NLRX1 Mediates the Disruption of Intestinal Mucosal Function Caused by Porcine Astrovirus Infection via the Extracellular Regulated Protein Kinases/Myosin Light–Chain Kinase (ERK/MLCK) Pathway
Jie Tao, Jinghua Cheng, Ying Shi, Benqiang Li, Pan Tang, Jiajie Jiao, Huili Liu

TL;DR
This study shows that NLRX1 contributes to intestinal damage caused by porcine astrovirus through the ERK/MLCK pathway, offering insights for antiviral therapies.
Contribution
The study identifies NLRX1's role in PAstV-induced intestinal mucosal disruption via the ERK/MLCK pathway.
Findings
PAstV-4 infection up-regulates NLRX1 and mitophagy in Caco-2 cells.
NLRX1 silencing or 3-MA treatment inhibits PAstV-4 replication and MLC phosphorylation.
ERK and MLCK inhibitors reduce mucosal damage but do not affect NLRX1 expression.
Abstract
Porcine astrovirus (PAstV) has a potential zoonotic risk, with a high proportion of co-infection occurring with porcine epidemic diarrhea virus (PEDV) and other diarrheal pathogens. Despite its high prevalence, the cellular mechanism of PAstV pathogenesis is ill–defined. Previous proteomics analyses have revealed that the differentially expressed protein NOD–like receptor X1 (NLRX1) located in the mitochondria participates in several important antiviral signaling pathways in PAstV–4 infection, which are closely related to mitophagy. In this study, we confirmed that PAstV–4 infection significantly up-regulated NLRX1 and mitophagy in Caco–2 cells, while the silencing of NLRX1 or the treatment of mitophagy inhibitor 3–MA inhibited PAstV–4 replication. Additionally, PAstV–4 infection triggered the activation of the extracellular regulated protein kinases/ myosin light-chain kinase…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
