An Optimized Protocol for the Generation of Alveolospheres from Wild-Type Mice
Mahsa Zabihi, Ali Khadim, Theresa M. Schäfer, Ioannis Alexopoulos, Marek Bartkuhn, Elie El Agha, Ana I. Vazquez-Armendariz, Susanne Herold

TL;DR
This paper presents an improved method to generate alveolospheres from wild-type mice, eliminating the need for transgenic models and ensuring purity.
Contribution
The novel contribution is a refined protocol that enables pure alveolosphere generation without transgenic mice.
Findings
The optimized protocol eliminates the need for transgenic mice to isolate alveolar epithelial cells.
The method produces pure alveolospheres without contamination from bronchiolar or bronchioalveolar organoids.
This approach enhances the standardization of alveolosphere generation for lung research.
Abstract
Organoid models have become an integral part of the research methodology in the lung field. These systems allow for the study of progenitor and stem cell self-renewal, self-organization, and differentiation. Distinct models of lung organoids mimicking various anatomical regions of mature lungs have emerged in parallel to the increased gain of knowledge regarding epithelial stem and progenitor cell populations and the corresponding mesenchymal cells that populate the in vivo niche. In the distal lung, type 2 alveolar epithelial cells (AEC2s) represent a stem cell population that is engaged in regenerative mechanisms in response to various insults. These cells self-renew and give rise to AEC1s that carry out gas exchange. Multiple experimental protocols allowing the generation of alveolar organoids, or alveolospheres, from murine lungs have been described. Among the drawbacks have been…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
