Differentiation of Synanthropic Fleas from Andalusia (Spain) through Geometric Morphometrics Analysis
Angela M. García-Sánchez, Ignacio Trujillo, Antonio Zurita, Cristina Cutillas

TL;DR
This study uses geometric morphometrics to differentiate flea species in Andalusia, showing it's a useful tool for identifying and managing these disease-carrying insects.
Contribution
The study demonstrates the effectiveness of geometric morphometrics in differentiating synanthropic flea populations in Andalusia.
Findings
Geometric morphometrics successfully differentiated three flea populations with minimal overlap.
Size differences were observed, with P. irritans males and A. erinacei females being larger.
The method proved useful for identifying fleas lacking clear morphological features.
Abstract
Fleas are blood-sucking insects that are not only a nuisance but can also act as vectors for various diseases in animals and humans, including dangerous ailments such as the bubonic plague. Identifying and classifying these insects accurately is crucial for understanding how they spread and how to control them. Geometric morphometrics, a cutting-edge technique, is proving to be an invaluable tool in this regard, alongside traditional methods and molecular biology. In the present study conducted in Andalusia, Spain, this technique successfully differentiated between three populations of fleas, providing insights into their distribution, size, and characteristics. Image processing software was employed to obtain measurements, such as perimeters and areas, of the fleas under study. These findings underscore the importance of geometric morphometrics in studying and managing arthropod…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
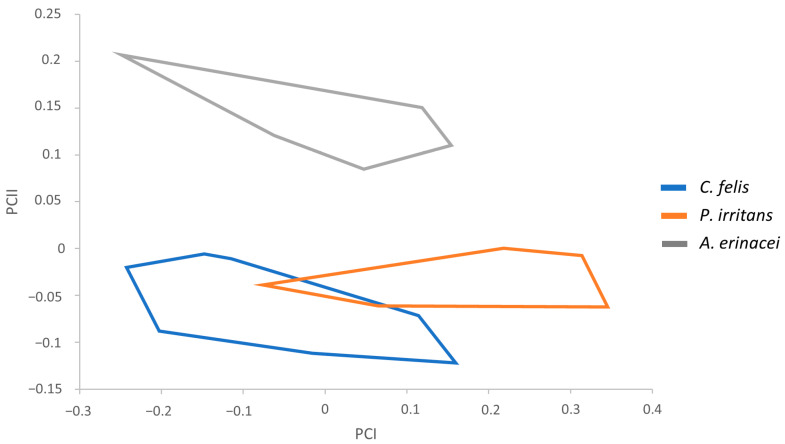 Figure 1
Figure 1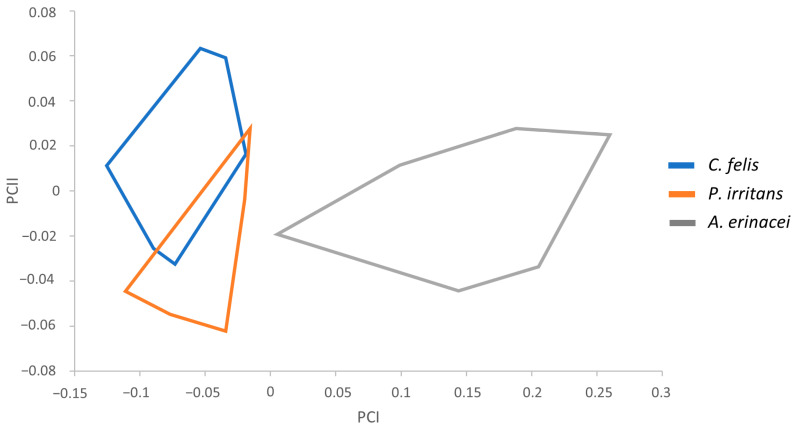 Figure 2
Figure 2- —Junta de Andalucía, European Regional Development Fund (FEDER)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsYersinia bacterium, plague, ectoparasites research · Zoonotic diseases and public health · Morphological variations and asymmetry
1. Introduction
Fleas (Siphonaptera) comprise a highly specialized order of holometabolous ectoparasitic insects with a cosmopolitan distribution and about 2700 species described so far [1]. In addition to being able to provoke bites and pruritic welts on the skin, these arthropods are also known to be vectors of different pathogens, responsible for causing a wide variety of diseases worldwide, including bartonellosis, rickettsiosis, and bubonic plague [2,3,4,5,6,7,8,9]. This is due to the nature of some flea species, which present a low host specificity that facilitates the exchanging of microorganisms, posing a potential threat to the health of both humans and animals [10,11,12].
