adult zebrafish. During the early stages of development, ace expression became detectable starting from 14 hpf. As the development progressed, the expression of this gene gradually increased (Figure 2C).
To further explore the spatio-temporal expression pattern of ace during development, WISH was performed. The results indicated that the ace gene was specifically expressed in the intestine of zebrafish at 4 dpf and 5 dpf (Figure 2D). This finding was corroborated by sections of the larvae following in situ hybridization (Figure 2E).
2.3. Ace Localized on the Cell Membrane
To investigate the subcellular localization of Ace, HEK293 T cells were transfected with either pcDNA3.1/V5/ace/eGFP or pcDNA3.1/V5/eGFP (control). Following transfection, the cells were stained with DAPI. In comparison to the control, which displayed a uniform distribution of eGFP throughout the cells, Ace exhibited distinct green fluorescence on the cell membrane and endoplasmic reticulum (Figure 3). This observation may be attributed to the transmembrane region of Ace [40].
2.4. Ace Deficiency Does Not Result in Intestine Defects in Zebrafish
To investigate the function of ace, we generated an ace knockout (ace^−/−^) zebrafish by the CRISPR/Cas9 approach. The mutation induced a frameshift in the protein-coding region, leading to premature termination of translation (Figure 4A). As expected, mRNA levels in ace^−/−^ mutants were significantly reduced compared to those in the wild type (Figure 4B). Surprisingly, during development, the ace-deficient zebrafish appeared normal, exhibiting survival to adulthood and fertility without any observable morphological or developmental abnormalities. To explore whether the absence of ace impacts intestinal tube development, we synthesized the intestinal marker fabp2 to probe intestinal development. The WISH analysis did not indicate any noticeable abnormalities, suggesting that the intestinal development program is not evidently affected at 5 dpf (Figure 4B). This was further confirmed by a histological examination (HE staining) of intestinal sections (Figure 4C). Thus, our results suggest that ace deficiency does not significantly alter intestinal development.
2.5. Ace Deficiency Induces Intestinal Inflammation
To investigate the differential expressed genes (DEGs) between wild-type and ace^−/−^ mutant larvae at 5 dpf, an RNA-seq analysis was performed. The results of the Gene Ontology (GO) analysis indicated an upregulation of endopeptidase activity in the absence of ace (Figure 5A). Additionally, Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses revealed an enrichment of DEGs in the phagosome signaling pathway, suggesting a potential involvement in immunity (Figure 5B). Notably, the DEGs also included genes expressed in the intestinal epithelium and associated with inflammation (Figure 5C). Further, ace is mainly expressed in leukocytes in fxr^−/−^ zebrafish larva intestine at 6 dpf [41], which is in line with the RNA-seq results above, indicating that ace plays an important role in intestinal immunity (Figure 5D).
We then assessed the disparity in inflammatory cytokine expression between wild-type and ace^−/−^ zebrafish larvae by real-time PCR. The results clearly demonstrate that the absence of ace substantially augments the expression of interleukin 1 beta (il1b), matrix metallopeptidase 9 (mmp9), leukocyte cell-derived chemotaxin-2-like (lect2l), chemokine (C-X-C motif) ligand 8b (cxcl8b), and the neutrophil marker myeloid-specific peroxidase (mpx) (Figure 6A), as well as some other validated members of the MMP families, such as mmp9, mmp13a, mmp14b, and mmp30 (Figure 6B). The heightened levels of pro-inflammatory cytokines in ace^−/−^ zebrafish indicate the occurrence of an inflammatory response. It is known that goblet cells secrete mucus to safeguard the intestines against infection, primarily concentrated in the mid- and posterior intestine regions. The abundance of intestinal mucus was further evaluated through AB-PAS staining. Interestingly, ace^−/−^ mutants, but not the wild type, exhibited enriched mucus in their digestive tracts at 5 dpf (Figure 6C,D), indicating that goblet cells and mucus could promote intestinal defense and homeostasis. The observation of heightened mucus secretion and elevated expression of pro-inflammatory cytokines in ace-deficient zebrafish suggests that the deletion of ace may contribute to the induction of intestinal inflammation and defense.
