Evolutionary reversal of physical dormancy to nondormancy: evidence from comparative seed morphoanatomy of Argyreia species (Convolvulaceae)
D M Nethani H Gunadasa, K M G Gehan Jayasuriya, Jerry M Baskin, Carol C Baskin

TL;DR
This study shows that some Argyreia plant species evolved from producing dormant seeds to non-dormant seeds, adapting to their wet habitats.
Contribution
The paper provides evidence of an evolutionary reversal from physical dormancy to non-dormancy in Argyreia seeds.
Findings
Three Argyreia species have non-dormant seeds, while two retain physical dormancy.
Non-dormant seeds are found in species with restricted geographic distribution.
Anatomical remnants suggest non-dormant seeds evolved from physically dormant ones.
Abstract
Argyreia is the most recently evolved genus in the Convolvulaceae, and available information suggests that most species in this family produce seeds with physical dormancy (PY). Our aim was to understand the evolution of seed dormancy in this family via an investigation of dormancy, storage behaviour, morphology and anatomy of seeds of five Argyreia species from Sri Lanka. Imbibition, germination and dye tracking of fresh intact and manually scarified seeds were studied. Scanning electron micrographs and hand sections of the hilar area and the seed coat away from the hilar area were compared. Scarified and intact seeds of A. kleiniana, A. hirsuta and A. zeylanica imbibed water and germinated to a high percentage, but only scarified seeds of A. nervosa and A. osyrensis did so. Thus, seeds of the three former species are non-dormant (ND), while those of the latter two have physical…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
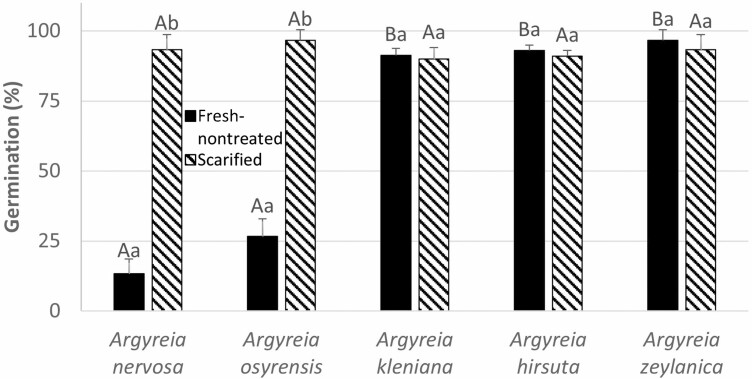 Figure 1
Figure 1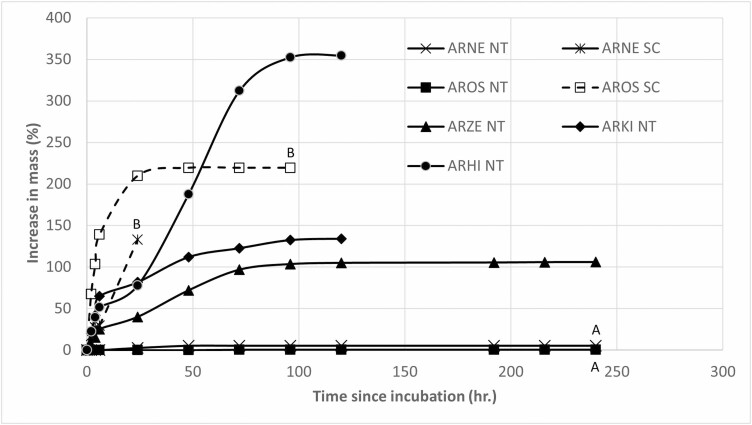 Figure 2