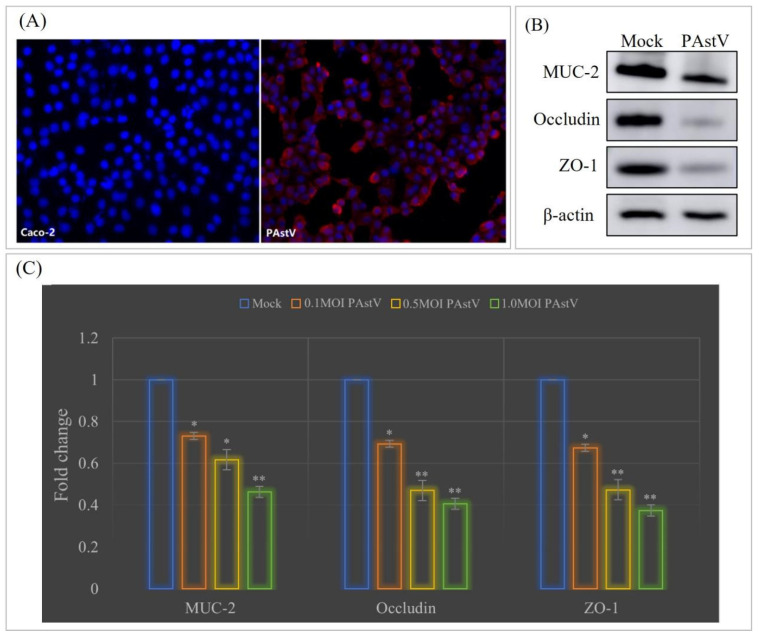 Figure 1
Figure 1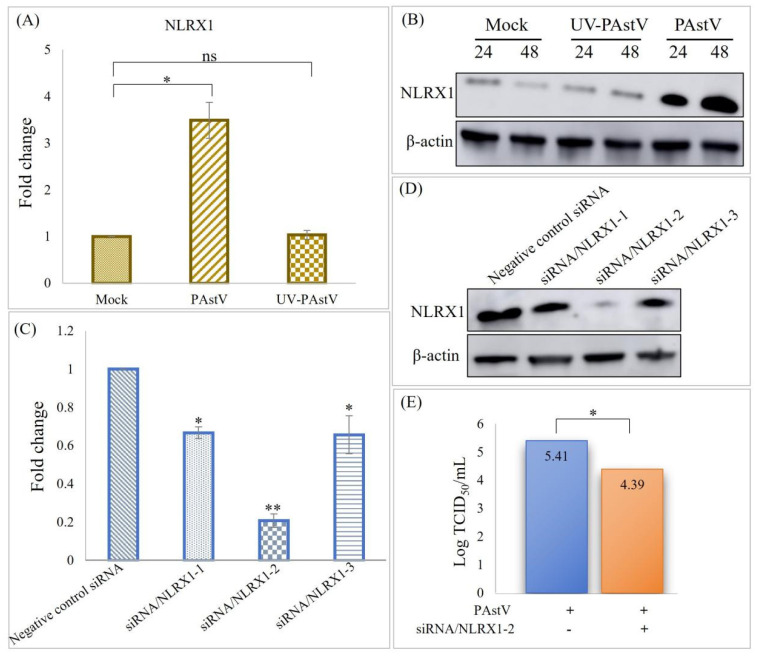 Figure 2
Figure 2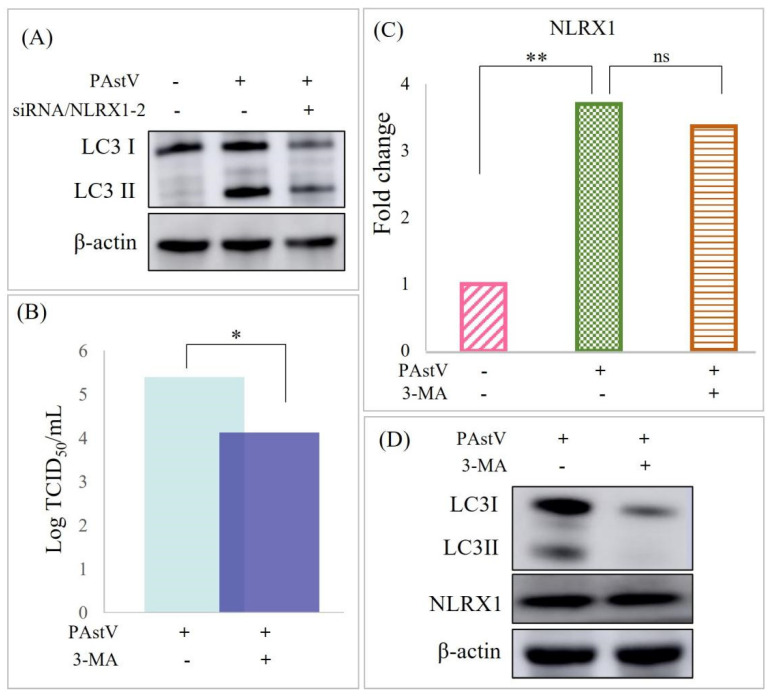 Figure 3
Figure 3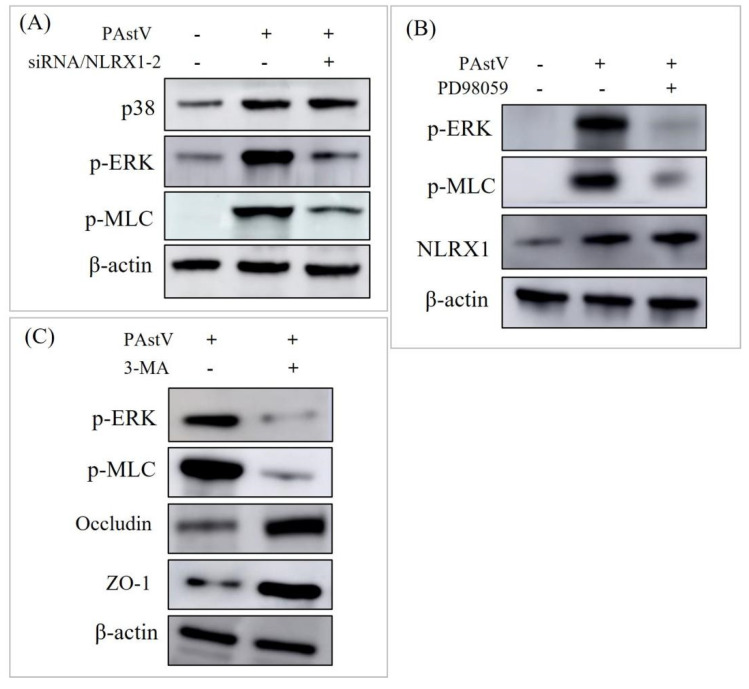 Figure 4
Figure 4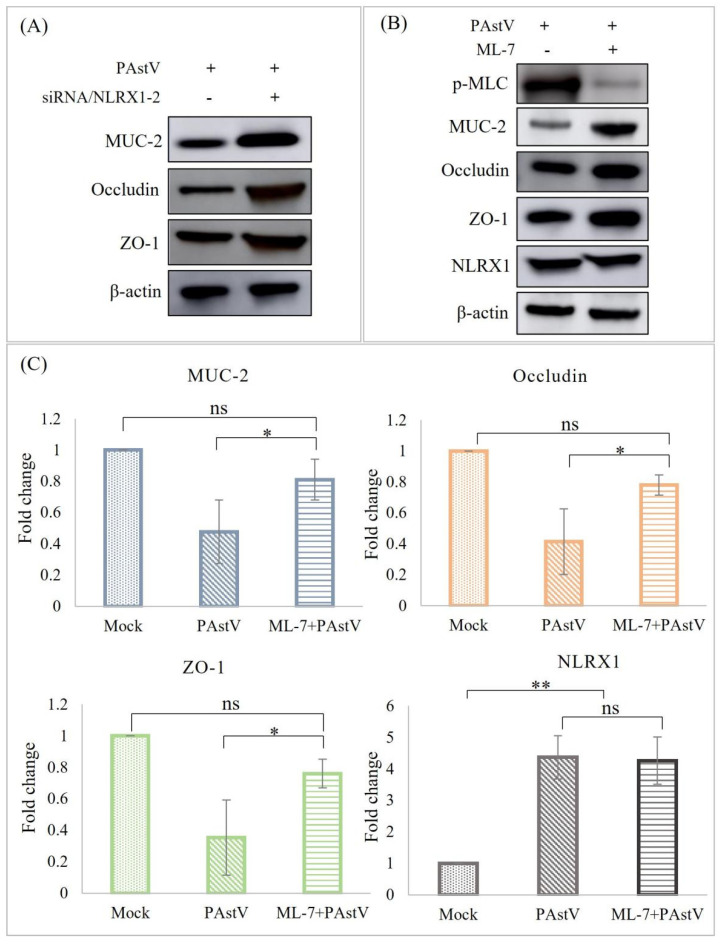 Figure 5
Figure 5- —SAAS Program for Excellent Research Team
- —Shanghai Agricultural Technology Development Program, China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVirus-based gene therapy research · Viral gastroenteritis research and epidemiology · Animal Virus Infections Studies
1. Introduction
Astrovirus (AstV) belongs to the family Astroviridae, including Mamastrovirus (33 species) and Avastrovirus (7 species) [1]. AstV is a potentially zoonotic pathogen that is widely distributed around the world and has been found to infect humans, dogs, cats, pigs, sheep, cattle, mink, deer, bats, dolphins, turkeys, ducks, 31 species of mammals, and 6 species of poultry [2]. This diarrheal disease ranks as the second leading cause of death among children under 5 years of age, accounting for approximately 2 billion cases worldwide annually. In China, the rate of diarrheal disease in children is reported to be 1.9 times per person per year [3]. Human astrovirus (HAstV) is recognized as the second leading cause of gastroenteric diarrhea in humans [4]. Porcine astrovirus (PAstV), which belongs to the Mamastrovirus genus, was first detected by electron microscopy