2.6. Ace-Deficient Zebrafish Are Susceptible to IBD
To determine whether ace-deficient zebrafish are susceptible to IBD, we first used DSS to construct an intestinal infection model. Three days after drinking DSS, the expression of pro-inflammatory factors, such as il1b, lect21, cxcl8b, mmp9, and mpx, was increased, indicating that the intestinal infection model was successfully generated (Figure 7A,B). When zebrafish larvae were exposed to DSS at a dose between 0.5 and 1% (w/v), the survival rate of ace^−/−^ mutants was lower than that of the wild type (Figure 7C). By HE staining on intestinal sections, we found that the intestinal tract of the ace^−/−^ mutant larvae was significantly smaller, with a thickened wall and a smaller diameter (Figure 7D).
Furthermore, AB-PAS staining showed more mucus in the ace^−/−^ mutants compared to wild-type zebrafish (Figure 8A). Additionally, we compared the expression of inflammatory factors between ace^−/−^ mutants and wild-type larvae treated with DSS. Compared to that in wild-type zebrafish, the expression of il1b, mmp9, lect2l, cxcl8b, and mpx was significantly upregulated in ace-deficient zebrafish (Figure 8B). Collectively, our data indicate that ace-deficient zebrafish are highly susceptible to IBD.
3. Discussion
Angiotensin-converting enzyme (ACE), a zinc-dependent dipeptidyl carboxypeptidase composed of two metalloproteinase domains, plays a vital role in the renin–angiotensin–aldosterone system (RAAS) and is involved in immune regulation [42,43]. In mice, ace deficiency has been reported to affect intestinal epithelial renewal, but its precise function in intestinal inflammation remains unexplored [34]. In this study, we characterized zebrafish ace and found that its deletion induced intestinal inflammation, thereby expanding our understanding of ACE’s functions.
In humans, ACE contains sACE and gACE isoforms. sACE is significantly expressed in various tissues such as the small intestine, duodenum, lungs, kidneys, choroid plexus, and placenta [44,45], while gACE is specifically expressed in testes and is associated with male fertility [46,47]. Notably, the highest expression level of sACE was observed in the small intestine [48]. This is in line with our finding in zebrafish that the expression of the ace gene is highest in the intestine. However, an analysis of tissue expression revealed a lack of notable expression in the testes in zebrafish, and we also found that both female and male ace mutants’ reproduction was not affected when compared to the wild type; this is different from the gACE detected in human, suggesting that there is no functional gACE in zebrafish.
Although the absence of ace appears to have no discernible impact on the zebrafish’s intestinal development as indicated by its morphology, transcripts of the ace gene were detected as early as 14 hpf and continued to be expressed throughout development in different adult tissues including the spleen and intestine. The zebrafish begins to form a digestive tract in a segmental fashion at the mid-somite stages (18 hpf) and completes gut tube morphogenesis at 34 hpf; after the onset of exogenous feeding (5 dpf), there is a dramatic increase in the size of the intestine and the appearance of epithelial cells and other cell types [49]. In accordance with this process, from 4 to 5 dpf, ace specifically localized in the intestine. Moreover, a single-cell transcriptome analysis of zebrafish intestine at 6 dpf showed that ace expressed in many cell types, including the highest expression in leukocytes and moderate expression in enteroendocrine cells (EECs), enterocytes, goblet cells, ionocytes, and secretory precursors [41], suggesting that ace may play an important role in intestinal homeostasis and immunity.