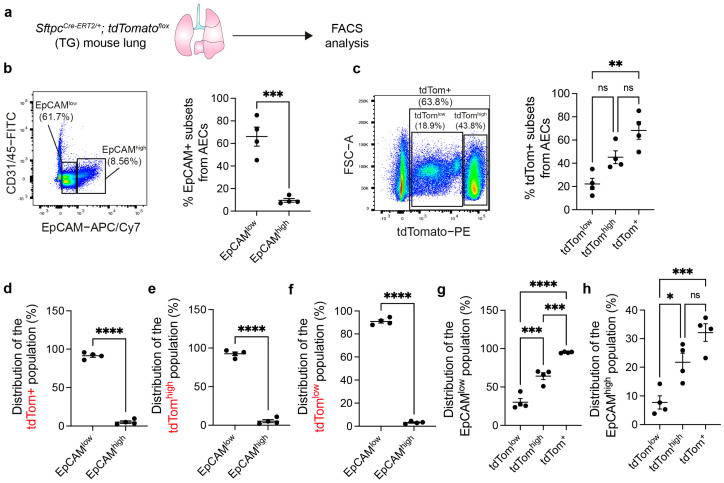 Figure 1
Figure 1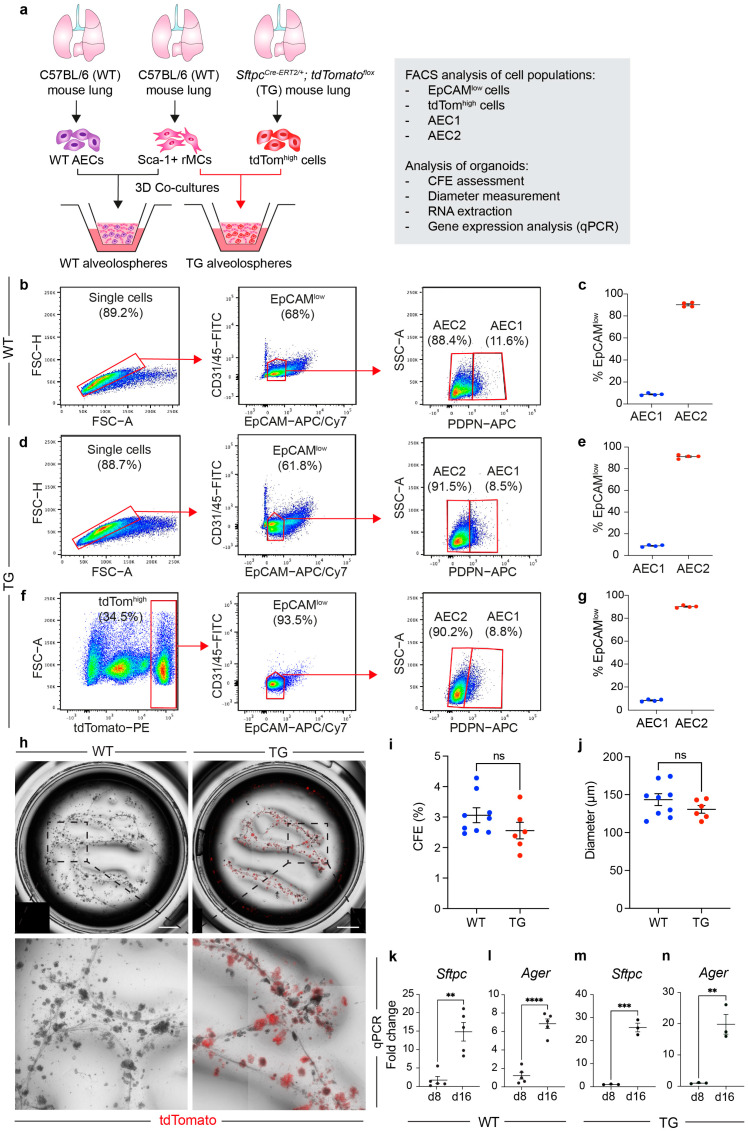 Figure 2
Figure 2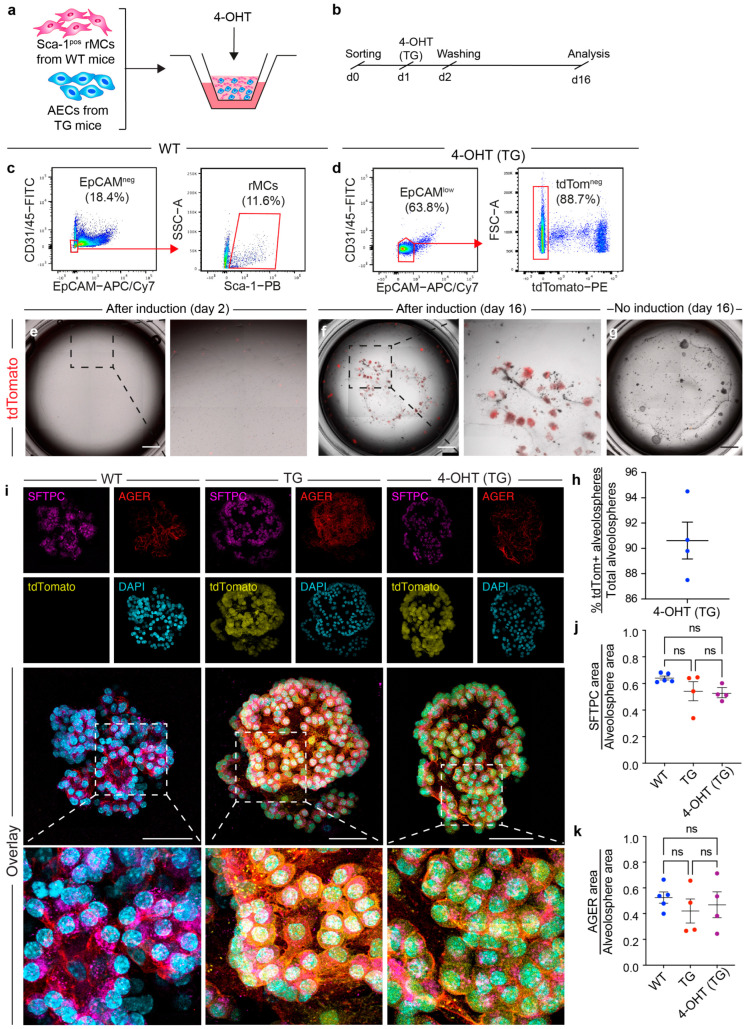 Figure 3
Figure 3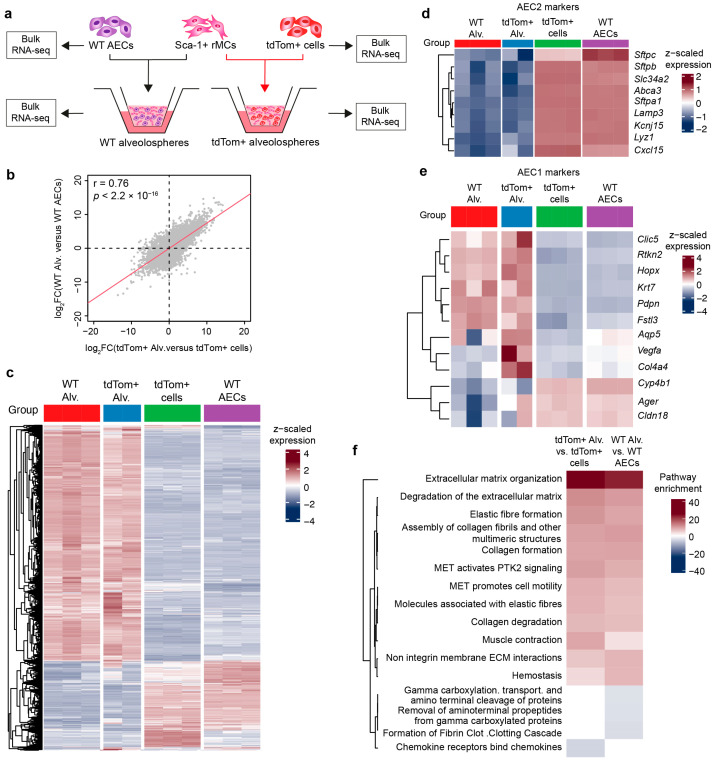 Figure 4
Figure 4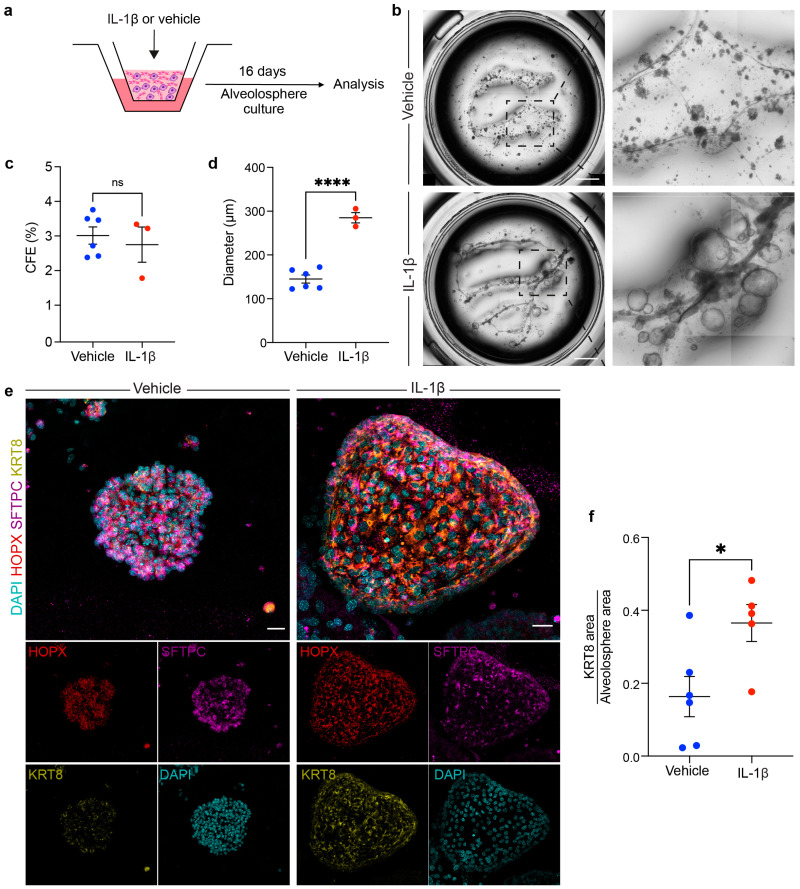 Figure 5
Figure 5- —German Research Foundation
- —Institute for Lung Health (ILH), Excellence Cluster Cardio-Pulmonary Institute
- —Transdisciplinary Research Area (TRA) “Life and Health”, the Life & Medical Science Institute (LIMES), the excellence cluster ImmunoSensation
- —Hessen State Ministry of Higher Education, Research and the Arts
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeonatal Respiratory Health Research · Congenital Diaphragmatic Hernia Studies · Medical Imaging and Pathology Studies
1. Introduction
Organoids are three-dimensional (3D) structures that are representative of the structure, cellular morphology, and certain functional features of the tissue or organ of interest. In recent years, murine and human organoids have emerged as powerful tools for studying organ development, cellular interaction, drug screening, and regenerative mechanisms. Due to their unlimited capacity to proliferate and differentiate, adult stem and progenitor cells have been widely used to develop organoids for research purposes [1,2].
In the context of pulmonary research, several murine organoid systems have been developed to create airway and/or alveolar structures resembling various aspects of lung organogenesis, homeostasis, and regeneration [3]. For instance, tracheal organoids, or tracheospheres, with a visible lumen can be grown from basal stem cells even in the absence of stromal cells or other non-basal cells [4]. Airway organoids, or bronchiolospheres, can be