Figure 2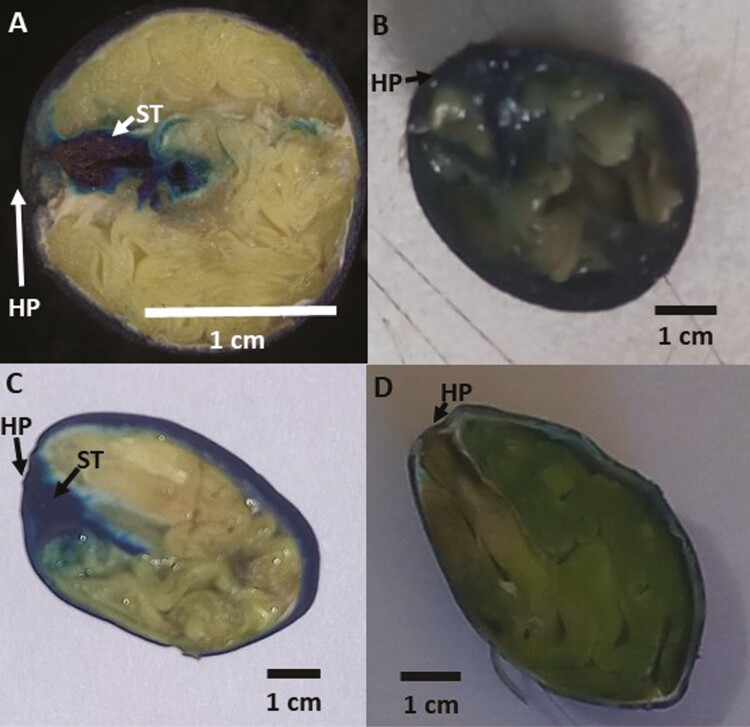 Figure 3
Figure 3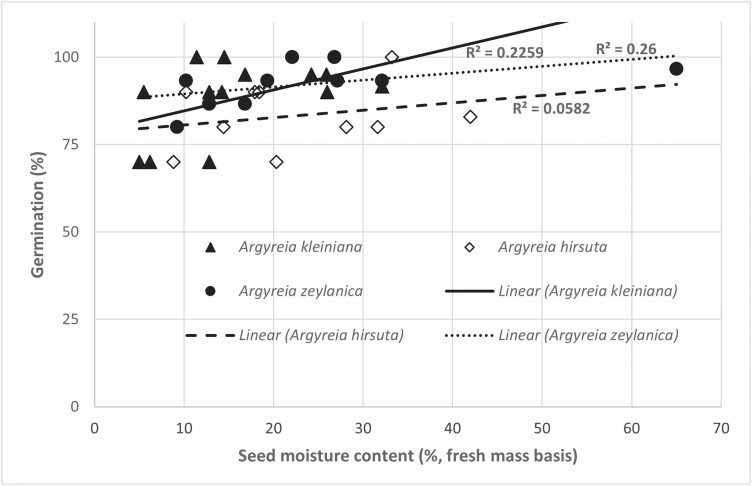 Figure 4
Figure 4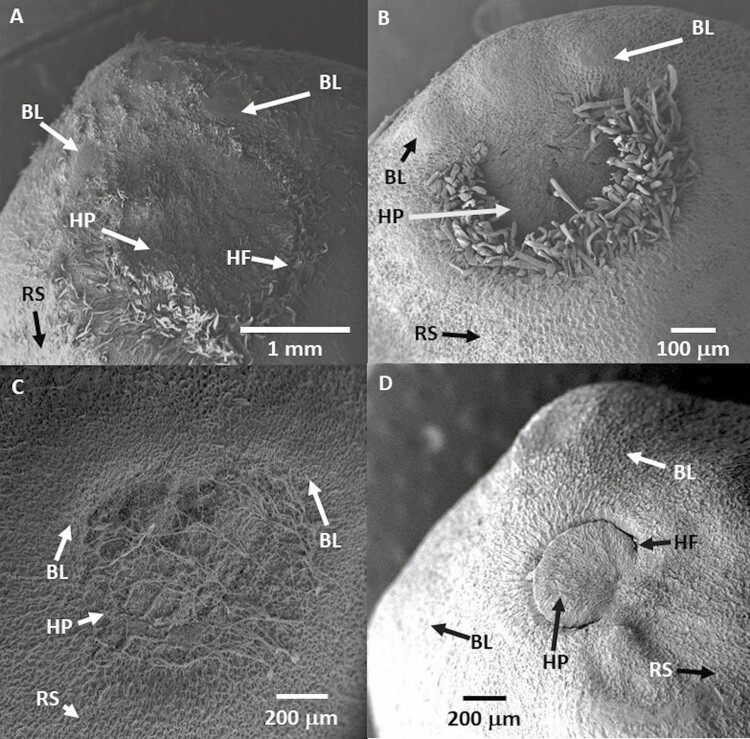 Figure 5
Figure 5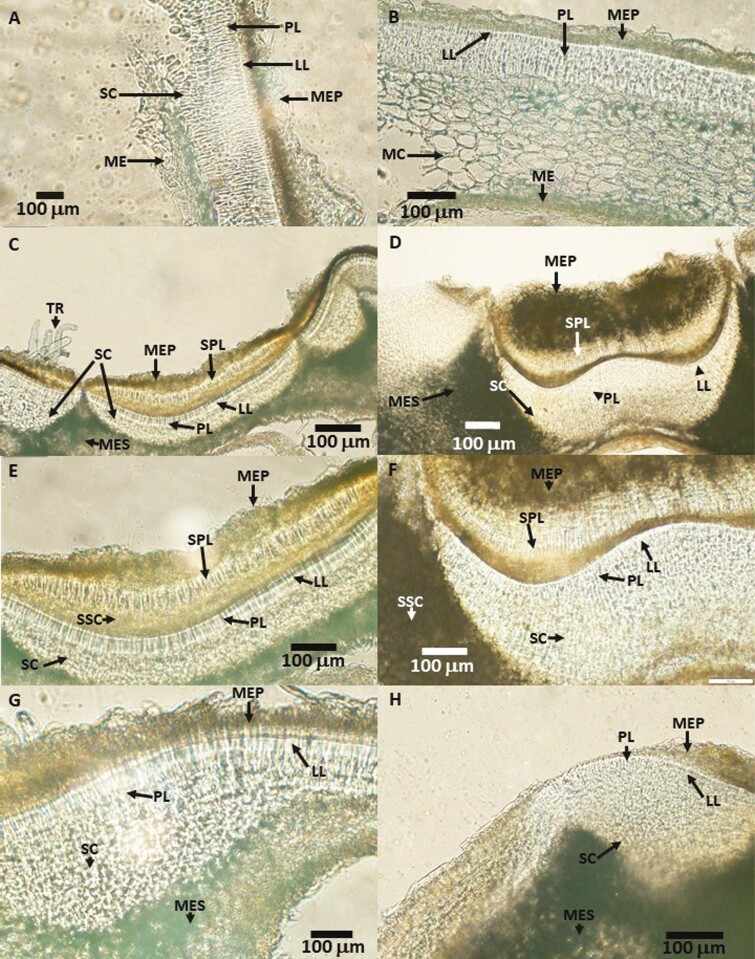 Figure 6
Figure 6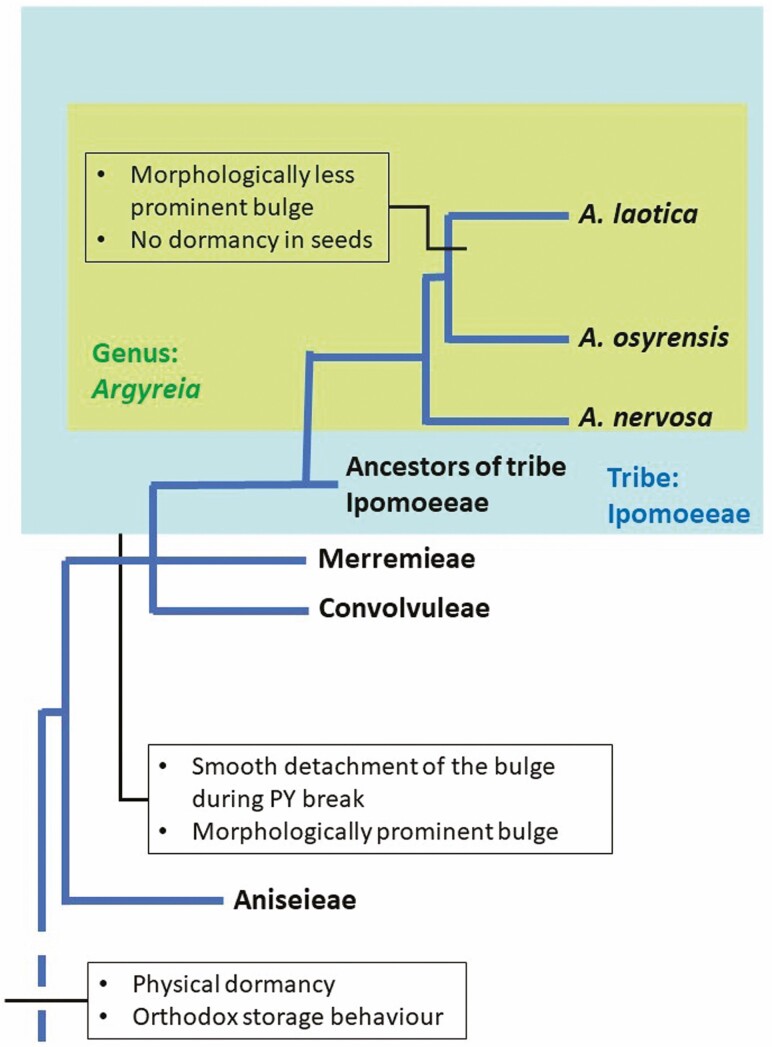 Figure 7
Figure 7| Species | Seed collection | Rainy season of the habitat (Survey Department of Sri Lanka, 2012) | |
|---|---|---|---|
| Site | Time | ||
|
| Seeds were purchased | ||
|
| From a secondary forest in Bowatenna | June, 2022 | October to February |
|
| Roadside in Peradeniya | May, 2022 | April to December |
|
| Roadside in Riverston, Knuckles forest reserve | October, 2022 | April to December |
|
| Secondary forest in Rambakan oya, Amapara | November, 2022 | October to February |
| Study species | Mean (± SE) fresh mass per seed (g) | Mean (± SE) dry mass per seed | Mean (± SE) seed moisture content (%) |
|---|---|---|---|
|
| 0.10 ± 0.02 | 0.091 ± 0.02 | 13.1 ± 1.4a |
|
| 0.015 ± 0.001 | 0.012 ± 0.001 | 12.7 ± 0.7a |
|
| 0.14 ± 0.03 | 0.09 ± 0.02 | 32.1 ± 4.2b |
|
| 0.45 ± 0.05 | 0.26 ± 0.04 | 42.0 ± 5.8c |
|
| 0.32 ± 0.05 | 0.12 ± 0.03 | 64.1 ± 4.4d |
| Current name | Synonym | Native/exotic | Distribution | Ecology | Fruit type | Dormancy class | |
|---|---|---|---|---|---|---|---|
| Species in Sri Lanka |
|
| Native (Extinct from SL) | Sri Lanka, India | Dry zone | No record | Could not infer |
|
| Native | Sothern India, Central Sri Lanka | Montane wet zone >2000 feet | Fruits fleshy, indehiscent | ND | ||
|
| Endemic | Sri Lanka | Submontane zone | Fruits fleshy, indehiscent | ND | ||
|
| Native | Central Sri Lanka, Nilagiri mountains India | Submontane zone | Fruits baccate, fleshy | ND | ||
|