in piglet diarrhea samples in 1980 [5]. However, the first molecular characterization of PAstV was not reported until 2001 [6]. To date, five genotypes of PAstV (PAstV–1 to PAstV–5) with different prevalences have been identified worldwide. In China, the positive rates of PAstV in East China, South China, North China, Central China, Northeast China, and Southwest China are 28.91%, 31.3%, 6.03%, 41.04%, 30.5%, and 32.3%, respectively [7]. PAstV exhibits diverse clinical symptoms due to the presence of different genotypes characterised by significant genetic variations [8]. PAstV–3 can damage the central nervous system (CNS), while the other four genotypes are associated with diarrhea [9,10]. Statistics show that PAstV–2 (81.48%) is the most predominant genotype in China, followed by PAstV–4 (11.11%) and PAstV–5 (7.41%) [11,12]. However, the prevalence ratio of PAstV–4 is highest in European countries (70.4%) and other Asian countries, including Korea (88.46%), Thailand (92.0%), and India (95.65%) [13,14,15]. Additionally, PAstVs frequently coexist with other diarrhea-inducing diseases, such as coronaviruses, caliciviruses, and sapelovirus [12,16,17,18]. It is speculated that PAstV may be an important pathogen involved in the overwhelming majority of co–infection cases. Despite its prevalence, little is known about the mechanism by which astrovirus causes diarrhea. Hence, it is crucial to focus on research regarding the pathogenesis of PAstV, aiming to identify potential antiviral drug targets. This effort could offer new strategies for treating both human and animal gastroenteritis.