generated by co-culturing club cells with airway smooth muscle cells (ASMCs) or repair-supportive mesenchymal cells (RSMCs), thus mimicking the intercellular communication that occurs during airway epithelial regeneration in vivo following naphthalene injury [5,6,7,8]. Such bronchiolospheres can also be generated by culturing club cells with [CD45− CD31− EpCAM− Sca-1+] (non-leukocytic, non-endothelial, non-epithelial, stem cell antigen-1-expressing) resident mesenchymal cells (rMCs) [5]. The most comprehensive system so far is the bronchioalveolar lung organoid (BALO) model, which relies on co-culturing bronchioalveolar stem cells (BASCs) and rMCs, resulting in a highly sophisticated mimicry of the bronchioalveolar region of the mature mouse lung [7,9].
Type 2 alveolar epithelial cells (AEC2s) represent a stem cell population in the alveolar compartment of the lung [10]. These cells self-renew and regenerate this compartment by differentiating into type 1 alveolar epithelial cells (AEC1s) that are responsible for gas exchange, eventually via intermediate cell types [11,12,13]. Although the heterogeneous nature of AEC2s was not obvious initially, emerging research suggests that there are indeed AEC2 subsets that display varying stem-cell features. For example, WNT-responsive AEC2s (expressing axis inhibition protein 2 or AXIN2), termed alveolar epithelial progenitors (AEPs), have been shown to represent a stable lineage during homeostasis, and are capable of rapid expansion following acute lung injury [14]. Another subpopulation of AEC2s is enriched in programmed cell death 1 ligand 1 (PD-L1 or CD274) expression and was dubbed “injury-associated alveolar progenitors” (IAAPs). These cells replenish the mature AEC2 pool upon alveolar injury [15,16].