| Exotic | Assam and Bengal to Mysore, Sri Lanka (as naturalized exotic) | Open woodlands, roadsides, disturbed sites and waste areas | Dry, dehiscent | PY | ||
|
| Native | Cambodia, Hainan, India, Laos, Myanmar, Sri Lanka, Sumatra, Thailand, Vietnam | Disturbed sites in the dry zone of Sri Lanka | Dry, dehiscent | PY | ||
|
|
| Native | Kerala, India and Sri Lanka | Dry zone of Sri Lanka | Fruits baccate, fleshy | ND | |
|
|
| Native | Kerala, India and Sri Lanka | Wet and dry zones in Sri Lanka | Fruits baccate, fleshy | ND | |
|
|
| Native | India, Burma, Sri Lanka | Montane wet zone > 2000 feet | Fruits baccate, fleshy | ND | |
|
| Endemic | Sri Lanka | Wet zone Lowland | Fruits baccate, fleshy | ND | ||
| Species not in Sri Lanka |
|
| China, Cambodia, India (Andaman Islands), Indonesia, Laos, Malaysia, Myanmar, Thailand, Vietnam | Dense forests wet zone in intermediate elavations | Fruits red or orange berry | ND | |
|
| China, Cambodia, NE India, Indonesia, Laos, Malaysia, Myanmar, Thailand, Vietnam | Open waste ground in wet zone | Fruits berry | ND |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSeed Germination and Physiology · Allelopathy and phytotoxic interactions · Plant and animal studies
Introduction
Physical dormancy (PY) in seeds, is caused by seed/fruit coat water impermeability (Rolston 1978; Baskin et al. 2000), and it has been identified in 18 angiosperm families (Mahadevan and Jayasuriya 2013; Baskin and Baskin 2014). Convolvulaceae is the only family with species producing physically dormant seeds in the taxonomically most recently evolved Asterid clade (Baskin et al. 2000). However, the Convolvulaceae not only includes species that produce seeds with PY but also those that produce seeds with combinational dormancy (i.e. physical + physiological dormancy) and with no dormancy (Jayasuriya et al. 2008).