The prevention and control of fleas require a large investment of money per year, which represents a significant economic burden [13]. It is crucial to enhance our knowledge about the taxonomy of fleas to develop effective strategies to reduce flea infestations and their negative impact on our environment.
Recently, in addition to the traditional morphological identification and molecular biology approaches, geometric morphometrics has proven to be a useful complementary technique for discriminating taxa across different groups [14,15,16]. One of the main features of geometric morphometrics is that it is especially helpful in cases of taxa that present morphological ambiguity [15], a situation relatively common in fleas [17,18,19]. This scenario invited the exploration of the affordable criterion offered by geometric morphometrics in systematic studies on flea genera, with promising results in Ctenocephalides Stiles & Collins, 1930 [20,21], Ctenophthalmus Kolenati, 1856 [22], Pulex Linnaeus, 1758 [23], and Stenoponia Jordan & Rothschild, 1911 [24].
In Europe, there is evidence of an escalating frequency of vector-borne diseases and heightened pathogen circulation, primarily influenced by human-related factors [25]. One region remarkably affected by arthropod-borne diseases is Andalusia, situated in the southern of Spain, where the West Nile Virus circulation is more widespread than initially considered [26], and outbreaks have been reported in recent years [27].
Furthermore, the incidence of murine typhus, a zoonosis caused by Rickettsia typhi da Rocha Lima, 1916 transmitted to humans by fleas, seems to be increasing slowly in Andalusia [28], an aspect that reveals the ability of some flea species present in the region to transmit pathogenic bacteria. Hence, it is essential to resort to techniques that allow us to safely discern between taxa.
The cat flea, Ctenocephalides felis Bouché, 1835, succeeded in its expansion as a global parasite, and it is one of the most common flea species identified in domestic dogs and cats worldwide [11,19]. In Southwestern Europe, C. felis is the dominant species, although, in Eastern Europe, the infestation by Ctenocephalides canis Curtis, 1826 and Pulex irritans Linnaeus, 1758 also occurs [19,29,30,31].
On the other hand, hedgehogs inhabit rural, urban, and suburban environments, and they are frequently parasitized by blood-sucking arthropods, such as hard ticks and fleas, including the hedgehog flea, Archaeopsylla erinacei Bouché, 1835, and other flea species such as C. felis, C. canis, and Nosopsyllus fasciatus Bosc, 1800 [32,33,34,35]. Since they usually cohabit with pets and humans, they can potentially act as reservoirs of pathogen microorganisms responsible for zoonoses [12].
The main objective of the present study was to determine the capacity of geometric morphometric analysis to identify and discriminate fleas from populations of C. felis, P. irritans, and A. erinacei collected in Andalusia, in order to strengthen its role as a useful complementary technique in arthropod taxonomical studies.
2. Materials and Methods
2.1. Collection of Samples
Over a period of 19 months, we collected flea samples from dogs (Canis lupus familiaris Linnaeus, 1758) and one hedgehog (Erinaceus europaeus Linnaeus, 1758) that coexisted with other dogs.
To gather flea samples from the hosts, we reached out to some veterinary clinics, veterinary hospitals, pet shelters, and some pet owners. In total, we contacted 145 veterinary clinics and 30 pet shelters and kennels. Among these, 18 centers agreed to participate in the sample collection (see Acknowledgements). All participants volunteered for this sampling process. Only animals parasitized by fleas were sampled. Veterinary practitioners performed an initial inspection of pets brought to their facilities. Each pet was checked for fleas and examined by a veterinarian who recorded clinical signs related to flea infestation. Adult flea counts were conducted according to the World Association for the Advancement of Veterinary Parasitology guidelines [36]. In brief, the animals were combed over their entire bodies with a fine-toothed comb for 5–10 min.