Isolation of AEC2s by FACS for alveolosphere cultures has so far mostly relied on the use of genetically modified mice (Sftpc^Cre-ERT2/+^; tdTomato^flox^) that allow labeling of surfactant protein C-positive (SFTPC+) AEC2s. Such cells can also be filtered through the additional use of Lysotracker, a dye that stains lysosomes and therefore detects lamellar bodies that are abundant in AEC2s [17]. A population of rMCs that expresses fibroblast growth factor 10 (FGF10) and contains neutral lipids (therefore possessing a lipofibroblast-like phenotype) promotes the formation of AEC2-derived alveolospheres containing AEC2s and AEC1s [18,19]. Alveolospheres had been initially described by co-culturing lineage-labeled AEC2s with platelet-derived growth factor receptor alpha (PDGFRα)-expressing mesenchymal cells, also termed type 2-associated stromal cells (TASCs), that display lipofibroblast characteristics [10,20]. The co-culture assay using Sca-1+ mesenchymal cells and lung epithelial progenitor cells had already been developed [21].
To overcome the need for transgenic mice for the generation of alveolospheres, we hereby provide a detailed protocol for culturing such organoids starting with wild-type (WT) mouse lungs. We validated various aspects of our approach including colony-forming efficiency (CFE), alveolosphere diameter, and cellular differentiation by comparison with alveolospheres obtained from Sftpc^Cre-ERT2/+^; tdTomato^flox^ mice. Our protocol serves as a resource for the lung stem-cell research community interested in AEC2 self-renewal and differentiation, and interaction with mesenchymal niche cells.
2. Materials and Methods
2.1. Mice and Tamoxifen Administration
Mice between 8 and 12 weeks of age were used and maintained on the C57BL/6 background. Male wild-type (WT) mice were purchased from Charles River (Strain code 632, Wilmington, MA, USA). Sftpc^tm1(Cre/ERT2,rtTA)Hap^ (or simply Sftpc^Cre-ERT2^) mice were previously described [22], and Gt(ROSA)26Sor^tm9(CAG-tdTomato)Hze^ (or simply tdTomato^flox^) mice were purchased from the Jackson Laboratory (stock number 007905, Bar Harbor, ME, USA). The two lines were bred to generate Sftpc^Cre-ERT2/+^; tdTomato^flox^ mice. Tamoxifen powder (Sigma-Aldrich, T5648, St. Louis, MO, USA) was dissolved in corn oil (Sigma-Aldrich, C8267) and injected intraperitoneally three times at a dose of 0.25 mg/g body weight per injection. Mice were kept under specific pathogen-free (SPF) conditions with unlimited access to food and water.
2.2. Lung Dissociation and Fluorescence-Activated Cell Sorting
Lungs from C57BL/6 (WT) and Sftpc^Cre-ERT2/+^; tdTomato^flox^ mice were collected for single-cell suspension preparation, as previously described [16]. Briefly, the lungs were flushed via transcardiac perfusion using 20 mL of Hank’s balanced salt solution (HBSS). Digestion was performed by intratracheal injection of 2 mL of dispase (5 U/mL; BD Biosciences, 354235, Franklin Lakes, NJ, USA) and incubation in 3 mL of dispase solution (5 U/mL) at room temperature (RT) for 40 min. After digestion, lung homogenates were passed through 70 µm and 40 µm cell strainers. The cell suspension was centrifuged and incubated with an antibody cocktail containing biotin rat anti-mouse CD45 (BD Biosciences, 553078), CD16/32 (BD Biosciences, 553143), and CD31 (BD Biosciences, 553371) at 37 °C for 30 min to deplete hematopoietic and endothelial cells. After centrifugation, cells were resuspended in DMEM (Dulbecco’s Modified Eagle Medium) containing Dynabeads Biotin Binder (Invitrogen, 11047, Waltham, MA, USA) for 30 min at RT on a rotator. For magnetic separation, cells were transferred to a magnetic rack for 10 min at RT. Cell suspensions were then blocked with gamma globulins (Gamunex 10%; Grifols, Barcelona, Spain), re-suspended in MASC buffer (PBS 1X, FBS 0.5%, UltraPure EDTA (Invitrogen, 15575-038, 2 mM)) and stained with anti-mouse CD31 (Alexa Fluor 488, Biolegend, 102514, 1:50, San Diego, CA, USA), CD45 (FITC, BD Biosciences, 553080, 1:50), CD326 (APC/Cy7, Biolegend, 118218, 1:50), Sca-1 (Pacific Blue, Biolegend, 108120, 1:50, Santa Monica, CA, USA), and Podoplanin antibodies (APC, Biolegend, 127410, 1:50) for 15 min at 4 °C. Next, CD45− CD31− EpCAM− Sca-1+ rMCs and CD45− CD31− EpCAM^low^ AECs were isolated by FACS. At least 40,000 rMCs and 350,000 AECs from one WT mouse and 250,000 tdTomato+ (tdTom+) cells from one TG mouse can be sorted. To get a higher yield of rMCs, WT lungs can be separately digested with collagenase type IV as previously described [6]. Sorting was performed using FACSAria III cell sorter (BD Biosciences). Data were analyzed using FlowJo software, version 10.10.0 (FlowJo LLC, Ashland, OR, USA).