Jayasuriya et al. (2008) have determined the kind of dormancy, germination requirements and storage behaviour of 46 species representing all 11 tribes of the subfamily Convolvuloideae (sensu Stefanovic et al. 2002). They found PY and combinational (PY + PD) dormancy and non-dormancy within the family. Further, orthodox and recalcitrant storage behaviour types were present within the family. Orthodox seeds are desiccation tolerant and have high storability, while recalcitrant seeds are desiccation sensitive and have low storability (Hong and Ellis 1996). Further, Jayasuriya et al. (2009) investigated the seed coat and ‘water gap’ morphology and anatomy of these species and described the evolutionary trends in seed dormancy within the subfamily Convolvuloideae. They showed that species in tribe Erycibeae, which has a tropical rainforest origin, have non-dormant (ND) recalcitrant seeds, while species in tribes Chardiochlaymeae and Cuscuteae in seasonal tropical climates have orthodox seeds with PY. Further, although most species in tribe Cresseae have seeds with PY, some members of Cresseae growing in wet aseasonal tropical climates have ND seeds, e.g. Bonamia menziesii (Jayasuriya et al. 2009). Moreover, members of the tribe Maripeae (with a wet tropical distribution) have ND seeds, while most members of sister tribes [Jacquemontieae, Cresseae and Dichondreae (Stefanovic et al. 2003)] have seeds with PY (Jayasuriya et al. 2009).
Ipomoeeae, the most recently evolved tribe in the Convolvulaceae, has a high number of species with a wide distribution in tropical and temperate habitats (Stefanovic et al. 2003; Mabberley 2017). Despite this wide geographical distribution and a high number of species, all species in Ipomoeeae studied thus far have seeds with PY (Baskin and Baskin 2014). Gupta (2003) suggested that Argyreia nervosa seeds might have PY or ND. Stefanovic et al. (2003) included Argyreia, Binkworthia, Lepistemon, Lepistermonopsis, Paralepistemon, Rivea, Stictocardia and Turbina in tribe Ipomoeeae based on molecular data. However, Austin (1973, 1998) and Wilkin (1999) included these genera in a separate tribe (Argyreieae) based on morphological, anatomical and physiological data. Moreover, Stafnovic et al. (2002) identified Argyreia and Rivea as forming a monophyletic group, but these two genera are well nested within the cluster formed by other Ipomoeeae genera. Seed dormancy and anatomy of tribe Argyreieae (sensuAustin 1998) have not been studied in detail; this tribe was not included in the studies by Jayasuriya et al. (2008, 2009) on seed dormancy/anatomy of the Convolvulaceae. Thus, the possibility that tribe Argyreieae may contain some species with ND seeds and others with physically dormant seeds makes this an important tribe with which to further explore the evolutionary relationship between ND and PY in the Convolvulaceae.
To contribute or our understanding of the evolution of seed dormancy in the family Convolvulaceae, we investigated the dormancy, morphology and anatomy of seeds of five Argyreia species, which represent the most recently split-off sub-tribe of the family: Argyreia nervosa (Burm. f.) Bojer, Argyreia kleiniana (Schult.) Raizada, Argyreia hirsuta Wight & Arn.. Argyreia osyrensis (Roth) Choisy and A. zeylanica (Gaertn.) Voigt. Argyreia has a paleotropical origin and distribution (Eserman et al. 2014), and it occurs in South and Southeast Asia and Madagascar (IPNI 2022). We hypothesized that seeds of Argyreia species occurring in aseasonal habitats such as tropical rainforests have ND seeds, while sister species growing in seasonal habitats have seeds with PY.
Methods and Materials
Study species
All five study species of Argyreia are perennial lianas. A. nervosa has a wide native distribution in Asam, Nepal, Bangladesh, India and Myanmar, and it has been introduced and naturalized in many other Asian (including Sri Lanka) and African countries (IPNI 2022). This species occurs in rainforests, open woodlands, waste ground, riverbanks and roadsides of wet habitats up to 900 m a.s.l. (Padhi et al. 2013). A. osyrensis occurs in Sri Lanka and in southern India to Burma, and it grows in disturbed sites in the dry zone (Austin, 1980). A. kleiniana is native to Sri Lanka and the Western Ghats of India, and it is found in rainforests, open woodlands, waste grounds and roadsides of lowland wet habitats (Austin 1980). A. hirsuta is found only in the Nilagiri Mountains of India and the hill country of Sri Lanka. Its habitats are the same as those of A. kleiniana, but the species occurs only above 600 m a.s.l. (Austin 1980). A. zeylanica is found only in southern India and northeastern Sri Lanka in the dry zone. It occurs in disturbed sites similar to those of other species.
Fruit collection and seed extraction
Cleaned non-treated seeds of A. nervosa were purchased from Nurserylive, Magarpatha, Maharashtra, India. Fruits of A. osyrensis, A. kleiniana, A. hirtusa and A. zeylanica were collected from at least five lianas in Bowatenna (dry zone), University of Peradeniya, Peradeniya (Wet zone), Knuckles conservation area (Intermediate zone) and Rambaken-Oya nature reserve, Ampara, Sri Lanka, respectively, during their peak dispersal time (Table 1). All the fruits were collected from plants (and not from the ground) and care was taken to collect only (yellow-coloured soft mature fleshy fruits at dispersal maturity vs. green-coloured hard non-matured fruits of A. kleiniana, A. hirtusa and A. zeylanica and red-coloured dry mature fruits vs. green-coloured freshy nonmature fruits of A. osyrensis). Collected fruits were placed in brown paper bags and transported to the seed biology laboratory at the University of Peradeniya. Fleshy fruits of A. kleiniana, A. hirtusa and A. zeylanica were immersed in water to separate seeds. Seeds were washed thoroughly to remove debris and air dried for 3 hr to remove surface water, after which they were stored in plastic bottles until used for laboratory experiments. Seeds of A. osyrensis were removed from the dry fruits by hand. Laboratory experiments were initiated for the four species collected from the wild within 3 days after collection, while those for A. nervosa were initiated within 3 days from the reception of the samples. Moreover, seeds of A. nervosa were collected about 1 month prior to shipping, and shipping the material from India to Sri Lanka took 8 days. A. nervosa seeds were stored in plastic bottles at ambient room temperature conditions (~ 32 ^o^C, 55% RH).
Seed dormancy and germination
Experiments were conducted to determine if seeds of the five study species had PY. If seeds have PY, manually scarified seeds imbibe water and germinate within a few days, while non-treated intact seeds do not. If nontreated intact seeds imbibe water and germinate within a few days, they are ND.