More mucus was secreted in the mid- and posterior intestine of ace^−/−^ mutants compared to the wild type at 5 dpf, indicating the occurrence of intestinal inflammation. Further, the transcriptome sequencing of 5 dpf larvae revealed that the absence of ace had a significant impact on immune pathways, leading to a notable increase in the expression of proinflammatory factors including il1b, lect21, cxcl8b, mpx, and mmp9, as well as several other members of the mmp gene family, such as mmp9, mmp13a, mmp14b, and mmp30. It has been observed that some of the MMP family members are associated with various inflammatory responses [50,51]. Thus, there is a potential correlation between intestinal inflammation and the deletion of ace. This correlation was further supported by the constructed IBD model, where intestinal inflammation induced by DSS showed a similar expression pattern of proinflammatory factors to that observed in ace^−/−^ mutants. Moreover, compared to the wild-type larvae, the ace^−/−^ mutants exhibited a significantly lower survival rate, increased mucus secretion in the intestine, and a notable upregulation of various inflammatory factors in this IBD model, suggesting that both ace deletion and DSS induction synergistically contribute to the occurrence and progression of inflammation. Although ACE in wild-type larvae is mildly upregulated in a dextran sodium sulfate colitis model [52], it is possible that ace deficiency impedes the ability of larvae to mount an appropriate immune response and increases their vulnerability to DSS stimuli, ultimately resulting in earlier mortality compared to the control group.
Many studies have documented the relationship between ACE and inflammation in mammals, and some results seem controversial. For example, ACE overexpression in myeloid-derived cells has been shown to increase the production of pro-inflammatory cytokines, such as IL-12β, TNF, or nitric oxide, while ACE overexpression in neutrophils has been shown to increase resistance to infections with MRSA, Klebsiella pneumoniae, and Pseudomonas aeruginosa [40]. Ace^−/−^ mice have a less vigorous immune response to MRSA infection [40]. However, ACE-overexpressing macrophages have been shown to attenuate neuropathology and neuroinflammation [51]. Although ACE inhibitors have been shown to reduce vascular inflammation, there is no convincing evidence indicating that ACE inhibitors reduce plasma levels of major inflammatory markers in hypertension models [53]. This controversy is partly attributed to the possibility that inflammatory responses may be controlled by local rather than global immune, vascular, and inflammatory cell responses to infection or injury [54], with other localized factors involved in the inflammation process.
In this study, ACE was specifically localized in the intestine and was highly expressed by leukocytes present in intestinal tissue. A deficiency of ACE induced a mild inflammatory response in the intestine of zebrafish, indicating a potential role for ACE in immune regulation and gut health. It appears that other factors also participate in the ace-deficiency-induced inflammatory response, which are likely to depend on immune cells and molecules at the site of tissue damage or infection.
4. Materials and Methods
4.1. Ethics Statement
Embryos were produced through natural mating. All zebrafish studies were conducted according to the Animal Care and Use Committee of the Ocean University of China (SD2007695).
4.2. Zebrafish Strains and Mutants
Zebrafish (D. rerio) from the AB strain were kept at 28 °C and fed twice daily during dark periods of 10 h and 14 h. The fragments of ace were amplified using the specific primers S1 and AS1 by PCR (Table 1). Using CRISPR/Cas9 technology, ace^−/−^ mutant lines were derived from the AB line of zebrafish. This study used 5′-TGGCTTCCATGAGGCCATCG-3′ in exon 8 as the knockout target. Mixtures of Cas9 mRNA and targeting gRNA were microinjected into one-cell-stage zebrafish embryos. A comparison with the wild-type zebrafish sequence confirmed the mutation sites. Fish from the F1 generation carrying a 7 bp deletion were crossed to obtain the F_2_ generation [55,56].
4.3. Bioinformatics Analysis
A set of 12 ACE sequences was primarily collected from National Center for Biotechnology Information (NCBI) website (http://www.ncbi.nlm.nih.gov/). Protein domains were predicted by the SMART website (http://smart.embl.de/); protein 3-D structures were predicted by Alphafold (https://alphafold.com/). A phylogenetic tree was constructed using IQ-tree with the maximum-likelihood algorithm, the L+G4 model, and 1000 bootstrap replications.