2.3. Alveolosphere Generation and Analysis
To generate alveolospheres, 17,000 rMCs and 5000 AECs from WT and Sftpc^Cre-ERT2/+^; tdTomato^flox^ mice were flow-sorted, mixed, and resuspended in 50 µL of culture medium containing minimum essential medium (MEM) alpha (Gibco, 41061029, Waltham, MA, USA), 10% Fetal Bovine Serum (FBS) (Gibco, 10270-106), 1% L-glutamine–penicillin–streptomycin (Sigma-Aldrich, G1146), 1% insulin/transferrin/selenium (ITS) (Gibco, 41400-045), and 0.0002% heparin (Stem Cell Technologies, 07980, Waltham, MA, USA). Next, 50 µL of cold growth factor-reduced Phenol Red-free Matrigel (Corning, 356231, Corning, NY, USA) was mixed at a 1:1 ratio with the cell suspension, transferred to a 24-well plate on a 0.4 µm cell culture insert (Millipore, PICM01250, Burlington, MA, USA), and incubated at 37 °C for 15 min for polymerization. After incubation, 350 µL of medium was added to the lower chamber and incubated at 37 °C with 5% CO_2_ for up to 16 days. To test the effect of IL-1β on alveolosphere formation, recombinant murine IL-1β (PeproTech, 211-11B-10UG, Waltham, MA, USA) was added to the organoid medium at a concentration of 20 ng/mL. The medium was changed every 2 days. Cultures were imaged using EVOS M7000 (Thermo Fisher, AMF7000, Waltham, MA, USA). The number and area of alveolospheres were measured by ImageJ (v2.14.0/1.54f). The colony-forming efficiency (CFE) was determined by dividing the number of counted alveolospheres by the initial number of seeded AECs.
2.4. Induction of Cre Activity in Ex Vivo Alveolosphere Cultures
To induce Cre-mediated recombination ex vivo, organoid medium containing 1 µM of 4-Hydroxytamoxifen (4-OHT) (Merck, H6278, Darmstadt, Germany) was added to the organoid cultures on day 1. After 24 h, the organoid medium containing 4-OHT was removed, and the organoid inserts were washed twice with 1X PBS. Then, the inserts were placed on a new 24-well plate and alveolospheres were cultured in organoid medium until day 16.
2.5. Whole-Mount Immunofluorescence
On day 16 of culture, alveolospheres were fixed with 4% paraformaldehyde (PFA) (Merck, 104005) for 20 min at RT, then washed three times with 1X PBS. Fixed alveolospheres were blocked and permeabilized with blocking buffer (5% serum, 0.5% Triton-X-100 in 1X PBS) overnight at 4 °C. The serum was chosen from the same species as the host of the secondary antibody. For immunofluorescence, alveolospheres were incubated with primary antibodies in a blocking buffer overnight at 4 °C. The next day, washing buffer (2% serum, 0.3% Triton-X-100 in 1X PBS) was added 3 times for 10–15 min, and secondary antibodies were added overnight at 4 °C. To stain nuclei, the NucBlue™ Fixed Cell ReadyProbes™ Reagent (Thermo Fisher) was utilized. Lastly, stained alveolospheres were washed three times with washing buffer for 10–15 min. The Matrigel containing the organoids was cut out of the insert with a scalpel. The sample was placed on a slide and covered with 5–6 drops of fructose–glycerol clearing solution (60% glycerol, 2.5 M fructose) [23]. A coverslip was placed on the sample and after drying for 1 day at 4 °C in the dark, imaging was carried out using an SP8 confocal microscope (Leica Microsystems, Wetzlar, Germany) controlled by LAS X software (version 3.5.7). For capturing the confocal z-stacks, we used 20x0.75NA air and 63xW 1.2NA objectives, while the scan settings were kept the same across all acquisitions. A list of primary and secondary antibodies can be found in Table 1.
2.6. Image Analysis
The acquired confocal images were cleared computationally, using the Lightning module of LAS X software. Subsequently, for the maximum intensity projection and quantification of the z-stack, we used the ImageJ/Fiji software [24]. With Fiji, every projected image was thresholded using the default integrated algorithm. Then, the area occupied by the positive signal (of each marker) as well as the total organoid area were measured. The final area ratio of each marker was calculated by dividing the marker’s area by the organoid’s area.
2.7. RNA Extraction and Gene Expression Analysis
RNA was extracted from alveolosphere cultures using the RNeasy micro kit (Qiagen, 74004, Hilden, Germany). cDNA synthesis was performed using the Quantitect reverse transcription kit (Qiagen, 205311) according to the manufacturer’s instructions. Quantitative real-time PCR (qPCR) was performed using a PowerUp SYBR green master mix (Thermo Fisher Scientific, A25742) and a LightCycler 480 II machine (Roche Applied Science, Penzberg, Germany). Gene expression levels were normalized to glyceraldehyde-3-phosphate dehydrogenase (Gapdh) and presented as fold change relative to day 8 of culture. Primer sequences are shown in Table 2. Bulk RNA sequencing was carried out according to standard procedures. For genome-wide analysis of gene expression, RNA sequencing libraries from polyadenylated mRNA were generated and sequenced using Illumina NovaSeq 6000 at the Institute for Lung Health (ILH)-Genomics and Bioinformatics at the Justus-Liebig University (JLU) Giessen (Germany).
2.8. Statistical Analysis
The number of biological replicates is provided in the corresponding figure legends. All data are presented as mean ± standard error of the mean (SEM). Student’s t test (unpaired, two tailed) was used to compare the means of two groups and One-way ANOVA was used to compare more than two groups. The differences in the means were considered statistically significant when the p value was less than 0.05. Data were assembled and analyzed using GraphPad Prism software, version 9.5.1 (528) (GraphPad Software, Boston, MA, USA).