The intestinal mucosal barrier serves as the primary defense mechanism against foreign pathogens in the body. When this barrier is compromised, it leads to increased intestinal permeability, disrupting the intestinal barrier [19]. Ultimately, the compromise of the intestinal mucosal barrier initiates an inflammatory response, contributing to the onset of intestinal diseases [20]. It has been reported that granulosomal dysfunction can impair intestinal epithelial cell function integrity, culminating in gastroenteritis disease [21]. It has been reported that AstV only replicates in the intestine, possibly because it needs to change the permeability of the intestinal epithelial cell barrier through capsid proteins, disrupt intestinal tight connections, and expose surface receptors to achieve invasion. The HAstV–1 infection of Caco–2 cells increases epithelial cell permeability by decreasing the occlusion protein and junction complex, and cadherin recombination. But the specific mechanism of PAstV is not clear, and different species of AstVs have great differences in pathogenicity [22].
Our previous research revealed that PAstV–4 infection in PK15 cells induces mitochondrial damage. Among the differential proteins, NLRX1, a member of the Nod-like receptor (NLR) family, is located in the mitochondria and is highly expressed in the intestine, and is known to modulate ROS production, mitochondrial damage, autophagy, and apoptosis [23]. This might suggest a potentially significant role for NLRX1 in the context of PAstV–4 infection. Hence, in this study, we examined the impact of NLRX1 on PAstV–4 replication and further investigated its regulatory role in disrupting intestinal mucosal barrier function induced by PAstV–4 infection. Our findings indicated that NLRX1 could enhance the replication of PAstV–4, and the silencing of NLRX1 alleviated the down-regulation of tight-junction protein in intestinal epithelial cells through the ERK/MLCK pathway. In conclusion, the data in this study revealed that NLRX1 plays a vital role in destruction of the intestinal mucosal barrier induced by PAstV–4 infection via mitophagy. Further, we explores whether this phenomenon is consistent in four other genotypes.
2. Materials and Methods
2.1. Cells, Viruses, and Antibodies
The human colon adenocarcinoma cell line Caco–2, provided by Prof. Guoqiang Zhu from Yang Zhou University, was cultured in Dulbecco’s modified Eagle’s medium (Gibco, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (Gibco, Carlsbad, CA, USA) within a humidified atmosphere containing 5% CO_2_ at 37 °C. The porcine astrovirus type 4 stain PAstV/SH/2022/CM1, identified in our laboratory, was propagated in Caco-2 cells with 15 μg/mL Pancreatin. The viral titer was determined by measuring the 50% tissue culture infective dose (TCID_50_). The mouse monoclonal antibody against PAstV was obtained from LanDu Biotechnology (Binzhou, China). Additionally, the anti–NLRX1 polyclonal antibody was received from Proteintech (Wuhan, China), and the anti–HA polyclonal antibody, anti–β–action antibody (Abcam, Cambridge, UK), HRP-conjugated anti-mouse IgG, and FITC–conjugated anti–mouse IgG (Abcam, Cambridge, UK) were utilized following the instructions of the manufacturers.