All captured fleas from each infested host were transferred to a plastic 1.5 mL tube containing 96% ethanol for subsequent identification and morphometrics analyses.
2.2. Morphological Identification and Metric Data Processing
For morphological analysis, all specimens were initially examined under an optical microscope for specific classification. Following this, the specimens were cleared using 10% KOH, prepared, and mounted on glass slides following conventional procedures with the EUKITT mounting medium (O. Kindler GmbH & Co., Freiburg, Germany) [37]. The cleared and mounted specimens were examined again for a more detailed morphological analysis using a BX61 microscope (Olympus, Tokyo, Japan) and submitted to image capture processes with the imaging software cellSens Standard version 4.2 (Olympus, Tokyo, Japan). The diagnostic morphological characters of all samples were analyzed by comparison with figures, keys, and descriptions reported previously [38,39,40,41,42]. The measurement images of each flea were made using the image analysis software Image-PRO v11 (Media Cybernetics, Rockville, MD, USA). A total of 28 different parameters were measured for males and 36 for females (Table 1 and Table 2).
Descriptive univariate statistics based on arithmetic mean, standard deviation, and coefficient of variation for all parameters were determined for male and female populations. The data were subjected to one-way ANOVA (analysis of variance) for statistical analysis of the parameters. The results were statistically significant when p < 0.05. Statistical analysis was performed using Microsoft Excel for Microsoft 365 MSO (v2402). In addition, biometric characters of fleas were compared between different species and the most significant parameters were assayed for a morphometrics study.
Morphological variation was quantified using geometric morphometrics [43], a technique that provides an estimate of size integrating different growth axes into a single variable known as “centroid size” [44]. The estimate of size was represented by a single variable that reflected variation in multiple directions, as many as there were landmarks under study, and shape was defined as their relative positions after correction for size, position, and orientation. With these informative data, and the corresponding software freely available to conduct complex analyses, significant biological and epidemiological features can be quantified more accurately [45].
Multivariate analyses were applied to assess phenotypic variations among the samples, using size-free canonical discriminant analysis on the covariance of log-transformed measurements. These analyses are applied to exclude the effect of within-group ontogenetic variations by reducing the effect of each character on the first pooled within-group principal component (a multivariate size estimator) [46]. principal component analysis (PCA) was used to summarize most of the variations in a multivariate dataset in a few dimensions [47]. Morphometric data were explored using multivariate analysis in three parameters in males (TW, HW, and AW) (Table 1) and females (Global Circularity, BULGAP, and HL) in females (Table 2) using BAC v.2 software [21,48].
3. Results
A total of 81 fleas (34 males and 47 females) were collected from different regions of Andalusia and classified as follows: 39 as C. felis (18 males and 21 females), 19 as P. irritans (6 males and 13 females), and 23 as A. erinacei (10 males and 13 females) (Table 3). All C. felis specimens and three P. irritans (the only male from Huelva and two female fleas from Seville) were collected from dogs (Canis lupus familiaris), whereas A. erinacei were collected from a hedgehog host. The rest of the P. irritans fleas were collected off-host from a neglected horse stable (Table 3).
To carry out the classification of the samples, we considered descriptions used traditionally to discern between these species and, additionally, remarkable morphological features based on the measurements performed. Statistical tests showed several significant measurements for subsequent morphometric analyses. Therefore, the following parameters were used: total width (TW), total width of the head (HW), and apex width (AW) in males (Table 1) and total length of the head (HL), perimeter of the bulga (BULGAP), and Global Circularity in females (Table 2). This perimeter is the length of the bulga’s boundary, whereas the circularity is calculated as the ratio of the area of an object against a circle whose diameter is equal to the object’s maximum feret. The influence of size was analyzed using PCA in C. felis, P. irritans, and A. erinacei, involving the regression of each character separately on the within-group first principal component (PC1). The resulting factor maps for male and female populations are shown in Figure 1 and Figure 2, respectively.