3. Results
3.1. AEC2s Can Be Isolated from WT Mouse Lungs by Sorting CD45− CD31− EpCAMlow Cells
To show a proof of concept that alveolospheres can be generated from AECs that are FACS sorted from WT lungs, we first carried out EpCAM staining on AEC cell suspensions prepared from Sftpc^Cre-ERT2/+^; tdTomato^flox^ lungs (Figure 1a). We identified EpCAM^low^ (66.1% ± 8.4% of AECs) and EpCAM^high^ (9.6% ± 1.6% of AECs) populations (Figure 1b). We have previously demonstrated that the EpCAM^low^ population is enriched with AEC2s [19], while the EpCAM^high^ cell population is enriched with airway cells such as ciliated and secretory cells [19] as well as bronchioalveolar stem cells (BASCs) [9]. Moreover, tdTom^low^ (22.2% ± 4.8% of AECs) and tdTom^high^ (45.2% ± 5.5% of AECs) corresponding to injury-activated alveolar progenitors (IAAPs) and mature AEC2s, respectively, were also identified (Figure 1c) as previously described [16]. IAAPs have been shown to be inefficient in generating alveolospheres [16]. Total tdTom+ cells accounted for around two-thirds of the AEC preparation (Figure 1c), similar to EpCAM^low^ cells (Figure 1b).
Gating for total tdTom+ cells (tdTom^low^ and tdTom^high^) showed that 91.4% ± 2.1% of these cells were contained in the EpCAM^low^ population compared to traces in EpCAM^high^ cells (Figure 1d). Gating for tdTom^high^ and tdTom^low^ populations showed similar results (Figure 1e,f). Further analysis of the EpCAM^low^ population showed that 95% ± 0.42% of these cells were tdTom+, with the majority being tdTom^high^ (64.1% ± 4.5%) and a minor proportion of tdTom^low^ (30.2% ± 4.7%) (Figure 1g). On the other hand, the EpCAM^high^ population showed lower proportions of tdTom+ cells (Figure 1h). Therefore, the EpCAM^low^ population was confirmed to be enriched with AEC2s.
Next, rMCs were co-cultured with either AECs isolated from WT lungs (hereafter referred to as the WT approach) or tdTom+ cells from Sftpc^Cre-ERT2/+^; tdTomato^flox^ lungs (hereafter referred to as the transgenic (TG) approach) (Figure 2a). WT AECs were sorted from single-cell suspensions based on CD45− CD31− EpCAM^low^ surface marker expression (Figure 2b). The majority of isolated cells were identified as CD45− CD31− EpCAM^low^ PDPN− AEC2s (90.2% ± 1.01% from the parent EpCAM^low^ gate), while only 8.8% ± 0.47% was CD45− CD31− EpCAM^low^ PDPN+ AEC1s (Figure 2c). Lungs from Sftpc^Cre-ERT2/+^; tdTomato^flox^ mice were similarly processed for FACS sorting. Staining for EpCAM and PDPN revealed comparable proportions of AEC2s and AEC1s within the EpCAM^low^ population similar to the WT approach (Figure 2d,e). Since IAAPs are quiescent in the absence of injury and therefore do not robustly generate alveolospheres [16], tdTom^high^ cells were initially selected for alveolosphere generation (Figure 2f). As shown in Figure 1e, 92.5% ± 2.4% of tdTom^high^ cells are found in the EpCAM^low^ gate. Staining for PDPN revealed that 90.1% ± 0.5% of tdTom^high^ EpCAM^low^ cells were AEC2s, while only 8.46% ± 0.45% were AEC1s (Figure 2g), resembling the data attained with WT EpCAM^low^ cells.
3.2. The EpCAMlow Population Robustly Generates Alveolospheres and It Is Comparable with Sorted Lineage-Traced AEC2s
The data described above clearly show that gating for CD45− CD31− EpCAM^low^ cells in WT lungs allows for the sorting of an AEC population that predominantly consists of AEC2s. The finding that lineage-traced AEC2s essentially belong to the EpCAM^low^ population (Figure 1d–f) strongly supports this conclusion. We therefore tested the ability of EpCAM^low^ cells to generate alveolospheres (WT approach) and compared the results to the TG approach. As expected, all alveolospheres in the TG approach cells were tdTom+ (Figure 2h). Both approaches were similarly efficient in generating alveolospheres (WT CFE 3.14% ± 0.23% vs. TG CFE 2.55% ± 0.27%) (Figure 2i) with comparable alveolosphere diameter (139.37 μm ± 6.86 μm for WT vs. 130.57 μm ± 5.01 μm for TG) (Figure 2j). Gene expression analysis was then carried out on WT and TG alveolospheres on day 8 (when AECs start to form cysts) and day 16 (when alveolospheres acquire a complex morphology and contain differentiated AEC2s and AEC1s). The results showed significant upregulation of the AEC2 marker Sftpc (Figure 2k,m) and the AEC1 marker Ager (Figure 2l,n) in both conditions.