4.4. Whole-Mount In Situ Hybridization (WISH) in Zebrafish Larvae
The zebrafish embryos for WISH were cultured from 12 h post-fertilization (hpf) in E3 embryo medium containing 0.004% PTU (Sigema, P6148, Fukushima, Japan). The fragments of ace were amplified using the specific primers S2 and AS2 by PCR (Table 1). The fragments were digested with Sph I (Takara Bio, 1246S, Kusatsu, Japan), and Sp6 RNA polymerase (Thermo Scientific, EP0131) (Waltham, MA, USA) was used to synthesize an antisense probe labelled with digoxigenin (DIG) (Sigema, 11277073910). WISH followed the protocol in its procedures [57]. Stereomicroscopy (Nikon, SMZ1270/1270i, Tokyo, Japan) was used to observe and photograph stained embryos.
4.5. RNA Extraction and RT-qPCR from Zebrafish Samples
Zebrafish total RNA was extracted using Total RNA Kit I (Omega, R6834-02) (Biel/Bienne, Switzerland). DNase was used to treat the RNA, and PrimeScript^TM^ RT reagent with gDNA Eraser (TaKaRa, RP047A) was used to synthesize the cDNA. As controls, reverse transcriptase-free samples were added. Quantitative PCR was performed on an ABI 7500 machine with ChamQ SYBR Color qPCR Master Mix (Vazyme, Q431-02) (Nanjing, China). To normalize the data, the β-actin gene was used as the internal reference gene. A comparative Ct method (2^−ΔΔCt^ method) was used to calculate relative expression levels. All quantitative PCR experiments were conducted in triplicate. This study used the primers shown in Table 1.
4.6. AB-PAS Staining and HE Staining in Zebrafish Larvae
Ice-cold acetone was used to fix zebrafish embryos. Wuhan Servicebio Technology Company performed microtomy and staining of the embryos. In each section (8 μm) after AB-PAS staining, mucus-containing areas of the staining in the mid-intestine and posterior intestine were quantified using ImageJ software (1.48v). The image processing followed the protocol in its procedures [58].
4.7. RNA-Seq Analysis
RNA was extracted from both wild-type and ace^−/−^ larvae at 5 days post-fertilization (dpf). The same batch of samples was sequenced for both control and knockout groups. The transcriptome was sequenced using Novogene (Beijing, China). The gene abundance was calculated and normalized using RPKM (reads per kb per million reads). DEGs (differentially expressed genes) between groups were determined using the EdgeR package (http://www.r-project.org/). Differential gene expression analysis was conducted with a fold change criterion of ≥2 or ≤0.5, and a false discovery rate (FDR) of <0.05 was considered significant. Subsequently, the KEGG pathways (Kyoto Encyclopedia of Genes and Genomes) were analyzed for enrichment. The single-cell profile of ACE in fxr^−/−^ zebrafish larvae at 6 dpf was generated using the Single Cell Portal website (https://singlecell.broadinstitute.org/single_cell/study/SCP1675/zebrafish-intestinal-epithelial-cells-wt-and-fxr?genes=ace&tab=distribution#study-visualize, accessed on 1 December 2023), and the raw data can be accessed from the NCBI GEO Database: GSE173570 [41].
4.8. DSS Treatment
Analyses of DSS (Yeasen Biotech, 60316ES25) effects were conducted on wild-type and ace^−/−^ larvae. All embryos (n = 26–32 larvae per concentration) were cultured to 3 dpf in E3 embryo medium before the tests. From a 10% DSS stock solution (w/v), dilutions were prepared (0.5%, 0.6%, 0.7%, 0.8%, 0.9% w/v, 1.0%) with the medium replaced daily. After 3 days of treatment, the survival rate was calculated. Later experiments (0.5% DSS) used the lowest toxicity dosage. From 3 dpf to 6 dpf, the wild-type and ace^−/−^ DSS groups were cultured in 0.5% DSS in E3 embryo medium, while the corresponding control groups were cultured in parallel in E3 embryo medium only. The RNA was isolated from half of the larvae at 6 dpf, and the other half were used for histopathological analysis [23,59].