Male variables significantly correlated with PC1, contributing 71% to the overall variation. The male factor maps showed global size differences in the flea populations, with a slightly larger size in P. irritans males (Figure 1). The three male communities are well grouped in the factor map, with a lack of noteworthy overlapping areas between them. Only C. felis and P. irritans showed a partial overlap but with no inconvenience in their identification.
On the other hand, female variables significantly correlated with PC1, contributing 90% to the overall variation. The resulting factor maps (Figure 2) clearly illustrate global size differences in the populations analyzed, including a bigger size in A. erinacei. As in the previous factor maps, there is a lack of notable overlapping areas between the female populations. As in the male factor maps, C. felis and P. irritans showed a partial overlap that did not prevent their identification.
4. Discussion
The accurate classification of fleas requires careful morphological examination or molecular confirmation, and therefore, the possibility that prior studies may have inadvertently misidentified fleas cannot be discarded [19,49]. In fact, authors like Ménier and Beaucournu reported numerous misidentifications in specimens of the genus Ctenocephalides [50]. The study of flea epidemiology, control, and prevention requires the accurate identification of species and subspecies.
In general, the classification of genera and species of fleas is based on external morphological characters. The presence or absence of combs and eyes, along with the length of the head, are typically significant features in morphological identification [51]. However, the size itself could never represent a way to reliably recognize the sex or the species of a flea specimen [52], and some flea species do not have easily identifiable morphological characters. For instance, A. erinacei and P. irritans do not possess pronotal or genal combs, so their classification process can be more complex in case the required specialized skills in flea identification are lacking. It is necessary to pay great attention to detail to recognize developmental stages and adequate sex identification [52]. Additionally, C. felis is a good example of a flea species known for its morphological ambiguity and the underlying issues in the study of their global populations [19]. In terms of molecular biology techniques, the notable lack of large-scale phylogenetic data for flea taxa causes some genera like Ctenocephalides to not have a defined genetic identity [17,19,40], especially if we consider subspecies [19].
Furthermore, it is still surprising that, despite the considerable veterinary and public health significance of dog fleas, studies investigating the diversity of these ectoparasite species on pets and the occurrence of flea-borne pathogens are scarce in certain regions [20].
Given the difficulties associated with flea morphological identification, the limited genetic information available, and the insufficient knowledge of the common pathogens of each flea species, the need to resort to complementary diagnosis techniques arises.
Geometric morphometrics analysis is one of these novel approaches applied to parasitological diagnosis, usually employed in arthropod identification [43]. The technique is based on the utilization of computer software for data processing and interpretation, with the advantage being that costly reagents and equipment are not required. Its affordability and the simplicity of data collection make it especially useful in low-resource settings [52,53].
To the best of our knowledge, the present work is the first in which flea measurements have been obtained using the imaging software Image-PRO. This program has been used before to study arthropods and other parasites [54,55,56,57]. It allowed us to incorporate into the analysis accurate measures such as areas, perimeters and circularities of the flea specimens for the first time. Global Circularity and BULGAP were revealed as useful features that contribute to the identification of flea species via geometric morphometrics.
The distribution of the three analyzed flea species showed a comparable pattern in both factor maps. A. erinacei appeared distant from C. felis and P. irritans, showing an appreciably larger size in the case of females. C. felis and P. irritans presented a small overlapping area, which did not prevent their individual identification. In both cases, P. irritans has always appeared larger than C. felis.
Although A. erinacei and C. felis are part of the Pulicinae family, the factor maps illustrated that fleas at the same taxonomic level are not necessarily closer at the morphological level since A. erinacei appears further away from both C. felis and P. irritans.
