The Honey Bee Colony’s Criterion for Candidate Selection: “Ongoing” or “One-Shot”?
Luxia Pan, Shiqing Zhong, Tianyu Xu, Weixuan Chen, Zhijiang Zeng

TL;DR
Honey bee colonies select queen candidates based on quality rather than nepotism, optimizing queen rearing through a multi-stage process.
Contribution
This study reveals that honey bee colonies use a quality-based, multi-stage selection process for queen candidates in the absence of nepotism.
Findings
Colonies prefer 1-day-old larvae for queen rearing, with higher survival and gene expression related to reproductive quality.
Queen selection is optimized through stages (larval, capping, emerging), eliminating lower-quality candidates.
Emerging queens from 1-day-old larvae attract more worker followers, indicating superior quality.
Abstract
The honey bee is a typical social insect, and a colony is composed of a queen, workers, and drones. The queen is responsible for reproduction and is essential for the development of the colony. When the queen dies or is injured, workers raise closely related larvae to develop into a queen. Without nepotism, how do colonies choose queen candidates during emergency queen rearing? Are there criteria for queen candidates? Is the colony’s criterion for candidate selection “ongoing” or “one-shot”? Our investigation focused on the emergency queen-rearing process in a natural colony (without nepotism), where we observed and documented the selection of queen candidates from different stages (the larval stage, capping stage, and emerging stage). We examined the physiological indicators and the expression of ovarian development-related genes (vg, hex110, and Jh), and found that the colony would…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1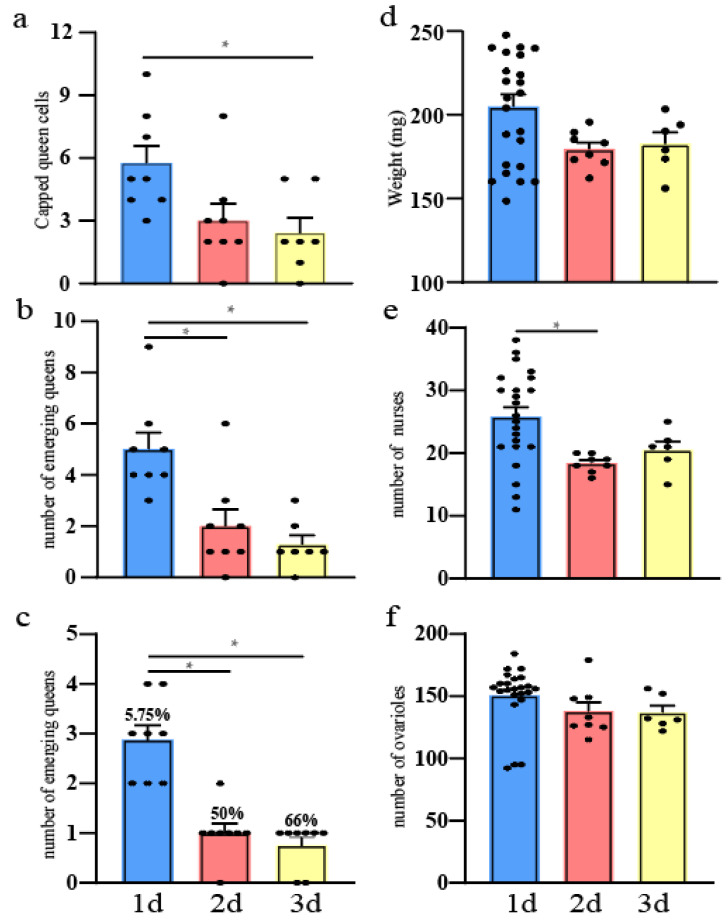 Figure 2
Figure 2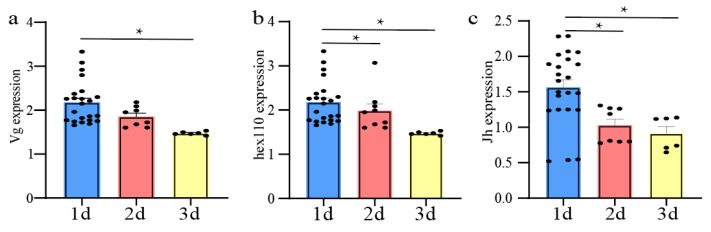 Figure 3
Figure 3- —Earmarked Fund for the China Agricultural Research System
- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Arachnid Ecology and Behavior · Plant and animal studies · Insect and Pesticide Research
1. Introduction
The honey bee queen’s death severely threatens the survival of the colony. To solve this potential problem, honey bees have evolved the ability to breed replacement queens, where in an emergency, new queens are reared from young worker larvae. This process was first demonstrated by Schirack in the late 17th century [1,2], but despite these early works, the process of emergency queen production is not well studied, especially the colony’s criteria for selecting queen candidates.