4.9. Subcellular Localization
To examine the Ace protein localization in subcellular compartments, a subcellular localization assay was conducted. The pcDNA3.1/V5-His A plasmid was used to create the pcDNA3.1/V5/eGFP vector by cloning eGFP. Next, with primers that contain EcoR I and EcoR V restriction enzyme cutting sites, the complete coding region of ace was amplified by PCR using specific primers (S5, AS5). Recombinant plasmids were constructed by inserting the fragments upstream of pcDNA3.1/V5/eGFP. To investigate the subcellular localization of Ace in HEK293 T cells, the pcDNA3.1/V5/ace/eGFP and pcDNA3.1/V5/eGFP recombinant plasmids were separately transfected. DAPI staining was performed 24 h post-transfection, and observations were made using confocal microscopes after washing the samples with PBS [60].
4.10. Statistical Analysis
Statistical analyses were conducted using Graphpad Prism 9.0.0 software. The data are presented as the mean ± standard deviation. Student’s t-test was used for statistical analysis. p < 0.05 was considered statistically significant.
5. Conclusions
In summary, we demonstrated the evolutionary conservation of zebrafish ace among vertebrates, with specific expression observed in the intestine. Interestingly, deficiency in ace does not lead to intestinal defects in zebrafish but rather triggers intestinal inflammation. Furthermore, zebrafish lacking ace show heightened susceptibility to inflammatory bowel disease (IBD) phenotypes. Future studies are warranted to identify the particular cell types impacted by Ace deficiency, along with the specific signaling pathways in which Ace is involved in innate immunity. This understanding holds significant potential for advancing novel therapeutic strategies for inflammatory bowel diseases in human patients.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bai L. Scott M.K.D. Steinberg E. Kalesinskas L. Habtezion A. Shah N.H. Khatri P. Computational drug repositioning of atorvastatin for ulcerative colitis J. Am. Med. Inf. Assoc.2021282325233510.1093/jamia/ocab 16534529084 PMC 8510297 · doi ↗ · pubmed ↗
- 2Molodecky N.A. Soon I.S. Rabi D.M. Ghali W.A. Ferris M. Chernoff G. Benchimol E.I. Panaccione R. Ghosh S. Barkema H.W. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review Gastroenterology 2012142465410.1053/j.gastro.2011.10.00122001864 · doi ↗ · pubmed ↗
- 3Chachu K.A. Osterman M.T. How to diagnose and treat IBD mimics in the refractory ibd patient who does not have IBD Inflamm. Bowel Dis.2016221262127410.1097/MIB.000000000000072626891261 · doi ↗ · pubmed ↗
- 4Abraham C. Cho J.H. Inflammatory bowel disease N. Engl. J. Med.20093612066207810.1016/j.pop.2023.03.00919923578 PMC 3491806 · doi ↗ · pubmed ↗
- 5Kozlowski C. Jeet S. Beyer J. Guerrero S. Lesch J. Wang X. Devoss J. Diehl L. An entirely automated method to score dss-induced colitis in mice by digital image analysis of pathology slides Dis. Model. Mech.2013685586510.1242/dmm.01175923580198 PMC 3634668 · doi ↗ · pubmed ↗
- 6Levite M. Neuropeptides, by direct interaction with T cells, induce cytokine secretion and break the commitment to a distinct T helper phenotype Proc. Natl. Acad. Sci. USA 199895125441254910.1073/pnas.95.21.125449770522 PMC 22867 · doi ↗ · pubmed ↗
- 7Blum A.M. Metwali A. Mathew R.C. Cook G. Elliott D. Weinstock J.V. Granuloma t lymphocytes in murine schistosomiasis mansoni have somatostatin receptors and respond to somatostatin with decreased ifn-gamma secretion J. Immunol.19921493621362610.4049/jimmunol.149.11.36211358973 · doi ↗ · pubmed ↗
- 8Takagi T. Homma T. Fujii J. Shirasawa N. Yoriki H. Hotta Y. Higashimura Y. Mizushima K. Hirai Y. Katada K. Elevated er stress exacerbates dextran sulfate sodium-induced colitis in prdx 4-knockout mice Free. Radic. Biol. Med.201913415316410.1016/j.freeradbiomed.2018.12.02430578917 · doi ↗ · pubmed ↗