2.2. Transfection and Silencing of the NLRX1 Gene Using siRNA
For siRNA knockdown experiments, Caco–2 cells were seeded in 6-well plates and transfected twice with 25 nM of the designated siRNAs over 48 h using Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA). The siRNA employed were as follows: siRNA/NLRX1–1, 5′–UUGUCAAUCUGCUGCGCAA–3′; siRNA/NLRX1–2, 5′–GUGCUGGGCUUGCGGAAGA–3′; siRNA/NLRX1–3, 5′–GCAUGUCUUCCGCCGGGAU–3′; negative control siRNA, 5′–UUCUCCGAACGUGUCACGUTT–3′.
2.3. TCID50 for PAstV
PAstV titers were assessed through an indirect fluorescence assay using the Reed–Muench method. Caco–2 cells were seeded in 96–well plates, and eight replicates of serial 10–fold dilutions of PAstV were inoculated. After 96 h, an indirect fluorescence assay was conducted to calculate the TCID_50_ using the PAstV specific antibody [24].
2.4. Quantitative Real-Time PCR (qPCR)
At the specified time points, total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Subsequently, reverse transcription was carried out utilizing a PrimeScript^®^ RT reagent Kit with a gDNA Eraser (Takara Bio, Otsu, Japan). Following reverse transcription, quantitative PCR (qPCR) was conducted using SYBR Green Real–Time PCR Master Mix (Takara Bio, Otsu, Japan) and an ABI7500 system (ABI, Madison, WI, USA). The amplification conditions included initial denaturation at 95 °C for 30 s, followed by 40 cycles of denaturation at 95 °C for 5 s, annealing at 60 °C for 30 s, and extension at 95 °C for 15 s. The β–actin gene served as an internal standard, and the results were calculated utilizing the 2^−ΔΔCt^ method [25]. The primer sequences employed for amplification were as follows: occludin (F: 5′–TCCAACGGGAAAGTGAACGA–3′; R: 5–GTGGATATTCCCTGATCCAGTCTT–3′), ZO-1 (F: 5′–AAGGTAAAGTCTGCTGAGGCTGAA–3′; R: 5′–GACACTGAATTACCTTCGCCG-3′), MUC–2 (F: 5′–GTCGAGTACATCCTGCTGACG–3′; R: 5′–GAGTCCTCTCTGTTTCCACACG–3′), NLRX1(F: 5′–CAGACCCTCACAAGCATCTA–3’; R: 5′–CACGGACATCCTCTTCAGA–3′), β-actin (F: 5′–TGGGTCAGAAGGACTCCTATG–3′; R: 5′–CAGGCAGCTCATAGCTCTTCT–3′).
2.5. Western Blotting Analysis
Cells cultured in 60 mm dishes were prepared by adding 200 μL of 2 × lysis buffer A (65 mM Tris–HCL, 4% SDS, 3% DL–dithiothreitol, and 40% glycerol) and incubated for 30 min on ice. Subsequently, the samples were mixed with 5×SDS buffer and boiled for 10 min. The proteins were separated by 12% SDS–PAGE and electroblotted onto a polyvinylidene difluoride (PVDF) membrane (Millipore, Billerica, MA, USA) using eBlot L1 (GenScriptTM, Nanjing, China). After blocking for 1 h at room temperature with Tris–buffered saline containing 5% skim milk, the membranes were incubated overnight at 4 °C with the specified primary antibodies. This was followed by incubation with HRP–conjugated secondary antibodies at room temperature for 1 h. Finally, the proteins were visualized via staining using enhanced chemiluminescence detection kits (Thermo Fisher Scientific, Waltham, MA, USA).
2.6. Statistical Analysis
The values are presented as mean ± standard deviation (SD). Data were analyzed using Student’s t-test and processed using SPSS. p values less than 0.05 were considered statistically significant.