Social insects use kinship to promote cooperation [3], but in honey bees (Apis mellifera L.), queen breeding can result in conflict due to polygamy [4,5]. In the same subfamily, the average relatedness among worker members is 0.75 (super-sister) [6], the average relatedness between workers and their reproductive offspring (drones) is only 0.5, while the average relatedness between workers in different subfamilies is only 0.25 (half-sisters) [7,8]. This view has led to studies on nepotism as a criterion for workers’ selection of larvae for the rearing of new queens [9]. When the colony is to breed a queen, the workers are faced with choosing between different nepotism indices (r = 0.75, r = 0.5, r = 0.25) for egg and larval breeding [10,11,12]. It is demonstrated that each worker is inclined to cultivate more close sister queens from its paternal line (r = 0.75) through nepotism, rather than raising less related half-sister queens from another paternal line (r = 0.25) [13]. This phenomenon often occurs when the colony’s queen dies/disappears or stops laying eggs, resulting in an emergency queen-rearing situation.
In the absence of nepotism, how do colonies choose queen candidates during emergency queen rearing? Are there criteria for queen candidates and is the colony’s criterion for candidate selection “ongoing” or “one-shot”? To investigate these questions, we chose to study queen rearing in the absence of nepotism, simulating an emergency queen replacement situation, and recording the potential criteria of new queen selections from the larval stage, capping stage, and exit from the chamber stage. The results from this study will aid in the understanding of a colony’s selection of queen candidates in emergency situations.
2. Materials and Methods
2.1. The Colony without Nepotism
Sixteen colonies were used in this study. Eight were kept at the Honeybee Research Institute of Jiangxi Agricultural University (28.46° N, 115.49° E) as egg-laying colonies. Eight colonies were purchased from Gaoan (Jiangxi, China; 28.14° N, 115.29° E) as queenless colonies. The distance between the two locations is 42 km, which ensures no relatedness between colonies.
Eight empty combs were chosen (no brood, pollen, or nectar) and divided into four areas with double-sided tape (Figure 1a). A square queen excluder (21 cm × 21 cm) divided one half of a comb into two areas. A queen from each colony was confined to one area to lay eggs for six hours, after which the queen was separated from the experimental hive using a vertical queen excluder. On days 2–4, the queen was controlled to lay eggs in the other three restricted areas using the same method. The larvae in the four regions were thus 1-day-old, 2-day-old, 3-day-old, and 4-day-old on the seventh day after the start of the experiment.
On the sixth day, queenless colonies were prepared. Each queenless colony had one comb with a lot of honey and approximately 10,000 workers. On the seventh day, we added one of the above experimental combs to a queenless colony, respectively.
2.2. Number of Capped Queen Cells and Newly Emerged Queens
The number of capped queen cells was counted 13 days after the experimental comb was placed in the queenless colony. Then, the number of queens that emerged from the natural queen cells on the first day was counted. The number of capped queen cells and newly emerged queens were analyzed by one-way ANOVA using SPSS 25.0 (IBM, SPSS, Inc., Chicago, IL, USA).
2.3. Nursing Behavior of Worker and Physiological Indicators of Queens
The birth weights of the newly emerged queens were measured with a Precise Electronic Balance (Mettler Toledo, ME204, 0.1 mg). Then, the newly emerged queens were placed in a queen cage and hung in the original colony (>1-day-old). We then counted the number of nurses feeding each newborn queen. Thereafter, each new virgin queen was killed and dissected, and one of its ovaries was used to determine the expression level of reproduction-related genes, while the other ovary was used to make paraffin sections according to the methodology of Yao Yi [12]. The number of ovarioles was counted and analyzed with one-way ANOVA using SPSS 25.0.
2.4. Gene Expression Analysis
Total RNA was extracted from the sampled ovaries using the Trans-Zol Up RNA Kit (TransGen Biotech, Beijing, China). The concentration and purity of the RNA were measured using a nucleic acid protein analyzer (IMPLEN). The integrity of the RNA was determined by running an aliquot on a 1% agarose gel. Thereafter, the RNA was used to synthesize cDNA using a reverse transcription kit (TaKaRa, Tokyo, Japan). The housekeeping gene gapdh of A. mellifera was used as an internal reference gene. Quantitative real-time PCR primers were designed based on mRNA sequences acquired from the NCBI database of the vg (vitellogenin LOC406088), hex110 (hexamerin 110 LOC551648), jh (Juvenile hormone, LOC406066), and gapdh (glyceraldehyde-3-phosphate dehydrogenase LOC410122) genes in Primer Premier 5.0 (Premier Biosoft). The primer sequences are listed in Table 1.