Seed germination.
Four replicates of 15 non-treated intact seeds of A. nervosa, A. osyresnsis and A. zeylanica and 25 non-treated seeds each of A. kleiniana, and A. hirtusa were placed on tissue papers moistened with distilled water in 9-cm-diameter Petri dishes. The number of seeds used per replicate differed depending on the availability of seeds. Seeds were incubated at laboratory temperature (~25 ^o^C) and light (diffused sunlight and white fluorescent light during the daytime and complete darkness during the night [12 hr]) conditions. Seeds were observed for germination and signs of imbibition (seed coat rupture and swelling of seeds) at 2-day intervals for 30 days or until all seeds had germinated. Emergence of the radicle to a length of ~1 mm was the criterion for germination.
Imbibition.
Ten manually scarified (with a single-edge razor blade) and 10 non-treated intact seeds each of A. nervosa were placed on tissue papers moistened with distilled water. Each seed was retrieved after 0, 2, 4, 6 and 24 hr of incubation and then at 1-day intervals for 10 days or until all seeds had germinated, blotted dry, reweighed and returned to the Petri dishes. The same experiment was conducted for A. osyrensis using 25 intact non-treated and 25 manually scarified seeds and for A. kleiniana, A. hirtusa and A. zeylanica using 15 intact non-treated seeds. Imbibition curves were constructed and compared.
Dye tracking of pathway of water entry into seeds.
Dye tracking was conducted to determine the potential pathway of water entry into the seeds of A. kleiniana, A. hirsute, A. nervosa and A. osyrensis. Five non-treated intact seeds each of A. hirsuta A. kleiniana, A. nervosa and A. osyrensis were immersed in a saturated methylene-blue solution. Seeds were retrieved after 1, 2, 4, 6 and 24 hr of immersion, cut into halves and observed under a dissecting microscope. The path of dye entry (or not) into the seed was stained blue. This experiment was not conducted for A. zeylanica.
Seed moisture content
Moisture content of seeds was determined as soon as seeds were extracted from fruits i.e. as soon as seeds were brought to the seed biology laboratory. Thus, the measured seed moisture content of five study species except for A. nervosa can be considered as the moisture content of fresh seeds. Five replicates of three halved seeds each of A. nervosa and A. osyrensis and five replicates of 15 halved seeds each of A. hirsuta, A. kleiniana and A. zeylanica were weighed with a digital analytical balance to the nearest 0.001 g. Seeds were oven dried at 120 ^o^C for 3 hr and reweighed. SMC was calculated on a fresh mass basis using the following equation.
Effect of desiccation on seed viability and seed desiccation sensitivity
Experiments were conducted to determine the desiccation sensitivity/tolerance of seeds of the five study species. Seeds that can survive <10% seed moisture content (SMC) were categorized as desiccation tolerant, and those that lose viability when dried to <10% SMC were categorized as desiccation sensitive.
This experiment was conducted only for seeds of A. hirsuta, A. kleiniana and A. zeylanica. Five samples containing three replicates of 15 seeds (5 samples × 3 replicates × 15 seeds for each species) each of the species were weighed and placed in open Petri dishes in a desiccator. Seed samples were retrieved at 1-day intervals and reweighed. When seeds attained ~10, 20 and 30 % SMC, a sample (three replicates) was placed on tissue papers moistened with distilled water in Petri dishes and incubated at ambient laboratory temperature and light conditions as described above. Seeds were observed for germination at 1-day intervals until all seeds had germinated or rotted. Percentage viability of seeds was determined based on seed germination percentage.
Seed morphology
Seeds of each of the study species were fixed to metal plates with double gum tape and sputter coated with gold/palladium mixture. Scanning electron micrographs of the hilum and bulge area were taken with a Zeiss EVO LS15 high-performance variable pressure environmental scanning electron microscope. This experiment was not conducted for A. zeylanica.
Seed anatomy
Transverse sections of the seed coat at the hilum, bulge and seed coat away from the hilum of A. osyrensis and A. kleiniana were made using a single-edge razor blade. Sections were mounted on glass slides and observed and micrographed using an Olympus DP74 camera connected to the Olympus BX53 light microscope.
Analysis of data
For all five species, germination data were analysed with the non-parametric Kruskal–Wallis test since the data did not follow a normal distribution. For A. nervosa and A. osyrensis, the final imbibition percentages of nontreated and scarified seeds were analysed separately with a Mann–Whitney test. Linear regressions were fitted separately for viability and moisture content of A. kliniana, A. hirsuta and A. zeylanica seeds to determine whether viability was reduced with decreasing moisture content.
Results
Seed dormancy and germination
Germination.
Fresh intact seeds of A. nervosa and A. osyrensis germinated to 13.3% and 26.7%, respectively, within 30 days, while those of A. kleiniana, A. hirsuta and A. zeylanica germinated to >90% (Fig. 1). Germination percentages of the five study species differed significantly ((H[χ^2^] = 25.8, P = 0.001). Manually scarified seeds of A. nervosa and A. osyrensis germinated to 93.3 ± 5.4 and 96.7 ± 3.8%, respectively, while the few remaining seeds rotted. Non-scarified seeds of A. nervosa and A. osyrensis that did not germinate within 30 days did not show any sign of imbibition, while scarified seeds and germinated nonscarified seeds increased in size and their seed coats were ruptured before germination.
Germination percentage of fresh intact seeds of Argyreia hirsuta, A. kleiniana A. nervosa, A. osyrensis and A. zeylanica at ambient laboratory temperature (~25 oC) and light/dark conditions. Different lowercase letters indicate significant differences between scarified and nonscarified seeds within the same species and different uppercase letters significant differences between species within the same treatment. Error bars are + 1 SE.
Imbibition.