3. Results
3.1. Down-Regulation of Tight-Junction Proteins in Caco–2 Cells Following PAstV–4 Infection
To investigate the impact of PAstV infection on the mucosal barrier function of intestinal epithelial cells, the susceptibility of PAstV–4 to human colon cancer epithelial cells (Caco–2) was initially assessed. The results confirmed the successful proliferation of the PAstV/SH/2022/CM1 strain on Caco–2 cells, with a viable titer of 10^5.23^TCID_50_/0.1 mL (Figure 1A). Subsequently, the effect of PAstV infection on epithelial mucosal barrier function was assessed. The results demonstrated down-regulation in the transcription and expression of the MUC–2 protein and the tight-junction proteins of occludin and ZO–1 upon PAstV–4 infection (Figure 1B). Furthermore, the degree of inhibition increased with an increasing infection dose of PAstV–4 (Figure 1C).
3.2. Up-Regulation of NLRX1 Expression Induced by PAstV Infection
Building upon our prior findings indicating that PAstV/SH/2022/CM1 infection up-regulated the expression of NLRX1 in PK15 cells, we sought to confirm this result in the intestinal epithelial cells during PAstV infection. To achieve this, caco−2 cells were infected with PAstV−4 at an MOI of 1. Real-time PCR analysis confirmed the significant up-regulation of NLRX1 expression at 24 h post-infection (Figure 2A). Notably, UV−inactivated PAstV−4 failed to induce NLRX1 mRNA expression, suggesting that the up-regulation of NLRX1 depends on viral replication (Figure 2A). Furthermore, the results revealed that PAstV−4 up–regulated NLRX1 dose-dependently (Figure 2B).
3.3. PAstV–4 Replication Augmented by NLRX1 Knockdown in Caco−2 Cells
To investigate the impact of the endogenous NLRX1 on PAstV−4 replication, three pairs of siRNA duplexes targeting NRLX1 were separately transfected into Caco-2 cells. The knockdown efficiency of these siRNAs was assessed through real-time PCR, revealing that siRNA/NLRX1−2 achieved the highest efficiency (Figure 2C). Western blot analysis confirmed that siRNA/NLRX1−2 significantly reduced the level of NLRX1 protein by approximately 89% compared to control cells transfected with NC siRNA (Figure 2D). Subsequently, siRNA/NLRX1−2 was transfected into Caco−2 cells, followed by PAstV–4 infection, which decreased PAstV–4 replication within Caco−2 cells (Figure 2E). This suggests that the endogenous NLRX1 can potentially promote the replication of PAstV–4 in Caco−2 cells. PAstV−4 infection also resulted in the up-regulation of LC3II proteins, and this effect was inhibited by treatment with siRNA/NLRX1 (Figure 3A). Additionally, treatment with 20 μM 3−MA (mitophagy inhibitor) decreased the viral titer of PAstV (Figure 3B) while not affecting NRLX1 expression (Figure 3C,D). We speculated that PAstV–4 infection activated mitochondrial autophagy to resist host innate immunity through the up-regulation of NLRX1, thereby promoting its replication.
3.4. Disruption of Caco−2 Mucosal Barrier through ERK/MLC Pathway Activation by NLRX1
The ERK/MLC pathway is crucially associated with intestinal mucosal barrier function. Following infection with PAstV−4 (1.0 MOI) for 24 h, increased p38, p−ERK, and p−MLC levels were observed through Western blot analysis. Moreover, the knockdown of NLRX1 hindered the expression of p−ERK and p−MLC induced by PAstV−4 infection (Figure 4A). Moreover, the treatment with 20 μM PD98059 (ERK inhibitor) decreased the expression of p−MLC. In contrast, NLRX1 expression remained unaffected (Figure 4B). Subsequently, the impact of mitophagy on the ERK/MLCK pathway was assessed. The results indicated that treatment with 3−MA down-regulates ERK and MLC phosphorylation and up-regulated the expression of occludin and ZO−1. This suggests that NLRX1 is implicated in regulating the activation of the ERK/MLCK pathway through mitophagy.
To investigate the role of NLRX1 in regulating intestinal mucosal barrier function, siRNA/NLRX1−2 was transfected into Caco−2 cells, followed by PAstV−4 infection. Subsequently, the expression levels of MUC−2, occludin, and ZO−1 were assessed using Western blot analysis. The results demonstrated that the down-regulation of the three proteins induced by PAstV−4 infection was alleviated (Figure 5A). Furthermore, treatment with 25 μM ML−7 (MLCK inhibitor) exhibited this phenomenon while not affecting NLRX1 expression (Figure 5B,C). MLCK is responsible for the phosphorylation of MLC. This suggests that NRLX1 might regulate mucosal barrier function through the ERK/MLCK pathway during PAstV−4 infection.