The selection of representative measurements for the morphometrics analyses was in accordance with previously published works. Total width (TW), head width (HW), and apex width (AW) are consolidated as useful parameters that define the morphological identity in males [21,50,58], as well as the total length of the head (HL) and the perimeter of the bulga (BULGAP) for females [21]. Due to the lack of genal ctenidium in P. irritans and A. erinacei, the difference in length between first and second spines (DEG parameter [21]) could not be considered in the present analysis, whereas the inclusion of the degree of elongation of the apical part (hilla) in females offered similar results but with a bit more overlap between P. irritans and C. felis. This is why the parameter APEHILL [21] was substituted by BULGAP, which permits the best differentiation between species.
The three analyzed flea species are among the most frequent in our environment, and, as a result, there is a notable risk of encountering them, with consequently associated parasitism suffered by humans and animals. After applying geometric morphometrics to differentiate flea communities of the same genera [21,22,23,24], the present work represents a step further, since this technique allowed us to identify different flea genera.
5. Conclusions
Accurate identification of fleas is necessary for studying the epidemiology, prevention, and control of these arthropods. In situations of uncertainty, alternative approaches are required to ensure correct classification. Geometric morphometrics is increasingly recognized as a reliable complementary technique for identifying flea species, particularly valuable in environments with limited resources.
In the present work, we were able to discern between the flea species A. erinacei, P. irritans, and C. felis using principal component analysis of males and females. Differences in overall size were also detected: A. erinacei presented the largest size in females, whereas P. irritans was slightly larger in males. Therefore, morphometrics is a relevant technique with great potential for application in the field of fleas, considering the existence of species that have traditionally posed challenges in their identification.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hastriter M.W. Bossard R.L. Lewis R.E. Flea (Siphonaptera) World Species List (Spreadsheet)2018 Available online: https://esanetworks.org/group/fleanews/page/flea-species-of-theworld-spreadsheetupdated-3-february-2018(accessed on 3 December 2023)
- 2Fleta Zaragozano J. Rickettsiosis transmitidas por piojos, pulgas y ácaros Med. Integr.200239147152
- 3Parola P. Davoust B. Raoult D. Tick- and flea-borne rickettsial emerging zoonoses Vet. Res.20053646949210.1051/vetres:200500415845235 · doi ↗ · pubmed ↗
- 4De Sousa R. Fournier P.E. Santos-Silva M. Amaro F. Bacellar F. Raoult D. Molecular detection of Rickettsia felis, Rickettsia typhi and two genotypes closely related to Bartonella elizabethae Am. J. Trop. Med. Hyg.20067572773110.4269/ajtmh.2006.75.72717038702 · doi ↗ · pubmed ↗
- 5Beliavskaia A. Tan K.K. Sinha A. Husin N.A. Lim F.S. Loong S.K. Bell-Sakyi L. Carlow C.K.S. Abubakar S. Darby A.C. Metagenomics of culture isolates and insect tissue illuminate the evolution of Wolbachia, Rickettsia and Bartonella symbionts in Ctenocephalides spp. Fleas Microb. Genom.2023900104510.1099/mgen.0.00104537399133 PMC 10438800 · doi ↗ · pubmed ↗
- 6Boyer S. Gillespie T.R. Miarinjara A. Xenopsylla cheopis (rat flea)Trends Parasitol.20223860760810.1016/j.pt.2022.03.00635527197 · doi ↗ · pubmed ↗
- 7Millán J. Sepúlveda-García P. Di Cataldo S. Canales N. Sallaberry-Pincheira N. Painean J. Cevidanes A. Müller A. Molecular identification of Bartonella spp. and Rickettsia felis in fox fleas; Chile Comp. Immunol. Microb.20239610198310.1016/j.cimid.2023.10198337099997 · doi ↗ · pubmed ↗
- 8Sepúlveda-García P. Alabi A. Álvarez K. Rojas L. Mella A. Gonçalves L.R. AndréM.R. Machado R. Müller A. Monti G. Bartonella spp. in households with cats: Risk factors for infection in cats and human exposure One Health 20231610054510.1016/j.onehlt.2023.10054537363214 PMC 10288095 · doi ↗ · pubmed ↗