Increase in mass of intact nontreated seeds of A. kleiniana, A. hirsuta and A. zeylanica seeds was > 100 % (Fig. 2), while that of intact nontreated seeds of A. nervosa and A. osyrensis was <10%. In contrast, scarified seeds of these two species had a mass increase of >100%. Mass increase percentages of intact nontreated seeds of A. nervosa (U = 3.74, P < 0.001) and A. osyrensis (U = 4.72, P < 0.001) were significantly lower than that of scarified seeds.
Mass increase (percentage) of intact and scarified seeds of Argyreia nervosa and A. osyrensis and of intact seeds of A. hirsuta, A. kleiniana and A. zeylanica during imbibition at ambient laboratory temperature (~25 oC) and light/dark conditions. ARHI NT, Argyreia hirsuta nontreated intact; ARKL NT, Argyreia kleiniana nontreated intact; ARNE NT, Argyreia nervosa nontreated intact; ARNE SC, Argyreia nervosa scarified; AROS NT, Argyreia osyrensis nontreated intact; AROS SC, Argyreia osyrensis scarified; and ARZE NT, A. zeylanica nontreated intact. Different uppercase letters indicate significant differences between the final mass increase of scarified vs. intact seeds of the same species (AROS NT vs. AROS SC and ARNE NT vs. ARNE SC).
Dye tracking of pathway of water entry into seeds.
No staining was observed inside any of the seeds after 1 hr of soaking in methylene blue. However, after 2 hr blue staining was observed below the hilum pad in the seeds of A. kleiniana (Fig. 3A) and A. hirsuta (Fig. 3C). After 4 hr, the whole inside of A. kleiniana (Fig. 3B) and A. hirsuta (Fig. 3 D) seeds was stained. Even after 24 hr, no staining was observed inside the seeds of A. nervosa or A. osyrensis (data not shown).
Seeds of A. kleiniana after 2 hr (A) and after 24 hr (B) and of A. hirsuta after 2 hr (C) and after 24 hr (D) of methylene blue staining. HP, hilar pad; ST, methylene blue dark staining.
Seed moisture content
A. hirsuta had the highest fresh seed mass and A. osyrensis the lowest (Table 2). The same trend was observed in seed dry mass. A. nervosa and A. osyrensis had an SMC < 15%, and the other three species had an SMC > 30%, at dispersal. SMC of the five study species differed significantly (H[χ^2^] = 35.8, P < 0.001).
Effect of drying on seed germination and seed desiccation sensitivity
Within about 3 days, SMC of A. hirsuta, A. kleiniana and A. zeylanica was <10%, and there were no significant differences in germination (viability) percentage for seeds dried to different seed moisture contents (A. kleiniana: H[χ^2^] = 4.06, P = 0.23, A. hirsuta; H[χ^2^] = 1.07, P = 0.57, A. zeylanica; H[χ^2^] = 3.98, P = 0.21) (Fig. 4). Although there were positive linear trends in germination, they did not differ significantly from a parallel line (R^2^ = 0.22, P = 0.10 [A. kleiniana], R^2^ = 0.06, P = 0.50 [A. hirsuta] and R^2^ = 0.26, P = 0.14 [A. zeylanica]).
Viability of Argyreia hirsuta, A. kleiniana and A. zeylanica seeds dried in a silica gel desiccator to different seed moisture contents. Linear regression lines were fitted to show the trend in change of the viability.
Seed morphology
There were similarities as well as differences in the morphology of seeds of the five study species (Fig. 5). Seeds of the four species (A. kleiniana, A. hirsuta, A. osyrensis and A. nervosa) have a hilum pad and bulges [water gap in seeds of Convolvulaceae (Jayasuriya et al. 2009)]. The whole surface of A. nervosa seeds has trichomes (Fig. 5A), while trichomes were observed only on the hilum pad of A. kleiniana and around the hilum pad of the A. osyrensis seeds (Fig. 5C). The hilum pad of A. kleiniana has short trichomes, which normally were not visible to the naked eye. In contrast, trichomes on A. nervosa seeds are not densely arranged. They were visible to the naked eye and longer than those of A. kleiniana. A. hirsuta and A. zeylanica seeds did not have trichomes. Bulges on A. osyrensis seeds were more prominent than those on seeds of A. kleiniana, A. hirsuta and A. nervosa. On the other hand, A. nervosa seeds also have more prominent bulges than those of A. kleiniana, A. hirsuta and A. zeylanica. The hilum pad and the seed coat away from the hilar area of A. nervosa, A. hirsuta and A. zeylanica were separated by a clear hilum fissure, but there was no obvious hilum fissure on A. kleiniana or A. osyrensis seeds.
Electron micrographs of Argyreia nervosa (A), A. osyrensis (B) A. kleiniana (C) and A. hirsuta (D) seeds showing the hilum and bulge area. BL, bulge; HF, hilum fissure; HP, hilum pad; RS, seed coat away from hilum.
Seed anatomy
The anatomy of the seed coat away from the hilum area of A. osyrensis and A. kleiniana seeds was similar to that of a typical water-impermeable seed coat. The outermost layer of the seed coat of both species consists of macerated cells (Fig. 6A and B). Below the macerated cells, both species have a palisade layer with a distinct light line followed by several layers of sclerenchyma cells. A slight difference in seed coat anatomy could be observed in this layer, i.e. A. osyrensis has 4–5 sclerenchyma layers, while A. kleiniana has only 2–3 sclerenchyma layers. Below the sclerenchyma layer, the A. osyrensis seed coat consists of a macerated cell layer, which is probably the remnants of the nucellus or the megagametophyte, and the A. kleiniana seed coat has a living mesophyll cell layer between the macerated cell and the sclerenchyma cell layers.
Micrographs of hand sections of Argyreia osyrensis (A, C, E, G) and Argyreia kleiniana (B, D, F, H) showing seed coat away from the hilum (A, B), hilum and hilum fissure (B, C), hilum (E, F) and the bulge (G, H) area. LL, light line; MC, mesophyll cell layer; ME, macerated cell layer below the seed coat; MEP, macerated cell layer above the seed coat; PL, palisade layer; SC, sclerenchyma cell layer; SPL, second palisade layer; SSC, second sclerenchyma layer.