4. Discussion
AstV is recognized as a potential zoonotic pathogen, with reported infections in thirty−one species of mammals and six poultry species, leading to symptoms such as diarrhea and neurological issues [26]. In particular, PAstV primarily induces symptoms like diarrhea, vomiting, and anorexia in piglets. Diarrheal pig feces are characterised by a mushy consistency or yellow water, with pigs exhibiting clinical symptoms similar to those caused by the porcine epidemic diarrhea virus (PEDV) [12,27]. Furthermore, synergistic effects with PEDV may exist, emphasizing the need for careful attention to such co−infections. It is necessary to first study the mechanism of PAstV infection before studying its synergistic pathogenesis. Our previous study discovered that the NLRX1 protein, predominantly in mitochondria, exhibited significant up−regulation following PAstV−4 infection. This up−regulation was associated with the mitochondrial autophagy pathway, indicating a potentially crucial role for NLRX1 in the context of PAstV infection. Hence, this study aimed to improve our understanding of the impact and regulation of the NLRX1 protein on PAstV infection.
NLRX1 is a modulator of pathogen-associated molecular pattern receptors and identifies a key therapeutic target for enhancing antiviral responses [28]. Li et al. found that NLRX1 regulated mitophagy via the FUNDC1−NIPSNAP1/NIPSNAP2 signaling pathway in intestinal ischemic reperfusion (IR) injury [29]. Based on the property of regulating mitophagy, NLRX1 was found to facilitate the replication of human immunodeficiency virus 1 (HIV−1), herpes simplex virus 1 (HSV−1), and hepatitis C virus (HCV), whereas it restricts influenza A virus (IAV), hepatitis A virus (HAV), and porcine reproductive and respiratory syndrome virus (PRRSV) replication [30]. Until now, the specific role of NLRX1 in AstVs infection was scarcely known. In this study, we found that PAstV−4 infection up−regulated the expression of NLRX1 and LC3II, while silencing the expression of NLRX1 inhibited PAstV replication and mitophagy. It was speculated that PAstV−4 activated mitophagy to resist host innate immunity through the up-regulation of NLRX1, thereby promoting viral replication. Several studies have confirmed the close relationship between intestinal barrier injury and mitophagy [31,32]. However, there are no report about NLRX1 being involved in this. This study first explored the role of NLRX1 on intestinal mucosal barrier injury induced by PAstV−4 infection.
In astrovirus infection, destruction of the intestinal epithelium and the stimulation of an inflammatory response are not the main mechanisms that cause diarrhea [33]. Astrovirus increased barrier permeability in a Caco−2 cells, correlated with disruption of the tight-junction protein occludin, and decreased the number of actin stress fibers [34]. Indeed, the impairment of tight junctions and adhesive junction structures has been established as a contributing factor in the pathogenesis of PEDV and various other pathogens [35,36,37]. Oral administration of the turkey astrovirus 2 (TAstV−2) structural (capsid) protein induced acute diarrhea, increasing barrier permeability [38]. HAstV infection increased the permeability of the endothelial cell barrier through the activation of MLC phosphorylation [39]. However, the effect of PAstV on the intestinal mucosal barrier remains unknown. Fang et al. reported that seven−day−old suckling piglets being infected with PAstV-GX1 (PAstV−1) led to mild diarrhea, growth retardation, and damage to the intestinal mucosal villi [40]. They speculated that PAstV infection has a great influence on the intestinal mucosal injury. The results of this study revealed that PAstV−4 infection activated ERK and MLC phosphorylation, followed by the down-regulation of MUC−2, occludin, and ZO−1 proteins. Moreover, the knockdown of NLRX1 hindered the activation of p−ERK and p−MLC and then alleviated intestinal mucosal barrier injury. We therefore hypothesized that endogenous NLRX1 might enhance PAstV−4 replication and contribute to intestinal mucosal barrier function impairment via the ERK/MLCK pathway. ERK/MLCK is closely related to the regulation of intestinal barrier function [41,42]. In this study, we present the first report of NLRX1 playing a key role in regulating intestinal barrier injury through the ERK/MLCK pathway. However, whether this regulation is direct or indirect still needs to be further elucidated. The current research on NLRX1 mainly focuses on its effect on viral replication and mitophagy. It has been demonstrated that the interaction between NLRX1 and viral components might be a key factor in determining the outcome of viral infections [30,43]. In view of this, we guess that there is an intermedium between NLRX1 and the ERK/MLCK pathway, which will be further explored. The intermedium may be a certain viral protein of PAstV−4 or a host protein in Caco−2 cells. Subsequently, screening and identification of the interacting proteins of NLRX1 will be carried out so as to resolve the specific mechanism of NLRX1 in the dysfunction of the intestinal mucosal barrier induced by PAstV−4 infection.