3.3. WT Alveolospheres Derive from Isolated AEC2s
To validate that the organoids indeed derive from AEC2s, the CD45− CD31− EpCAM^low^ tdTom− AEC population was isolated from uninduced Sftpc^Cre-ERT2/+^; tdTomato^flox^ lungs and co-cultured with WT rMCs. Cre-mediated recombination was induced ex vivo by exposure to the active metabolite of tamoxifen, 4-OHT, between days 1 and 2 (Figure 3a–d). By day 2, cultures revealed single tdTom+ cells (Figure 3e), with most alveolospheres (90.61% ± 1.45%) being tdTom+ on day 16 (Figure 3f,h). In the absence of 4-OHT, organoids were negative for tdTom (Figure 3g). These data strengthen the notion that the EpCAM^low^ population indeed enriches for AEC2s that are capable of ex vivo expansion and subsequent alveolosphere formation.
Alveolospheres generated using the WT, TG, and 4-OHT (TG) approaches were then subjected to immunofluorescence for SFTPC and AGER, followed by whole-mount confocal imaging (Figure 3i). The data revealed similar ratios relative to alveolosphere area among the three groups for SFTPC (WT 0.64 ± 0.01 vs. TG 0.54 ± 0.07 vs. 4-OHT (TG) 0.52 ± 0.02) (Figure 3j) and AGER (WT 0.52 ± 0.04 vs. TG 0.42 ± 0.09 vs. 4-OHT (TG) 0.46 ± 0.1) (Figure 3k), without apparent differences in terms of overall morphology, suggesting that the cellular composition within these conditions is highly comparable.
To further demonstrate that the alveolospheres generated using the WT approach are comparable to those generated using the TG approach, we carried out RNA-seq on isolated WT AECs, tdTom+ cells, and the corresponding organoids (Figure 4a). The full list of genes is shown in Supplementary Table S1. Notably, the same gene sets are modulated during organoid formation regardless of whether the starting material is tdTom+ cells or WT AECs (Figure 4b,c). Moreover, a comparison of AEC2 and AEC1 markers showed no difference between the two groups (Figure 4d,e), confirming that the cell population isolated using our WT EpCAM^low^ approach is very similar to the tdTom+ population isolated from transgenic animals. We also found that the AEC1 signature is upregulated in alveolospheres compared with isolated AEC2s, while the AEC2 signature is diluted in alveolospheres (Figure 4d,e), which is expected given the emergence of AEC1s and the abundance of mesenchymal cells in the alveolospheres but not in purified AEC2s. Pathway analysis of the top genes differentially regulated between alveolospheres and AEC2s was related to ECM organization, elastic fibers, collagen biosynthesis, and cell motility (Figure 4f).
3.4. Stimulation with Recombinant Murine IL-1β Alters Alveolosphere Formation
It has been reported that IL-1β treatment leads to enlarged alveolospheres that feature accumulation of the KRT8+ intermediate cell state [12]; therefore, recombinant IL-1β was added to WT cultures as another proof of concept that such organoids behave similarly to those obtained using the TG approach (Figure 5a). After 16 days of culture, organoids clearly showed a cyst-like morphology rather than a typical alveolosphere morphology in the IL-1β group (Figure 5b), which was also reflected by the quantification of organoid diameter (285.08 μm ± 11.6 μm vs. 145.04 μm ± 9.16 μm) (Figure 5d). Analysis of the CFE did not reveal a difference between the two groups (Figure 5c). Notably, immunofluorescence showed a significant increase in the coverage of the KRT8 signal relative to the overall alveolosphere area upon IL-1β treatment (0.36 ± 0.05 vs. 0.16 ± 0.05) (Figure 5e,f). These data indicate that our WT approach can recapitulate previous findings attained using the TG approach.
4. Discussion
Over the past decade, lung organoids have emerged as important models to investigate morphogenic and regenerative aspects of lung biology and disease such as assessing epithelial stem/progenitor cell self-renewal, self-organization, and differentiation, as well as the supportive potential (or niche activity) of corresponding mesenchymal cells. These organoid systems mimic various anatomical regions of the mammalian lung and are therefore dubbed tracheospheres, bronchiolospheres, bronchioalveolar organoids, and alveolospheres [3,25,26,27,28]. As with any culture assay, there is a strong need for standardization of the operating procedure to maximize data quality, robustness, and reproducibility.
One of the pioneering studies in the field is that of McQualter et al., who demonstrated the ability of lung EpCAM+ cells to form airway, alveolar, and mixed organoids when co-cultured with EpCAM− Sca-1+ mesenchymal cells in Matrigel [21]. The generation of organoids from murine and human AEC2s was later described, where the co-culture of AEC2s with PDGFRα+ mesenchymal cells or TASCs, a population that includes lipofibroblasts, led to the formation of alveolospheres containing AEC2s and AEC1s [10,20]. Alveolosphere cultures were also used to characterize the WNT-responsive fraction of AEC2s in the murine lung and investigate alveolar niche interactions. Such an AEC2 subset displayed superior alveolosphere formation capacity and responds to WNT and FGF stimulation [14,29,30]. Another AEC2 subset that acts as a quiescent repertoire for mature AEC2s is the IAAPs, which do not show robust alveolosphere formation capacity but are important for replenishing the bona fide AEC2 pool [15,16,31,32].