The anatomy of the hilar area of A. osyrensis and A. kleiniana is different from that of the seed coat away from the hilar area (Fig. 6C and D). The hilar pad of A. osyrensis is surrounded by a sub-hilar area, which is separated from the hilar pad by the hilar fissure. However, the sub-hilar area is not present on the micropylar side of the seed. The outermost layer of the sub-hilar area consists of trichomes and macerated cells and then of anatomical layers that are similar to those of the seed coat away from the hilar area. However, the sclerenchyma area consists of more layers than that of the seed coat away from the hilar area. The sub-hilar area of the A. kleiniana seed is not as conspicuous as it is in A. osyrensis.
The hilar pad of the seeds of A. osyrensis and A. kleiniana is very similar (Fig. 6C–F). The outermost layer of the hilar pad on seeds of both species consists of macerated cells. A palisade layer without a light line can be observed below the layer of macerated cells. A layer of sclerenchyma cells is below the palisade layer. The palisade layer with the light line, which is comparable to the palisade layer of the seed coat away from the hilar area, occurs below the sclerenchyma layer. A second sclerenchyma layer is present below the palisade layer with the light line. Below the second sclerenchyma cell layer is a mesophyll cell layer, which may be cells remaining from the nucellus.
Seeds of A. osyrensis and A. kleiniana have two bulges slightly above the hilar area. The seed coat of the bulges is thicker than that of the seed coat away from the hilum area (Fig. 6G, H). The thickness of the sclerenchyma layer in the bulges is responsible for the increased thickness of the seed coat on the bulges.
There are no clear differences between the two species in the anatomy of seed coat away from the hilum, hilum area and bulges.
Discussion
Imbibition and germination experiments clearly showed that seeds of A. kleiniana A. hirsuta and A. zeylanica are ND, since >95% of intact non-treated seeds imbibed water and germinated. Since seeds of A. nervosa and A. osyrensis germinated <30% and manually scarified seeds imbibed significantly higher amounts of water than the nonscarified seeds, they have PY. On the other hand, >90% survival during the desiccation of seeds of A. kleiniana, A. hirtusa and A. zeylanica, and high viability (>90%) at the dispersal moisture content (~12%) of A. nervosa and A. osyrensis revealed that they were desiccation tolerant. Thus, seeds of A. kleiniana, A. hirsuta and A. zeylanica which have a restricted distribution in Southern India and Sri Lanka (Austin 1980) produce desiccation-tolerant ND seeds, while A. nervosa and A. osyrensis with a wide distribution in the dry zone of India and Sri Lanka (Austin 1980) produce desiccation-tolerant physically dormant seeds.
Seed dormancy and desiccation sensitivity/tolerance are two important seed traits that determine the timing of germination (Rubio de Casas et al. 2012), and thus they are highly influenced by the environmental conditions in the habitat of a plant species. A. kleiniana and A. hirtusa are distributed strictly in high-humidity habitats where they are exposed to high pathogenic and predatory pressures (Vazquez-Yanes and Orozco Segovia 1984). As an escape strategy from these pressures, seeds of species in humid habitats tend to germinate faster than those in dry habitats (Dalling et al. 2011, 2020). Thus, it may be advantageous for the ND seeds of A. kleiniana and A. hirtusa to germinate quickly under the humid conditions of their habitat. In contrast, ND seed-producing A. zeylanica occurs in the dry zone of southern India and northeastern Sri Lanka. Nevertheless, this species disperses seeds at the beginning of the wet season (December, personal observation by KMGGJ), and thus fast germination may allow them to escape the high pathogenicity during the wet season of the dry zone. A. nervosa occurs in a wide range of habitats including seasonally dry ones (Padhi et al. 2013), and A. osyrensis occurs mainly in dry zone habitats including dry mixed evergreen forest fringe and disturbed secondary scrub in the dry zone (Austin 1980). As such, PY of A. nervosa and A. osyrensis seeds may be an adaptation to their habitats that synchronizes germination to a time favourable for seedlings to develop, i.e. the wet season. These two species occur in rather dry habitats and probably do not germinate during intermittent rains in the dry season because the seeds are dormant. Notably, the natural dormancy-breaking cues for A. nervosa and A. osyrensis seeds have not been determined. On the other hand, physical dormancy is also considered an adaptation to defend against seed predators (Paulsen et al. 2013; but see Jayasuriy et al. 2015) and pathogens (Dalling et al. 2020).
Argyreia is among the most recently split-off genera in the Convolvulaceae, and Argyreia and Revia form a monophyletic group well nested within the tribe Ipomoeeae (Stafnovic et al. 2002). All Ipomoea species and all other studied species in tribe Ipomoeeae are reported to have seeds with PY. Thus, PY can be considered to be the ancestral dormancy state in the genus Argyreia, and ND in seeds of A. kleiniana, A. hirsuta and A. zeylanica can be considered to be a derived character. Therefore, the evolution of ND from PY within the clade Argyreiae could be a result of the movement of Argyreia from dry to wet aseasonal habitats. However, Argyreia seeds have retained their ability to tolerate desiccation, although recalcitrancy is considered to be an adaptive advantage in wet aseasonal habitats (Pammenter and Berjak 1999).
The evolution of ND from PY in the genus Argyreia also is supported by the comparative anatomy and morphology of the seeds of A. kleiniana (ND) and A. osyrensis (PY). Seeds of both species have similar morpho-anatomy, including a conspicuous hilum area and a bulge similar to those in seeds of other Ipomoeeae species. Although the bulge of the ND A. kleiniana seeds is functionally not active (as a water gap), it is morphologically and anatomically distinguished from the rest of the seed coat and is similar to the (probably) functional bulge of A. osyrensis seeds that is the water gap of Convolvulaceae seeds with PY (as identified by Jayasuriya et al. 2009). Furthermore, the seed coat away from the bulge and the hilum of A. kleiniana seeds has anatomy similar to that of seed coats of Convolvulaceae seeds with PY, including those of A. osyrensis. The seed coat of ND seeds of A. kleiniana contains a palisade layer with a light line, a specific feature of seeds with PY (see Baskin and Baskin 2022), which likely is a relic of ancestors whose seeds had PY.