There are five PAstV genotypes prevalent in the world with different clinical characteristics. Except PAstV−3, diarrhea is the main feature of all of the PAstV genotypes. PAstV−4 is the most prevalent genotype worldwide. So, the data in this study can provide valuable clues for PAstV pathogenesis. However, the regulatory effects of NLRX1 on the other PAstV genotypes also need to be explored.
In summary, this study first proposed that PAstV−4 infection impaired the mucosal barrier function of Caco−2 cells by activating the ERK/MLCK pathway, and NLRX1 played a vital regulatory role in this network. These findings contribute to a better understanding of the clinical pathogenesis of PAstV infection and provide a foundation for future research on antiviral drug development. Importantly, exploring the mechanisms of intestinal mucosal barrier injury caused by different porcine diarrhea viruses contributes to finding the most common target molecules, so as to develop common antiviral drugs for the prevention and control of porcine diarrhea disease.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rawal G. Linhares D.C.L. Scoping review on the epidemiology, diagnostics and clinical significance of porcine astroviruses Transbound. Emerg. Dis.20226997498510.1111/tbed.1412333900029 · doi ↗ · pubmed ↗
- 2Nguyen T.V. Piewbang C. Techangamsuwan S. Genetic characterization of canine astrovirus in non-diarrhea dogs and diarrhea dogs in Vietnam and Thailand reveals the presence of a unique lineage Front. Vet. Sci.202310127841710.3389/fvets.2023.127841737818392 PMC 10561284 · doi ↗ · pubmed ↗
- 3Chen J. Wan C.M. Gong S.T. Fang F. Sun M. Qian Y. Huang Y. Wang B.X. Xu C.D. Ye L.Y. Chinese clinical practice guidelines for acute infectious diarrhea in children World J. Pediatr.20181442943610.1007/s 12519-018-0190-230269306 · doi ↗ · pubmed ↗
- 4Vu D.L. Bosch A. Pinto R.M. Guix S. Epidemiology of Classic and Novel Human Astrovirus: Gastroenteritis and Beyond Viruses 201793310.3390/v 902003328218712 PMC 5332952 · doi ↗ · pubmed ↗
- 5Bridger J.C. Detection by electron microscopy of caliciviruses, astroviruses and rotavirus–like particles in the faeces of piglets with diarrhoea Vet. Rec.19801075325336258286 · pubmed ↗
- 6Jonassen C.M. Jonassen T.O. Saif Y.M. Snodgrass D.R. Ushijima H. Shimizu M. Grinde B. Comparison of capsid sequences from human and animal astroviruses J. Gen. Virol.200182 Pt 51061106710.1099/0022-1317-82-5-106111297680 · doi ↗ · pubmed ↗
- 7Zhou Y. Wang Y. Sun J. Su M. Song H. Research progress of porcine astrovirus (P Ast V) epidemic in China Chinese J. Pre. Vet. Med.20244616
- 8Zhao C. Chen C. Li Y. Dong S. Tan K. Tian Y. Zhang L. Huang J. Zhang L. Genomic characterization of a novel recombinant porcine astrovirus isolated in northeastern China Arch. Virol.20191641469147310.1007/s 00705-019-04162-830868264 · doi ↗ · pubmed ↗