WNT and FGF signaling have been established as critical determinants of the AEC2 cell fate by independent studies [7,31,33,34,35,36]. The mesenchymal niche can be fractionated into AXIN2+ (a WNT-responsive population enriched with Fgf7 expression) and FGF10+ subsets. Both fractions show lipofibroblast features including proximity to AEC2s and neutral lipid content [18,37]. How distinct these two subsets are, especially in terms of in vivo function during homeostasis, repair, and regeneration, remains to be seen. A third niche population is represented by WNT ligand-producing leucine-rich repeat-containing G protein-coupled receptor-5-positive (LGR5+) fibroblasts that have shown the ability to support alveolosphere formation [7]. In the current protocol, the bulk rMC population was used to achieve a simple and universal protocol to generate alveolospheres from WT mice without the need for transgenic animals or discriminating between rMC subsets. Table 3 highlights the main features of some of the published protocols to generate murine alveolospheres. The emerging picture is that (1) most of the studies listed below relied on the use of transgenic mice to generate alveolospheres, (2) a starting material of 5000 AEC2s combined with mesenchymal cells at a ratio of 1:10 and later mixed with Matrigel (1:1) is the most used approach, and (3) the data show varying CFE values even when using similar protocols.
5. Conclusions
Our protocol thus allows robust and reproducible generation of murine alveolsopheres using the EpCAM^low^ gating strategy, which represents a refinement of previous EpCAM gating strategies and avoids lumping EpCAM^high^ with EpCAM^low^ cells. As club cells and BASCs are not found in the EpCAM^low^ fraction [9,19], our approach minimizes contamination with bronchiolar and bronchioalveolar organoids, allowing to predominantly obtain alveolospheres. We believe that our protocol represents a step toward the standardization of this important research tool.
Finally, we were able to reproduce the effect of IL-1β on alveolosphere formation using our WT approach. The accumulation of KRT8+ transitional cells was evident based on whole-mount immunofluorescence data. The involvement of these and other transitional AEC2 populations in various respiratory diseases has gained tremendous interest in recent years [11,13] and further research is warranted to fully understand this phenomenon, decipher how to convert such dysplastic events into healthy scarless regeneration, and ultimately reconstitute the alveolar stem cell niche and restore optimal gas exchange in the diseased lung.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lancaster M.A. Knoblich J.A. Organogenesis in a Dish: Modeling Development and Disease Using Organoid Technologies Science 2014345124712510.1126/science.124712525035496 · doi ↗ · pubmed ↗
- 2Zhao Z. Chen X. Dowbaj A.M. Sljukic A. Bratlie K. Lin L. Fong E.L.S. Balachander G.M. Chen Z. Soragni A. Organoids Nat. Rev. Methods Primer 202229410.1038/s 43586-022-00174-y PMC 1027032537325195 · doi ↗ · pubmed ↗
- 3Vazquez-Armendariz A.I. Herold S. From Clones to Buds and Branches: The Use of Lung Organoids to Model Branching Morphogenesis Ex Vivo Front. Cell Dev. Biol.2021963157910.3389/fcell.2021.63157933748115 PMC 7969706 · doi ↗ · pubmed ↗
- 4Rock J.R. Onaitis M.W. Rawlins E.L. Lu Y. Clark C.P. Xue Y. Randell S.H. Hogan B.L. Basal Cells as Stem Cells of the Mouse Trachea and Human Airway Epithelium Proc. Natl. Acad. Sci. USA 2009106127711277510.1073/pnas.090685010619625615 PMC 2714281 · doi ↗ · pubmed ↗
- 5Vazquez-Armendariz A.I. Seeger W. Herold S. El Agha E. Protocol for the Generation of Murine Bronchiolospheres STAR Protoc.2021210059410.1016/j.xpro.2021.10059434169288 PMC 8209683 · doi ↗ · pubmed ↗
- 6Moiseenko A. Vazquez-Armendariz A.I. Kheirollahi V. Chu X. Tata A. Rivetti S. Günther S. Lebrigand K. Herold S. Braun T. Identification of a Repair-Supportive Mesenchymal Cell Population during Airway Epithelial Regeneration Cell Rep.20203310854910.1016/j.celrep.2020.10854933357434 PMC 8363050 · doi ↗ · pubmed ↗
- 7Lee J.H. Tammela T. Hofree M. Choi J. Marjanovic N.D. Han S. Canner D. Wu K. Paschini M. Bhang D.H. Anatomically and Functionally Distinct Lung Mesenchymal Populations Marked by Lgr 5 and Lgr 6Cell 201717011491163.e 1210.1016/j.cell.2017.07.02828886383 PMC 5607351 · doi ↗ · pubmed ↗
- 8Chu X. Lingampally A. Moiseenko A. Kheirollahi V. Vazquez-Armendariz A.I. Koepke J. Khadim A. Kiliaris G. Shahriari Felordi M. Zabihi M. GLI 1+ Cells Are a Source of Repair-Supportive Mesenchymal Cells (RSM Cs) during Airway Epithelial Regeneration Cell. Mol. Life Sci.20227958110.1007/s 00018-022-04599-236333491 PMC 9636301 · doi ↗ · pubmed ↗