Among the five species studied, A. nervosa and A. osyrensis produce seeds with PY as well as dry dehiscent fruits, while A. kleiniana, A. hirsuta and A. zeylanica produce ND seeds and fleshy indehiscent fruits. Dry dehiscent fruits of Convolvulaceae generally are explosively dispersed, and seeds may be dispersed (primary dispersal) for a relatively short distance away from the mother plant. It is advantageous for these seeds to be dormant because it gives them time to be secondarily dispersed (Athugala et al. 2021). On the other hand, fleshy fruits are adapted for dispersal by frugivores. Thus, the seeds potentially can be dispersed for long distances and to specific sites suitable for germination, e.g. aseasonal habitats, or in the wet season of seasonal habitats where germination and seedling establishment can occur successfully without delay. Further, PY cannot develop in seeds in fleshy fruits because they require maturation drying to a species-specific low moisture content of ~8–13% to do so (Qu et al. 2010; Baskin and Baskin 2014; Jaganathan 2016, 2022). We speculate that the evolution of ND from PY and of fleshy fruits from dry dehiscent fruits may have occurred simultaneously. The dye-tracking experiment showed that water uptake (imbibition) in both A. kleiniana and A. hirsuta seeds is mainly through the hilar fissure. Moreover, even if the A. kleiniana seeds were desiccated they absorbed water through the hilar fissure (personal observations by first author).
There are 10 Argyreia spp. in Sri Lanka, and except for A. nervosa all of them are native to this country (Austin, 1980). A. nervosa was introduced from India to Sri Lanka as well as to many other countries. Among the nine species native to Sri Lanka, A. hancorniaefolia and A. thwaitesii are endemic to Sri Lanka and four species, A. elliptica, A. hirsuta, A. zeylanica and A. kleiniana, are restricted to southern India and Sri Lanka. A. nervosa A. osyrensis and A. laotica have a wider geographical distribution, occurring in peninsular India, Sri Lanka and other neighbouring countries like Burma. According to the descriptions given by Austin (1980; Table 3), all of the Argyreia species, except A. nervosa and A. osyrensis have indehiscent fleshy fruits. Thus, we can speculate that these species also produce ND seeds. Fruits of A. nervosa and A. osyrensis are dry and dehiscent, and both species produce seeds with PY. According to our study and speculations made based on the fruit type, species with a wide distribution have PY seeds (except A. laotica), while species with a restricted distribution have ND seeds.
In the phylogenetic analysis by Stafanovic et al. (2002), A. nervosa is the out-group to A. osyrensis and A. laotica (syn. A. splendens), i.e. A. laotica and A. osyrensis have split-off more recently than A. nervosa. We speculate that the most recently split-off taxon, A. laotica, produces ND seeds (Table 3), while the out-group A. nervosa produces PY seeds. Moreover, based on the phylogenetic analysis by Manos et al. (2001), A. nervosa is the out-group to A. osyrensis, A. laotica, A. mollis and A. capitiformis (Fig. 7). According to Ke et al. (1995), fruits of A. laotica, A. mollis and A. capitiformis are berries, and thus we speculate that the seeds do not have PY. We suggest that ND is a derived character in seeds of these Argyreia species and that it is a reversal of PY to the most ancestral dormancy state of the family Convolvulaceae, i.e. ND in Erycibe and Humbertia as described by Jayasuriya et al. (2009).
Proposed pathway for the evolution of seed dormancy in clade Convolvuloideae of the Convolvulaceae.
Jayasuriya et al. (2009) suggested that the PY of species in Convolvulaceae evolved when the Convolvulaceae species dispersed to seasonal habitats like those in dry-tropical and temperate biomes. Further, these authors have shown that in several taxa PY of seeds has reverted back to ND as these species became redistributed in the aseasonal habitats like those in tropical rain forests. Tribe Maripeae in the clade Dicranostyloideae and Bonamia menziesii are examples of taxa in which PY has reverted back to ND, which is the most ancestral state of seed dormancy in the family. Our study showed that the same phenomenon has occurred in the most recently split-off genus (Argyreia) in the most recently split-off tribe (Ipomoeeae) in Convolvulaceae, where some Argyreia species that are distributed in aseasonal habitats (mainly the tropical wet habitats) have evolved ND seeds.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Athugala YS , Jayasuriya KMGG, Gunaratne AMTA, Baskin CC. 2021. Seed dormancy of 80 tropical montane forest species in Sri Lanka, the first dormancy profile for a tropical montane forest community. Plant Biology Journal 23: 293–299.10.1111/plb.1320333091220 · doi ↗ · pubmed ↗
- 2Austin DF. 1973. The American Erycibeae (Convolvulaceae): Maripa, Dicranostyles, and Lysiostyles I. Systematics. Annals of the Missouri Botanical Garden 60:306–412.
- 3Austin DF. 1980. Convolvulaceae. In: Dassanayake MD, Fosberg FR, eds. A Revised Handbook to the Flora of Ceylon, Volume I. Boca Raton: CRC Press, 288–263.
- 4Austin DF. 1998. Parallel and convergent evolution in the Convolvulaceae. In: Mathews P, Sivadasan M, eds. Biodiversity and taxonomy of tropical flowering plants. Calicut: Mentor Books, 201–234.
- 5Baskin CC , Baskin JM. 2014. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. Second edition. San Diego: Academic Press.
- 6Baskin JM , Baskin CC. 2022. Seed (true seed plus endocarp) dormancy in Anacardiaceae in relation to infrafamilial taxonomy and endocarp anatomy. Seed Science Research 32:187–199.
- 7Baskin JM , Baskin CC, Li X. 2000. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biology 15:139–152.
- 8Dalling JW , Davis AS, Schutte BJ, Arnold AE. 2011. Seed survival in soil: interacting effects of predation, dormancy and the soil microbial community. Journal of Ecology 99:89–95.