The qRT-PCR reaction system (10 μL) consisted of 1 μL of cDNA, 5 μL of TB Green, 3 μL of ddH2O, 0.4 μL each of forward and reverse primers, and 0.2 μL Rox. Amplification conditions were as follows: 50 °C for 2 min, pre-denaturation at 95 °C for 10 min, followed by 40 cycles of 95 °C for 10 s, (vg at 58 °C; hex110 at 56.9 °C; jh at 58 °C; and gapdh at 52.9 °C) for 1 min. For each sample, the specificity of the PCR amplification was verified by melting curve analysis.
Each reaction had four technical replicates for each of the vg, hex110, and jh genes and expression levels were calculated using the 2^−(∆∆Ct)^ method [14]. The expression differences of the vg, hex110, and jh genes were analyzed by one-way ANOVA in SPSS 25.0.
3. Results
3.1. Number of Capped Queen Cells
The results verify that the number of capped queen cells containing 1-day-old larvae was significantly higher than those containing 3-day-old larvae (Independent-Samples Kruskal–Wallis Test, N = 87, p < 0.05). However, there was no difference between the queen cells with 2-day-old larvae and 3-day-old larvae (Independent-Samples Kruskal–Wallis Test, p > 0.05) (Figure 2a). Four-day-old larvae do not have capped queen cells. The statistical analysis results are shown in Supplementary Table S1.
3.2. Number of Newly Emerged Queens
The number of emerging queens reared from 1-day-old larvae was significantly higher than the queens reared from 2-day-old and 3-day-old larvae (Independent-Samples Kruskal–Wallis Test, N = 65, p < 0.05). There was no significant quantitative difference between the number of emerging queens reared from 2-day-old or 3-day-old larvae (Independent-Samples Kruskal–Wallis Test, p > 0.05) (Figure 2b). The statistical analysis results are shown in Supplementary Table S1.
3.3. Quantity Statistics of Newly Emerged Queens on the First Day
On the first day, the number of emerging queens reared from 1-day-old larvae was significantly higher than the queens reared from 2-day-old and 3-day-old larvae (Independent-Samples Kruskal–Wallis Test, N = 37, p < 0.05). There was no significant quantitative difference between the number of emerging queens from 2-day-old and 3-day-old larvae (Independent-Samples Kruskal–Wallis Test, p > 0.05) (Figure 2c).
3.4. Queen Birth Weight
There was no significant difference between the birth weights of queens reared from 1-day-old, 2-day-old, and 3-day-old larvae (ANOVA, df_2,34_, p > 0.05) (Figure 2d).
3.5. Nursing Behavior and Ovariole Numbers of New Queens
Data showed that 1-day-old queen larvae triggered more worker followers than 2-day-old queen larvae (ANOVA, df_2,34_, p < 0.05), but 2-day-old and 3-day-old queen larvae did not attract significantly different numbers of followers (ANOVA, p > 0.05) (Figure 2e).
The number of ovarioles in newly emerged queens from 1-day-old, 2-day-old, and 3-day-old larvae showed no significant difference (Independent-Samples Kruskal–Wallis Test, N = 37, p > 0.05) (Figure 2f).
3.6. Expression of Genes Related to Ovarian Development
The relative expression of the vg gene in queens reared from 1-day-old larvae was significantly higher than that in queens reared from 3-day-old larvae (Independent-Samples Kruskal–Wallis Test, N = 37, p < 0.05) (Figure 3a). In addition, the expression of the hex110 and jh genes in queens developed from 1-day-old larvae was significantly higher than those in queens reared from 2-day-old and 3-day-old larvae (Independent-Samples Kruskal–Wallis Test, N = 37, p < 0.05) (Figure 3b,c).
4. Discussion
Larval age at the time of selection for queen rearing is an important factor affecting queen quality. In this study, in the absence of nepotism, we evaluated queens reared from 1-day-old, 2-day-old, 3-day-old, and 4-day-old larvae. The results confirmed that 3-day-old or younger larvae were selected for queen rearing (Figure 1c), as is widely understood [15,16]. After placing the experimental comb in the queenless colony, we found that the queen cells began to appear in the 1-day-old larvae area, suggesting that 1-day-old larvae are considered queen candidates. Sagili et al. [17] showed that honey bees perceive the nutritional state of larvae and use that information when selecting larvae for rearing queens in the natural emergency queen replacement process. In addition, Margarita and Rangel et al. reported that workers recognize larval hormones to distinguish between the larval grades and developmental stages [18,19], suggesting that both nutrition and hormones may be important in screening queen candidates in the larval stage. Unfortunately, the behavioral aspect of the workers’ selection of queen candidate larvae was not captured. However, recent reports have suggested that learning behavior in bees starts at an early age, and whether it includes judgment is worth considering [20].
It is known that not all the queens from capped queen cells will emerge, as some are destroyed by workers before or after capping [14,21,22,23]. In this study, the worker’s removal of capped and uncapped queen cells started on the sixth day and did not target a particular larval age (Figure 1b,c). This suggests that workers consistently select queen candidates based on quality criteria. Here, the workers cleared the uncapped queen cells because the larvae had stagnated or died, and the same might be true for the capped queen cells (Figure 1d). Morse et al. [24] also found that if the larvae were removed from a capped queen cell, the workers would remove the queen cell. We found that the number of capped queen cells containing 1-day-old larvae was significantly higher than those containing 3-day-old larvae (Figure 2a). Data suggest that workers prefer 1-day-old larvae for filling queen cells, probably because 1-day-old larvae develop into higher quality queens. Rehman et al. [25] used different-aged larvae (12–24, 24–48, and 48–72 h) for grafting, and found that 1-day-old larvae had the highest acceptance rate. Thus, the selection of queen candidates by the colony adheres to the initial selection criteria during the larval stage, but is also a gradual process of optimization.
Interestingly, queens reared from different-aged larvae (1-day-old, 2-day-old, and 3-day-old) all emerged on the first day (Figure 2c), but most of these were from 1-day-old larvae. This may be because queen cells first appeared in the 1-day-old larvae area and these larvae first consumed royal jelly, promoting larval development. Ucak Koc [26] found that the 10-HDA content of royal jelly decreased with increasing larval age in different day-old larvae transferred to the queen table, which supports our findings. This indicates that when colonies screen queen candidates from larvae of different ages, they prioritize the 1-day-old larvae as good quality. However, because of the particularity of the queenless colony, they also select good-quality older larvae (3-day-old larvae) as candidate queens.
During the emerging stage, we placed newly emerged queens into the original colony and allowed nurse bees to feed them. We found that the queens that developed from 1-day-old larvae had more nurse bees. Many studies also found that 1-day-old queen larvae triggered more worker followers than 2-day-old and 3-day-old queen larvae [19,27,28,29]. Thus, an older larval queen is an emergency measure, and the colony will choose the most suitable queen for the growth of the colony.
Various studies have shown that the reproductive potential of queens can vary quite considerably and can be measured in their size, number of ovaries, and weight [16,30,31,32,33,34,35]. The expression of the vg, hex110, and Jh genes can also be used to determine the quality of queens, as they are related to physiological indicators and ovarian activation [36,37,38]. Interestingly, there was no significant difference in the birth weights and number of ovarioles of queens reared from 1-day-old, 2-day-old, and 3-day-old larvae (Figure 2d,f), but our results showed that the relative expression levels of the vg, hex110, and Jh genes were higher in queens reared from 1-day-old larvae than those reared from 3-day-old larvae. This suggests that the quality of the queens reared from 1-day-old larvae was superior (Figure 3).
5. Conclusions
While the exact mechanism by which workers assess queen quality is unknown, the result of this layered filtering of investments in queen candidates may be to maximize colony development. In the absence of nepotism, the selection of queen candidates by workers is not final but is a gradually optimized process. Thus, workers can play an important role in the quality control of the queen selection process, which is highlighted by the results of this study.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Huber F. Nouvelles Observations sur les Abeilles, I and II (New Observation on Bees, I and II)Create Space Independent Publishing Platform Scotts Valley, CA, USA 1926
- 2Ribbands C.R. The Behaviour and Social Life of Honeybees Research Bees London, UK 1953
- 3Crozier R.H. Pamilo P. Genetic Intrigues. (Book Reviews: Evolution of Social Insect Colonies. Sex Allocation and Kin Selection.)Science 199627414771478
- 4Page R.E.Jr. Sperm utilization in social insects Annu. Rev. Entomol.19863129732010.1146/annurev.en.31.010186.001501 · doi ↗
- 5Strassmann J. The rarity of multiple mating by females in the social Hymenoptera Insectes Sociaux 20014811310.1007/PL 00001737 · doi ↗
- 6Meixner M. Moritz R. Clique formation of super-sister honeybee workers (Apis mellifera) in experimental groups Insectes Sociaux 200451434710.1007/s 00040-003-0701-5 · doi ↗
- 7Getz W.M. Smith K.B. Genetic kin recognition: Honey bees discriminate between full and half sisters Nature 198330214714810.1038/302147 a 0 · doi ↗
- 8Makert G.R. Paxton R.J. Hartfelder K. Ovariole number—A predictor of differential reproductive success among worker subfamilies in queenless honeybee (Apis mellifera L.) colonies Behav. Ecol. Sociobiol.20066081582510.1007/s 00265-006-0225-x · doi ↗